
Comparison of Behavior and Space Use of the European Bullhead Cottus gobio and the Round Goby Neogobius melanostomus in a Simulated Natural Habitat

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Acquisition and Maintenance
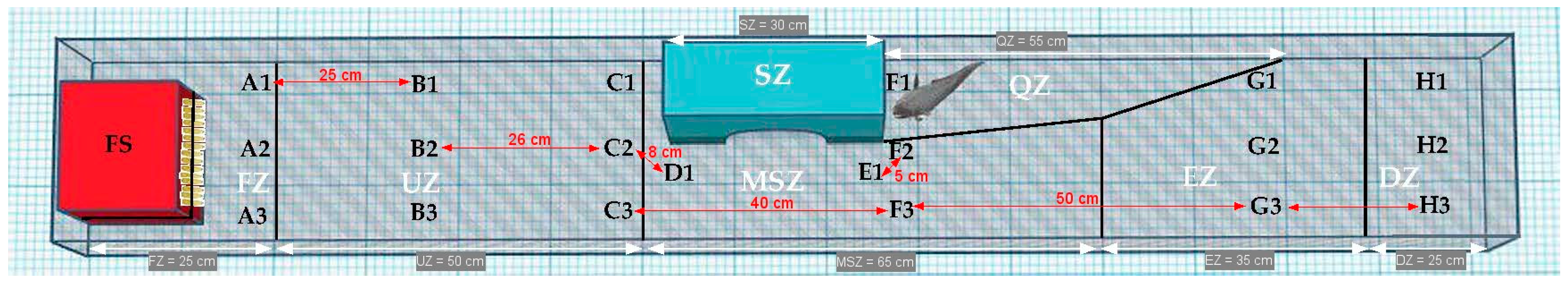
2.2. Experimental Setup
2.3. Statistical Analyses
3. Results
3.1. Water Velocity
3.2. Food Intake
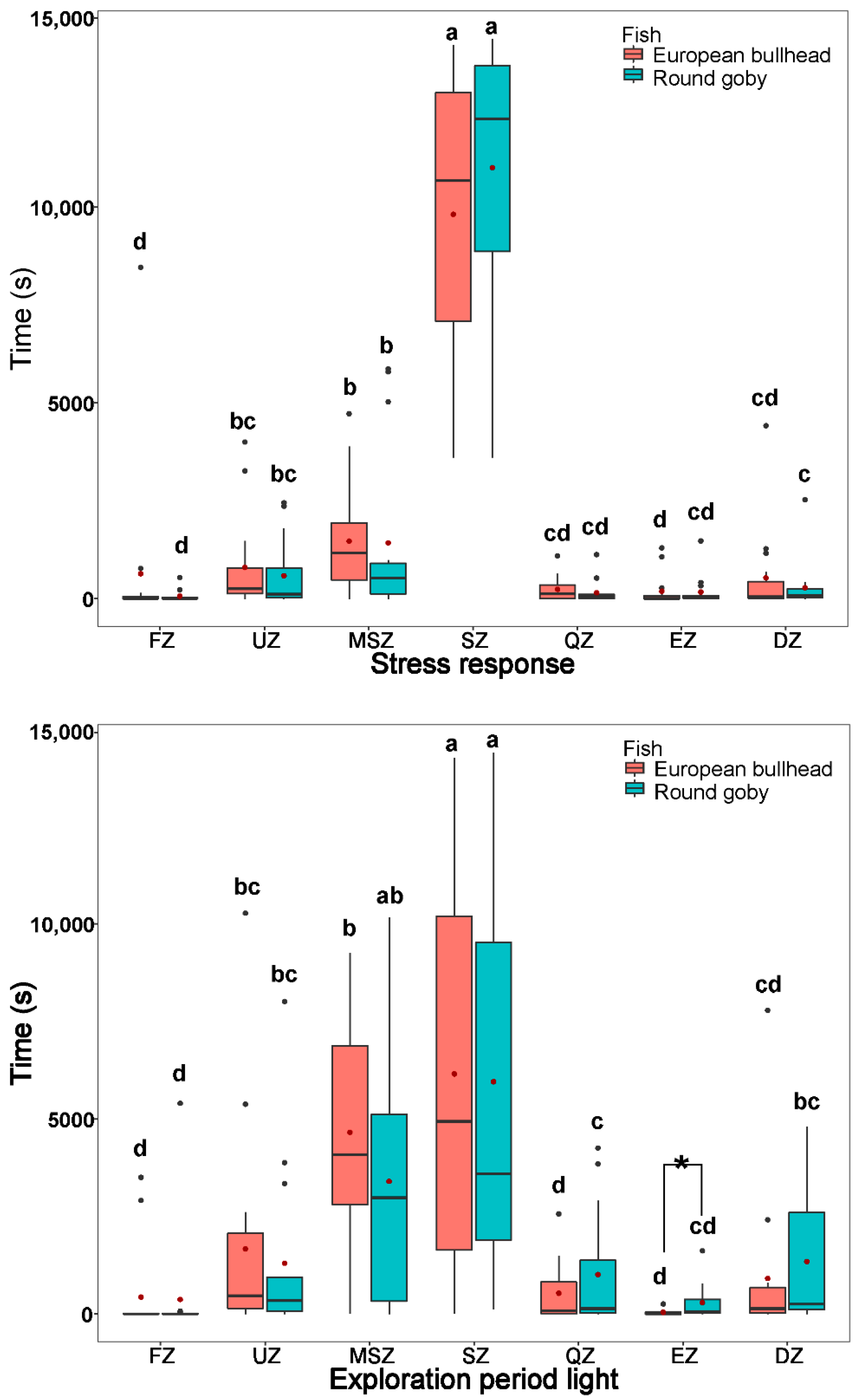
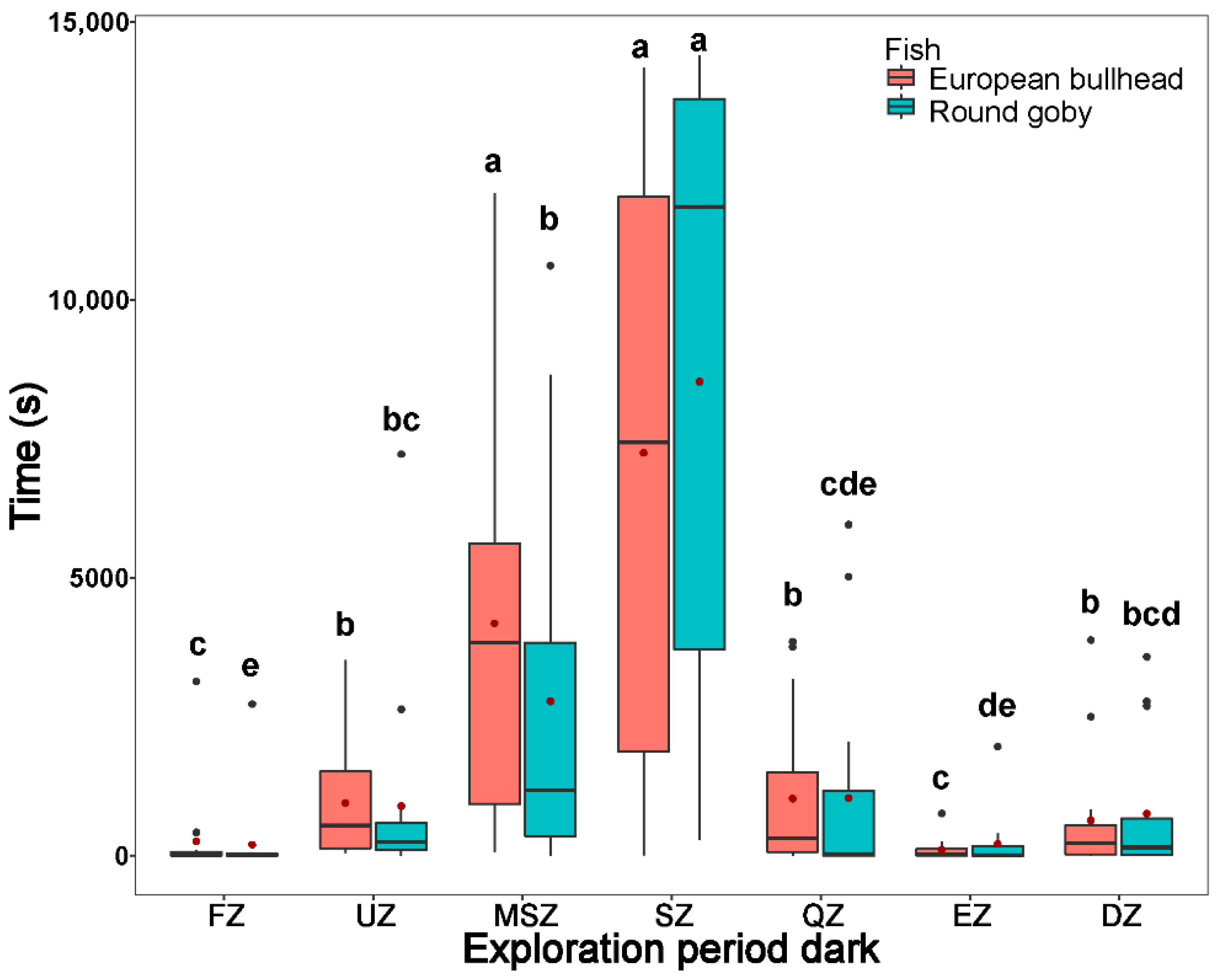
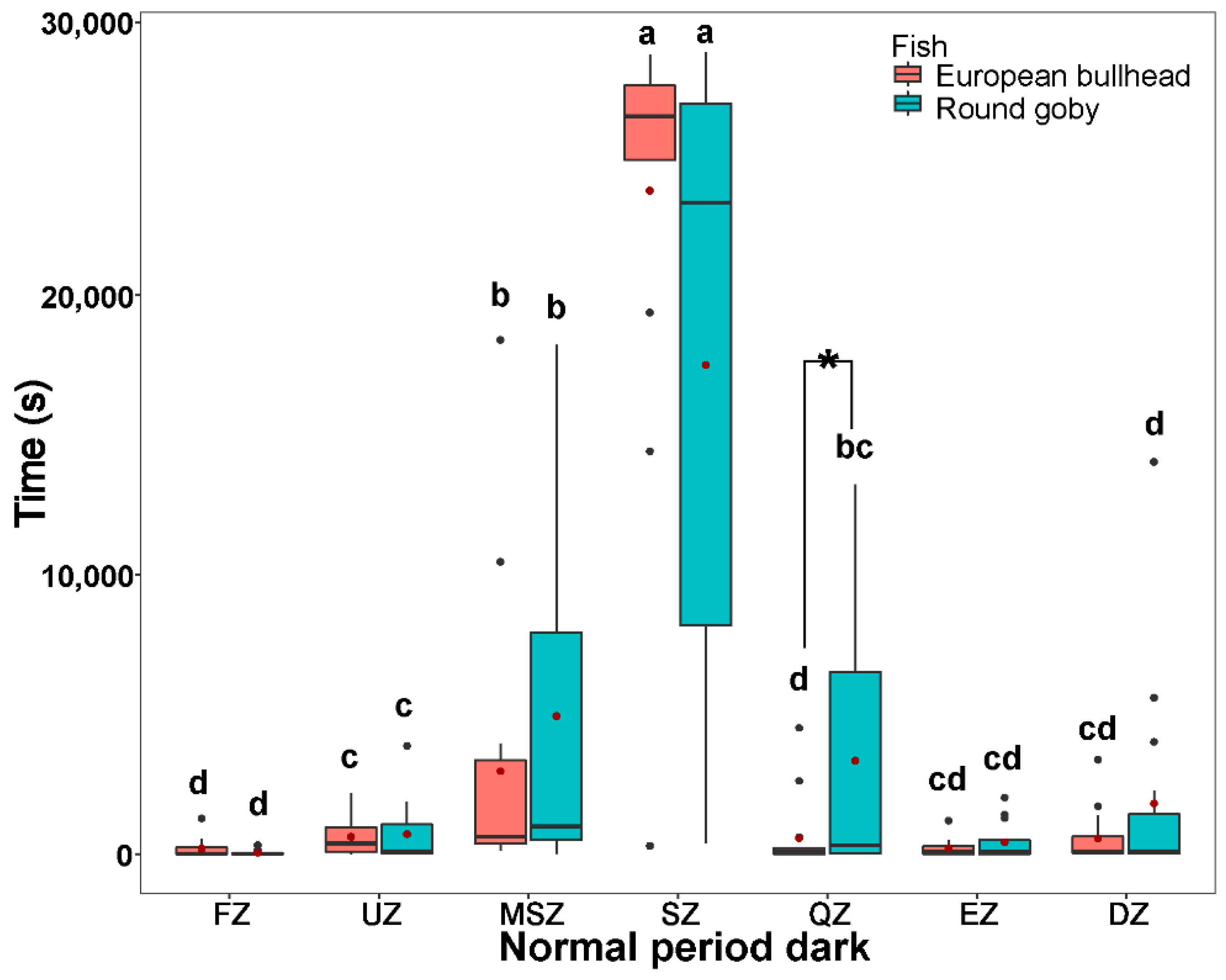
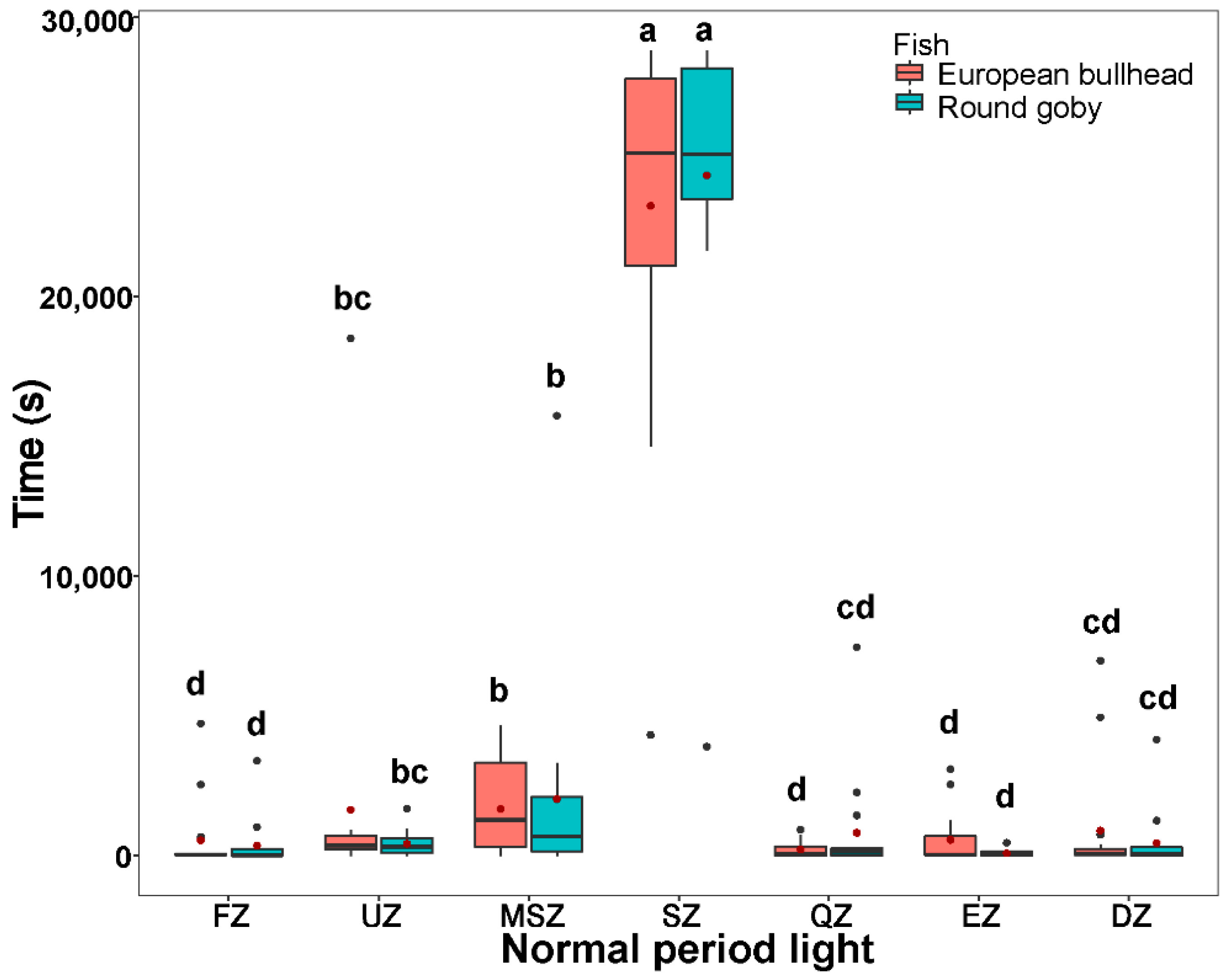
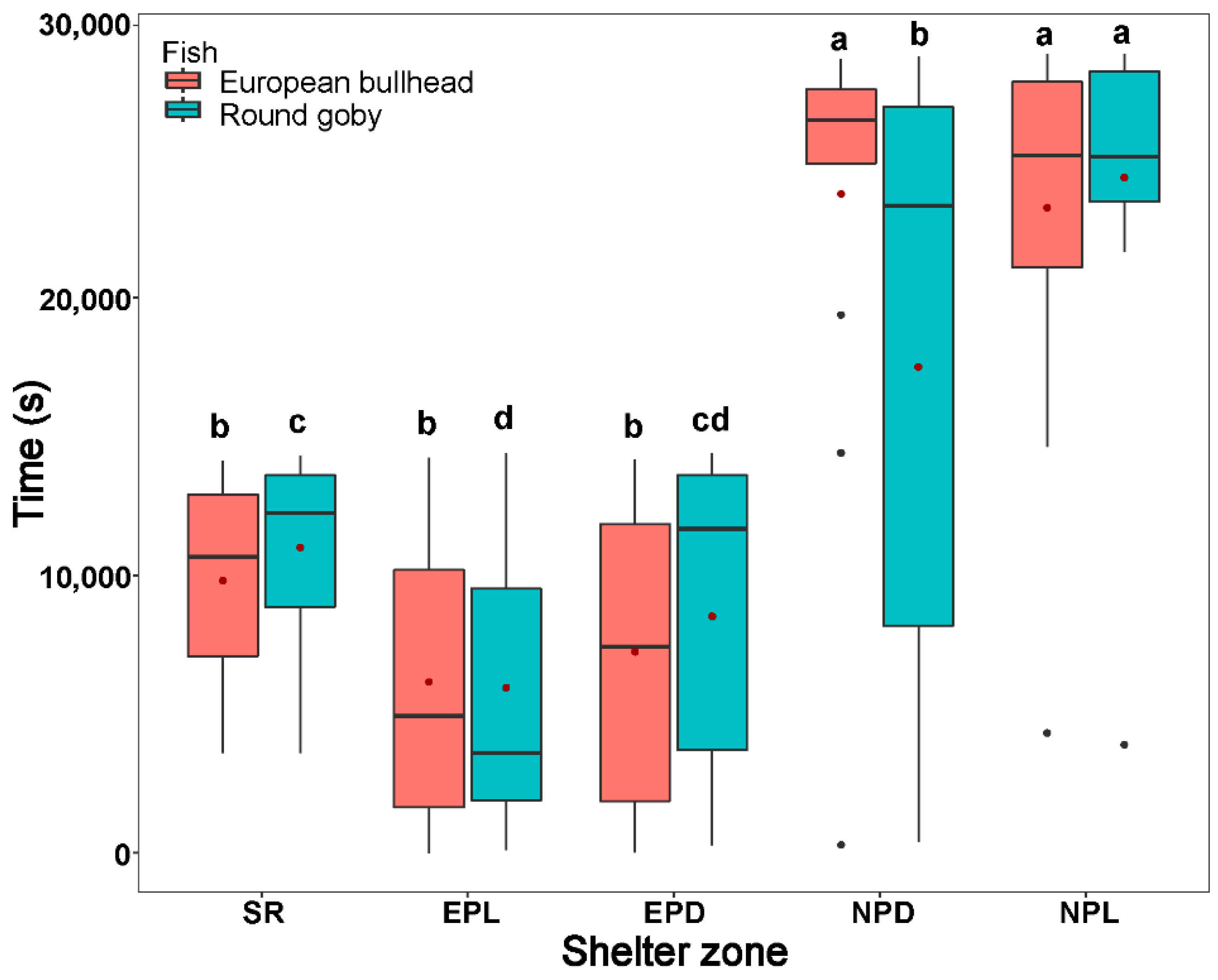
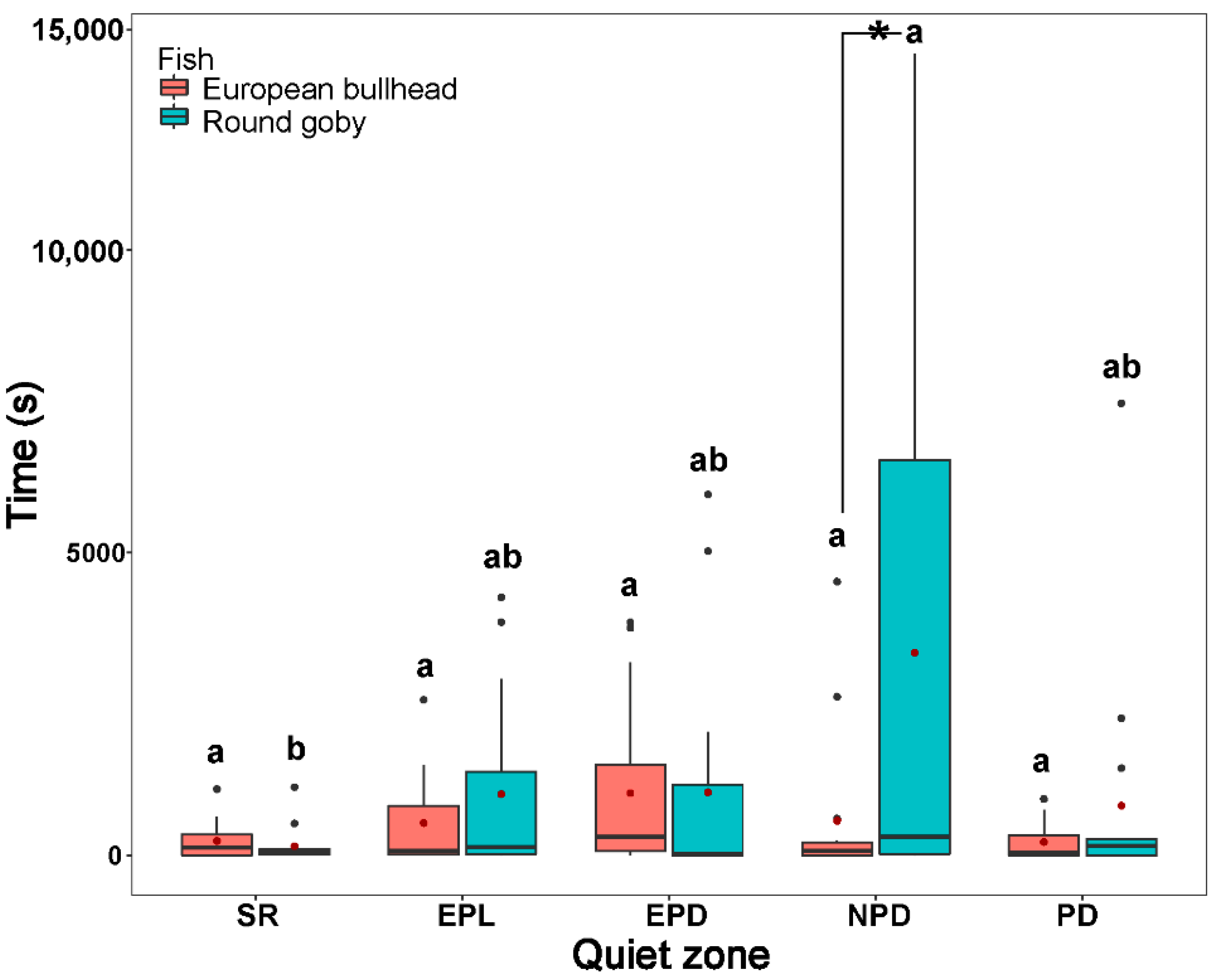
3.3. Spatial Pattern in Fish Diurnal/Nocturnal Activity
3.4. The Temporal Pattern in Fish Diurnal/Nocturnal Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ricciardi, A.; Palmer, M.E.; Yan, N.D. Should biological invasions be managed as natural disasters? Bioscience 2011, 61, 312–317. [Google Scholar] [CrossRef]
- Kakareko, T.; Kobak, J.; Poznańska, M.; Jermacz, Ł.; Copp, G.H. Underwater evaluation of habitat partitioning in a European river between a non-native invader, the racer goby and a threatened native fish, the European bullhead. Ecol. Freshw. Fish 2016, 25, 60–71. [Google Scholar] [CrossRef]
- Gherardi, F. Biological invasions in inland waters: An overview. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Gherardi, F., Ed.; Invading Nature: Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2007; pp. 3–25. [Google Scholar] [CrossRef]
- Janssen, J.; Jude, D.J. Recruitment failure of mottled sculpin Cottus bairdi in Calumet Harbor, southern Lake Michigan, induced by the newly introduced round goby Neogobius melanostomus. J. Great Lakes Res. 2001, 27, 319–328. [Google Scholar] [CrossRef]
- Waage, J.; Krebs, J.R.; Davies, N.B. Behavioural Ecology. J. Anim. Ecol. 1981, 50, 332. [Google Scholar] [CrossRef]
- Gebauer, R.; Veselý, L.; Vanina, T.; Buřič, M.; Kouba, A.; Drozd, B. Prediction of ecological impact of two alien gobiids in habitat structures of differing complexity. Can. J. Fish. Aquat. Sci. 2019, 76, 1954–1961. [Google Scholar] [CrossRef]
- Genin, A.; Karp, L.; Miroz, A. Effects of flow on competitive superiority in scleractinian corals. Limnol. Oceanogr. 1994, 39, 913–924. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Nakano, S. Condition-specific competition: Implications for the altitudinal distribution of stream fishes. Ecology 2000, 81, 2027. [Google Scholar] [CrossRef]
- Jermacz, L.; Kobak, J.; Dzierzyńska, A.; Kakareko, T. The effect of flow on the competition between the alien racer goby and native European bullhead. Ecol. Freshw. Fish 2015, 24, 467–477. [Google Scholar] [CrossRef]
- Bussmann, K.; Burkhardt-Holm, P. Round gobies in the third dimension—Use of vertical walls as habitat enables vector contact in a bottom-dwelling invasive fish. Aquat. Invasions 2020, 15, 683–699. [Google Scholar] [CrossRef]
- Šlapanský, L.; Janáč, M.; Roche, K.; Jurajda, P. Round goby movement patterns in a non-navigable river. Can. J. Fish. Aquat. Sci. 2020, 77, 475–483. [Google Scholar] [CrossRef]
- Buřič, M.; Worischka, S.; Richter, L.; Drozd, B.; Bláha, M.; Berendonk, T. Aquatische Invasive Neozoen in Sachsen und dem Norden der Tschechischen Republik—Mögliche Nachhaltige Management-Strategien für die Projektregion/Invazní Sladkovodní Druhy Saska a Severních Čech—Možné Udržitelné Strategie Managementu v Projektovém Region; European Union: Brussels, Belgium, 2020; Volume 63. [Google Scholar] [CrossRef]
- Brandner, J.; Cerwenka, A.F.; Schliewen, U.K.; Geist, J. Bigger is better: Characteristics of round gobies forming an invasion front in the Danube River. PLoS ONE 2013, 8, e73036. [Google Scholar] [CrossRef] [PubMed]
- Ojaveer, H. The round goby Neogobius melanostomus is colonising the NE Baltic Sea. Aquat. Invasions 2006, 1, 44–45. [Google Scholar] [CrossRef]
- Ähnelt, H.; Duchkowitsch, M.; Scattolin, G.; Zweimüller, I.; Weissenbacher, A. Neogobius gymnotrachelus (Kessler, 1857) (Teleostei: Gobiidae), die Nackthals-Grundel in Österreich. Österreichs Fischerei 2001, 54, 262–266. [Google Scholar]
- Ähnelt, H.; Bänärescu, P.; Spolwind, R.; Harka, Á.; Waidbacher, H. Occurrence and distribution of three gobiid species (Pisces, Gobiidae) in the middle and upper Danube region—Examples of different dispersal patterns? Biologia 1998, 53, 665–678. [Google Scholar]
- Freyhof, J. Immigration and potential impacts of invasive freshwater fishes in Germany. Regul. Biocoenoces 2003, 17, 51–58. [Google Scholar]
- Kessel, N.V.; Dorenbosch, M.; Spikmans, F. First record of Pontian monkey goby, Neogobius fluviatilis (Pallas, 1814), in the Dutch Rhine. Aquat. Invasions 2009, 4, 421–424. [Google Scholar] [CrossRef]
- Grabowska, J.; Pietraszewski, D.; Ondračková, M. Tubenose goby Proterorhinus marmoratus (Pallas, 1814) has joined three other Ponto-Caspian gobies in the Vistula River (Poland). Aquat. Invasions 2008, 3, 261–265. [Google Scholar] [CrossRef]
- Copp, G.H.; Bianco, P.G.; Bogutskaya, N.G.; Erős, T.; Falka, I.; Ferreira, M.T.; Fox, M.G.; Freyhof, J.; Gozlan, R.E.; Grabowska, J.; et al. To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 2005, 21, 242–262. [Google Scholar] [CrossRef]
- Lusk, S.; Vetešník, L.; Halačka, K.; Lusková, V.; Pekárik, L.; Tomeček, J. Round goby Neogobius (Apollonia) melanostomus recorded for the first time in the confluence area of the Morava and Dyje rivers, Czech Republic. Biodivers. Fishes Czech Repub. 2008, 7, 114–118. [Google Scholar]
- Buřič, M.; Bláha, M.; Kouba, A.; Drozd, B. Upstream expansion of round goby (Neogobius Melanostomus)—First record in the upper reaches of the Elbe River. Knowl. Manag. Aquat. Ecosyst. 2015, 32, 1–5. [Google Scholar] [CrossRef]
- Roche, K.; Janáč, M.; Šlapanský, L.; Mikl, L.; Kopeček, L.; Jurajda, P. A newly established round goby (Neogobius melanostomus) population in the upper stretch of the river Elbe. Knowl. Manag. Aquat. Ecosyst. 2015, 33, 1–11. [Google Scholar] [CrossRef]
- Sapota, M.R. The round goby (Neogobius melanostomus) in the Gulf of Gdańsk—A species introduction into the Baltic Sea. Hydrobiologia 2004, 514, 219–224. [Google Scholar] [CrossRef]
- Charlebois, P.M.; Marsden, J.E.; Goettel, R.G.; Wolfe, R.K.; Jude, D.J.; Rudnika, S. The round goby, Neogobius melanostomus (Pallas): A review of European and North American literature. Illinois-Indiana Sea Grant Program and Illinois Natural History Survey. Ill. Nat. Hist. Surv., Spec. Publ. 1997, 20, 1–76. [Google Scholar]
- Smirnov, A.I. Perciformes (Gobioidei), Scorpaeniformes, Pleuronectiformes, Gobiesociformes, Lophiiformes; Naukova Dumka: Kiev, Ukraine, 1986; Volume 8. [Google Scholar]
- Verliin, A.; Kesler, M.; Svirgsden, R.; Taal, I.; Saks, L.; Rohtla, M.; Hubel, K.; Eschbaum, R.; Vetemaa, M.; Saat, T. Invasion of round goby to the temperate salmonid streams in the Baltic Sea. Ichthyol. Res. 2017, 64, 155–158. [Google Scholar] [CrossRef]
- Grabowska, J.; Kakareko, T.; Błońska, D.; Przybylski, M.; Kobak, J.; Jermacz, Ł.; Copp, G.H. Interspecific competition for a shelter between non-native racer goby and native European bullhead under experimental conditions—Effects of season, fish size and light conditions. Limnologica 2016, 56, 30–38. [Google Scholar] [CrossRef]
- Kessel, N.V.; Dorenbosch, M.; De Boer, M.R.M.; Leuven, R.S.E.W.; Van der Velde, G. Competition for shelter between four invasive gobiids and two native benthic fish species. Curr. Zool. 2011, 57, 844–851. [Google Scholar] [CrossRef]
- Knaepkens, G.; Verheyen, E.; Galbusera, P.; Eens, M. The use of genetic tools for the evaluation of a potential migration barrier for the bullhead. J. Fish Biol. 2004, 64, 1737–1744. [Google Scholar] [CrossRef]
- Lelek, A.; Wheeler, A. The Freshwater Fishes of Europe. Vol. 9. Threatened Fishes of Europe. European Committee for the Conservation of Nature and natural resources. Aula-Verlag, Wiesbaden, Germany. Copeia 1988, 9, 1099–1100. [Google Scholar] [CrossRef]
- Janáč, M.; Roche, K.; Šlapanský, L.; Polačik, M.; Jurajda, P. Long-term monitoring of native bullhead and invasive gobiids in the Danubian rip-rap zone. Hydrobiologia 2018, 807, 263–275. [Google Scholar] [CrossRef]
- Knaepkens, G.; Bruyndoncx, L.; Coeck, J.; Eens, M. Spawning habitat enhancement in the European bullhead (Cottus gobio), an endangered freshwater fish in degraded lowland rivers. Biodivers. Conserv. 2004, 13, 2443–2452. [Google Scholar] [CrossRef]
- Andreasson, S. Locomotory activity patterns of Cottus poecilopus Heckel and C. gobio L. (Pisces). Oikos 1969, 20, 78. [Google Scholar] [CrossRef]
- Kakareko, T.; Kobak, J.; Grabowska, J.; Jermacz, Ł.; Przybylski, M.; Poznańska, M.; Pietraszewski, D.; Copp, G.H. Competitive interactions for food resources between invasive racer goby Babka gymnotrachelus and native European bullhead Cottus gobio. Biol. Invasions 2013, 15, 2519–2530. [Google Scholar] [CrossRef][Green Version]
- Dubs, D.O.L.; Corkum, L.D. Behavioral interactions between round gobies (Neogobius melanostomus) and mottled sculpins (Cottus bairdi). J. Great Lakes Res. 1996, 22, 838–844. [Google Scholar] [CrossRef]
- Savino, J.F.; Riley, S.C.; Holuszko, M.J. Activity, aggression, and habitat use of ruffe (Gymnocephalus cernuus) and round goby (Apollonia melanostoma) under laboratory conditions. J. Great Lakes Res. 2007, 33, 326–334. [Google Scholar] [CrossRef]
- Knaepkens, G.; Bervoets, L.; Verheyen, E.; Eens, M. Relationship between population size and genetic diversity in endangered populations of the European bullhead (Cottus gobio): Implications for conservation. Biol. Conserv. 2004, 115, 403–410. [Google Scholar] [CrossRef]
- Jurajda, P.; Černý, J.; Polačik, M.; Valová, Z.; Janáč, M.; Blažek, R.; Ondračková, M. The recent distribution and abundance of non-native Neogobius fishes in the Slovak section of the River Danube. J. Appl. Ichthyol. 2005, 21, 319–323. [Google Scholar] [CrossRef]
- Dorenbosch, M.; Van der Velde, G. Population increment of native and alien fish species in the Dutch rivers Rhine and Meuse: Competition and relations with environmental variables. In Proceedings of the XVI International Conference on Aquatic Invasive Species (ICAIS), Montreal, QC, Canada, 19–23 April 2009; pp. 19–23. [Google Scholar]
- Corkum, L.D.; Sapota, M.R.; Skora, K.E. The round goby, Neogobius melanostomus, a fish invader on both sides of the Atlantic Ocean. Biol. Invasions 2004, 6, 173–181. [Google Scholar] [CrossRef]
- Perello, M.M.; Simon, T.P.; Thompson, H.A.; Kane, D.D. Feeding ecology of the invasive round goby, Neogobius melanostomus (Pallas, 1814), based on laboratory size preference and field diet in different habitats in the western basin of Lake Erie. Aquat. Invasions 2015, 10, 436–474. [Google Scholar] [CrossRef]
- Balshine, S.; Verma, A.; Chant, V.; Theysmeyer, T. Competitive interactions between round gobies and logperch. J. Great Lakes Res. 2005, 31, 68–77. [Google Scholar] [CrossRef]
- Bergstrom, M.A.; Mensinger, A.F. Interspecific resource competition between the invasive round goby and three native species: Logperch, slimy sculpin, and spoonhead sculpin. Trans. Am. Fish. Soc. 2009, 138, 1009–1017. [Google Scholar] [CrossRef]
- Freyhof, J.; Kottelat, M.; Nolte, A. Taxonomic diversity of European Cottus with description of eight new species (Teleostei: Cottidae). Ichthyol. Explor. Freshw. 2005, 16, 107. [Google Scholar]
- Kessel, N.V.; Dorenbosch, M.; Kranenbarg, J.; Van der Velde, G.; Leuven, R.S.E.W. Invasive Ponto-Caspian gobies rapidly reduce the abundance of protected native bullhead. Aquat. Invasions 2016, 11, 179–188. [Google Scholar] [CrossRef]
- Tomlinson, M.L.; Perrow, M.R. Ecology of the bullhead. Conserving Natura 2000 Rivers. English Nature, Peterborough, England. Ecol. Ser. 2003, 4, 16–19. [Google Scholar]
- Miller, P.J.; Bath, H. The freshwater fishes of Europe, Vol. 8. I Mugilidae, Atherinidae, Atherinopsidae, Blenniidae, Odontobutidae, Gobidae 1. In The Freshwater Fishes of Europe, Vol. 8. I Mugilidae, Atherinidae, Atherinopsidae, Blenniidae, Odontobutidae, Gobidae 1; Miller, P.J., Bath, H., Eds.; AULA-Verlag: Wiebelsheim, Germany, 2003; pp. 264–279. [Google Scholar]
- Smyly, W.J.P. The life-history of the bullhead or Miller’s thumb (Cottus gobio L.). Proc. Zool. Soc. Lond. 1957, 128, 431–454. [Google Scholar] [CrossRef]
- Jude, D.J. Round gobies: Cyberfish of the third millennium. Great Lakes Res. Rev. 1997, 3, 27–34. [Google Scholar]
- Andreasson, S. Feeding habits of a sculpin (Cottus gobio L. Pisces) population. Rep. Inst. Freshw. Res. Drottingholm 1971, 51, 5–30. [Google Scholar]
- Fischer, S.; Kummer, H. Effects of residual flow and habitat fragmentation on distribution and movement of bullhead (Cottus gobio L.) in an alpine stream. In Assessing the Ecological Integrity of Running Waters; Springer: Dordrecht, The Netherlands, 2000; pp. 305–317. [Google Scholar] [CrossRef]
- Tudorache, C.; Viaene, P.; Blust, R.; Vereecken, H.; De Boeck, G. A comparison of swimming capacity and energy use in seven European freshwater fish species. Ecol. Freshw. Fish 2008, 17, 284–291. [Google Scholar] [CrossRef]
- Kornis, M.S.; Vander Zanden, M.J. Forecasting the distribution of the invasive round goby (Neogobius melanostomus) in Wisconsin tributaries to Lake Michigan. Can. J. Fish. Aquat. Sci. 2010, 67, 553–562. [Google Scholar] [CrossRef]
- Egger, B.; Wiegleb, J.; Seidel, F.; Burkhardt-Holm, P.; Emanuel Hirsch, P. Comparative swimming performance and behaviour of three benthic fish species: The invasive round goby (Neogobius melanostomus), the native bullhead (Cottus gobio), and the native gudgeon (Gobio gobio). Ecol. Freshw. Fish 2020, 30, 391–405. [Google Scholar] [CrossRef]
- Laverty, C.; Green, K.D.; Dick, J.T.; Barrios-O’Neill, D.; Mensink, P.J.; Médoc, V.; Spataro, T.; Caffrey, J.M.; Lucy, F.E.; Boets, P.; et al. Assessing the ecological impacts of invasive species based on their functional responses and abundances. Biol. Invasions 2017, 19, 1653–1665. [Google Scholar] [CrossRef]
- Carter, M.G.; Copp, G.H.; Szomlai, V. Seasonal abundance and microhabitat use of bullhead Cottus gobio and accompanying fish species in the River Avon (Hampshire), and implications for conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 395–412. [Google Scholar] [CrossRef]
- Roje, S.; Richter, L.; Worischka, S.; Let, M.; Veselý, L.; Buřič, M. Round goby versus marbled crayfish: Alien invasive predators and competitors. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 1–10. [Google Scholar] [CrossRef]
- Franta, P.; Gebauer, R.; Veselý, L.; Buřič, M.; Szydłowska, N.Z.; Drozd, B. The Invasive Round Goby Neogobius melanostomus as a Potential Threat to Native Crayfish Populations. Animals 2021, 11, 2377. [Google Scholar] [CrossRef]
- Gosselin, M.P.; Petts, G.E.; Maddock, I.P. Mesohabitat use by bullhead (Cottus gobio). Hydrobiologia 2010, 652, 299–310. [Google Scholar] [CrossRef]
- Knaepkens, G.; Bruyndoncx, L.; Bervoets, L.; Eens, M. The presence of artificial stones predicts the occurrence of the European bullhead (Cottus gobio) in a regulated lowland river in Flanders (Belgium). Ecol. Freshw. Fish 2002, 11, 203–206. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | n | TL (mm) | SL (mm) | W (g) | Eaten Larvae |
|---|---|---|---|---|---|
| RG | 15 (12F-3M) | 85.3 ± 11.5 | 70.7 ± 9.7 | 8.5 ± 5.9 | 4.7 ± 5.0 a |
| EB | 15 | 93.3 ± 13.3 | 78.7 ± 12.5 | 8.3 ± 3.9 | 4.7 ± 5.1 a |
| Species | Time Period | Distance Moved (cm) | Outside the Shelter (s) | Outside the Shelter (%) | Motion outside the Shelter (sec) | Motion outside the Shelter (%) | Active Movement during the Trial (%) |
|---|---|---|---|---|---|---|---|
| RG | SR | 1129.5 ± 1456.1 a | 2671.5 ± 2693.0 a | 18.6 ± 18.7 c | 1058.0 ± 881.6 a | 53.6 ± 27.4 b | 7.3 ± 6.1 c |
| EB | SR | 1551.4 ± 1624.4 a | 3864.8 ± 3077.1 a | 26.8 ± 21.4 b,c | 1091.9 ± 975.5 a | 35.5 ± 23.4 b | 7.6 ± 6.8 c |
| RG | EPL | 3194.2 ± 2871.4 a | 7720.0 ± 5265.8 a | 53.6 ± 36.6 b | 2556.5 ± 2603.6 a | 35.5 ± 28.5 b | 17.8 ± 18.1 b |
| EB | EPL | 3773.8 ± 3342.2 b | 8242.5 ± 5117.0 a | 57.2 ± 35.5 c | 2415.2 ± 1716.5 b | 39.8 ± 28.1 c | 16.8 ± 11.9 d |
| RG | EPD | 2478.8 ± 2410.7 a | 3217.1 ± 4501.2 a | 22.3 ± 31.3 c,d | 1119.2 ± 1600.3 a,b | 46.7 ± 27.5 b,c | 7.8 ± 11.1 d |
| EB | EPD | 2951.4 ± 2579.4 a | 2030.4 ± 2217.1 b | 14.1 ± 15.4 d | 659.7 ± 579.3 b | 47.2 ± 32.1 c | 4.6 ± 4.0 d |
| RG | NPD | 2294.0 ± 3761.2 b | 10622.4 ± 8817.1 a | 36.8 ± 30.7 c | 4234.0 ± 5488.9 b | 44.5 ± 27.1 c | 14.6 ± 18.9 d |
| EB | NPD | 1207.1 ± 1234.3 c | 10411.7 ± 10275.1 a | 36.1 ± 35.6 d | 3640.2 ± 4753.0 b | 45.4 ± 25.1 d | 12.6 ± 16.5 e |
| RG | NPL | 1772.6 ± 2797.5 b | 9796.1 ± 8455.6 a | 34.1 ± 29.4 c,d | 4933.0 ± 4637.1 a | 52.5 ± 26.1 c | 17.1 ± 16.2 d |
| EB | NPL | 1828.3 ± 2117.4 b | 6828.0 ± 7682.5 a | 23.7 ± 26.7 d | 2864.3 ± 3243.2 a,b | 54.8 ± 22.9 c | 9.9 ± 11.3 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roje, S.; Drozd, B.; Richter, L.; Kubec, J.; Polívka, Z.; Worischka, S.; Buřič, M. Comparison of Behavior and Space Use of the European Bullhead Cottus gobio and the Round Goby Neogobius melanostomus in a Simulated Natural Habitat. Biology 2021, 10, 821. https://doi.org/10.3390/biology10090821
Roje S, Drozd B, Richter L, Kubec J, Polívka Z, Worischka S, Buřič M. Comparison of Behavior and Space Use of the European Bullhead Cottus gobio and the Round Goby Neogobius melanostomus in a Simulated Natural Habitat. Biology. 2021; 10(9):821. https://doi.org/10.3390/biology10090821
Chicago/Turabian StyleRoje, Sara, Bořek Drozd, Luise Richter, Jan Kubec, Zdeněk Polívka, Susanne Worischka, and Miloš Buřič. 2021. "Comparison of Behavior and Space Use of the European Bullhead Cottus gobio and the Round Goby Neogobius melanostomus in a Simulated Natural Habitat" Biology 10, no. 9: 821. https://doi.org/10.3390/biology10090821
APA StyleRoje, S., Drozd, B., Richter, L., Kubec, J., Polívka, Z., Worischka, S., & Buřič, M. (2021). Comparison of Behavior and Space Use of the European Bullhead Cottus gobio and the Round Goby Neogobius melanostomus in a Simulated Natural Habitat. Biology, 10(9), 821. https://doi.org/10.3390/biology10090821