
Downregulation of Ribosomal Contents and Kinase Activities Is Associated with the Inhibitive Effect on the Growth of Group B Streptococcus Induced by Placental Extracellular Vesicles

 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Collection of Placental EVs
2.2. Collection of Bacteria
2.3. Measurement of Bacterial Growth
2.4. Determination of the Interaction of Placental EVs with Bacteria
2.5. Measurement of Antibiotic Susceptibility
2.6. Proteomic Analysis
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
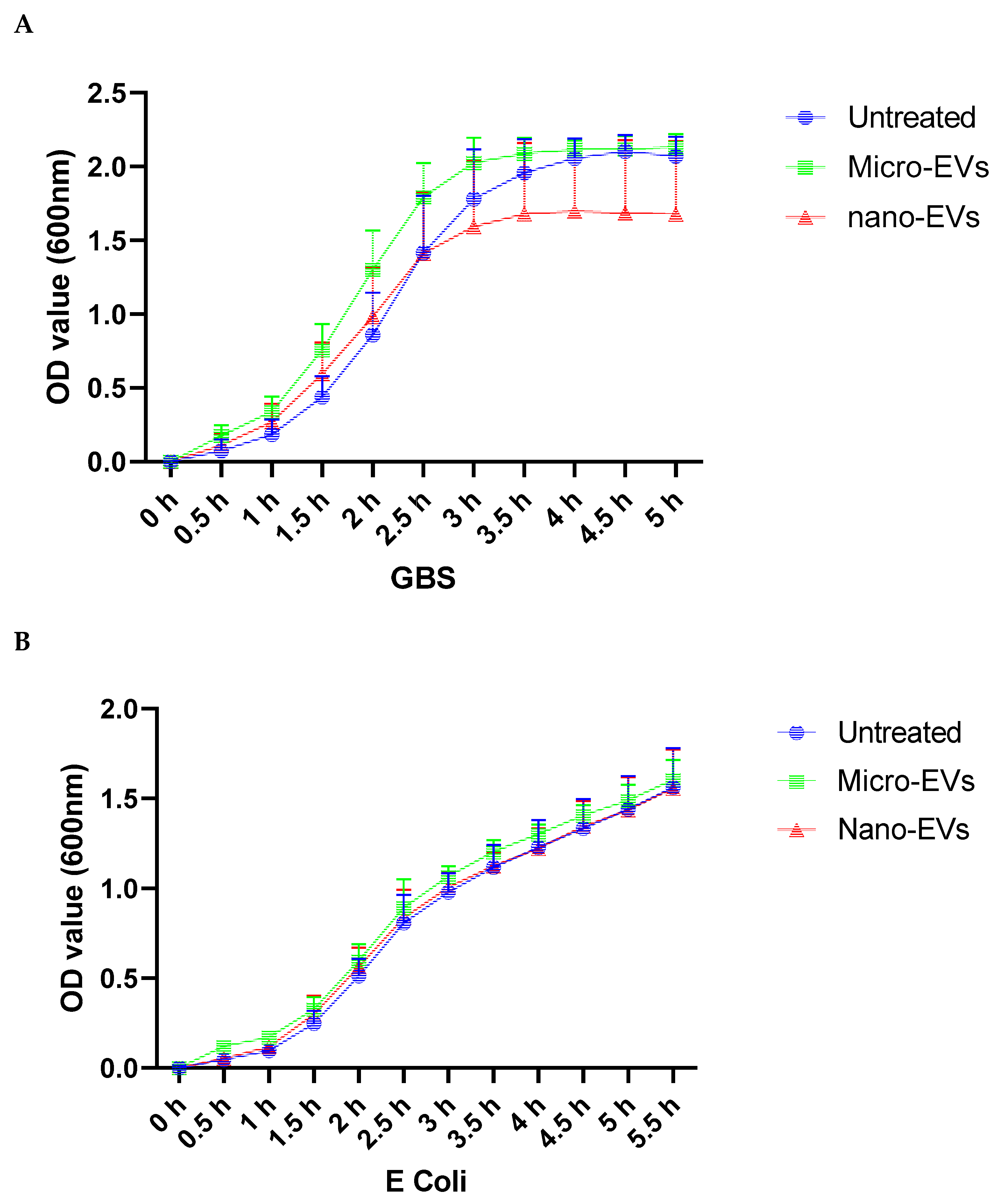
3.1. Placental EVs Attenuated GBS Growth In Vitro
3.2. Placental EVs Inhibited the Growth of GBS
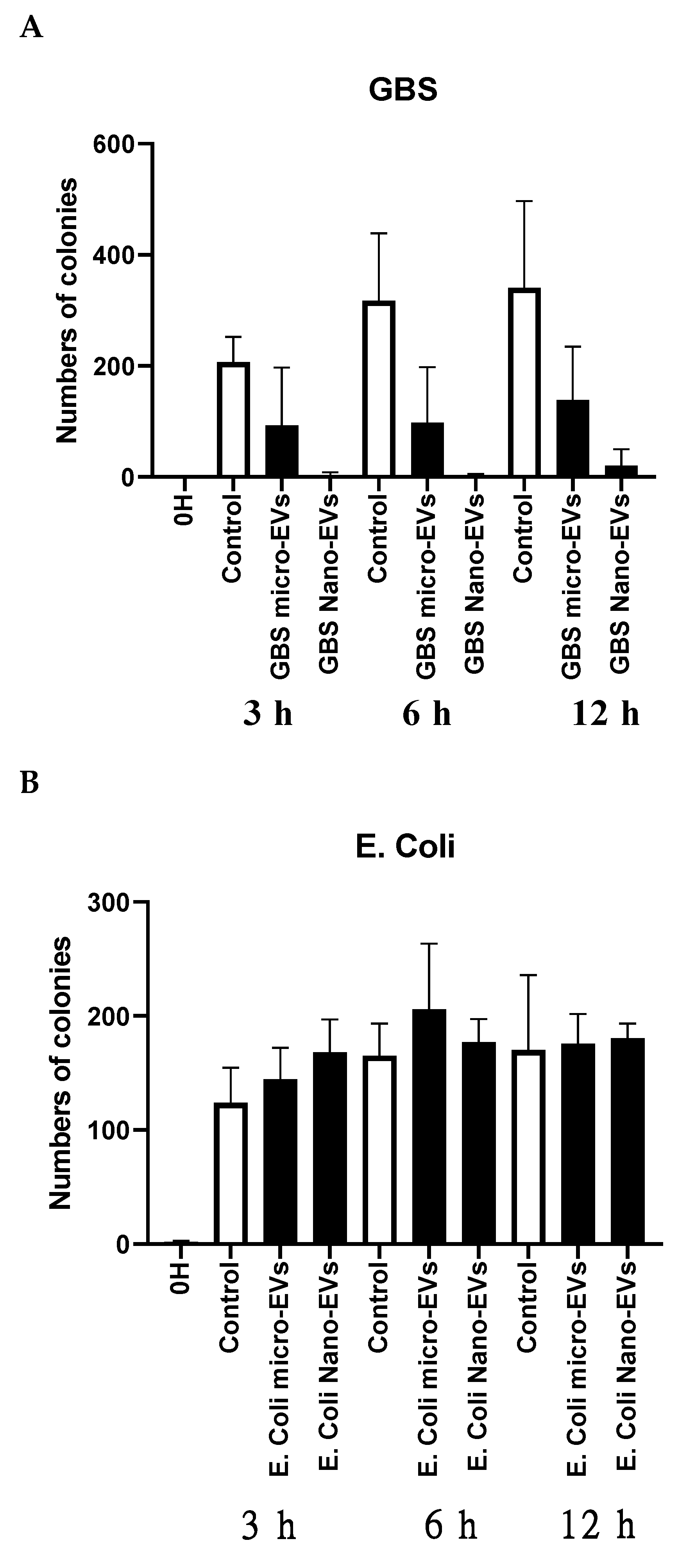
3.3. GBS but Not E. coli Interacted with Placental EVs
3.4. Placental EVs Do Not Affect Bacterial Susceptibility or Resistance to Antibiotics
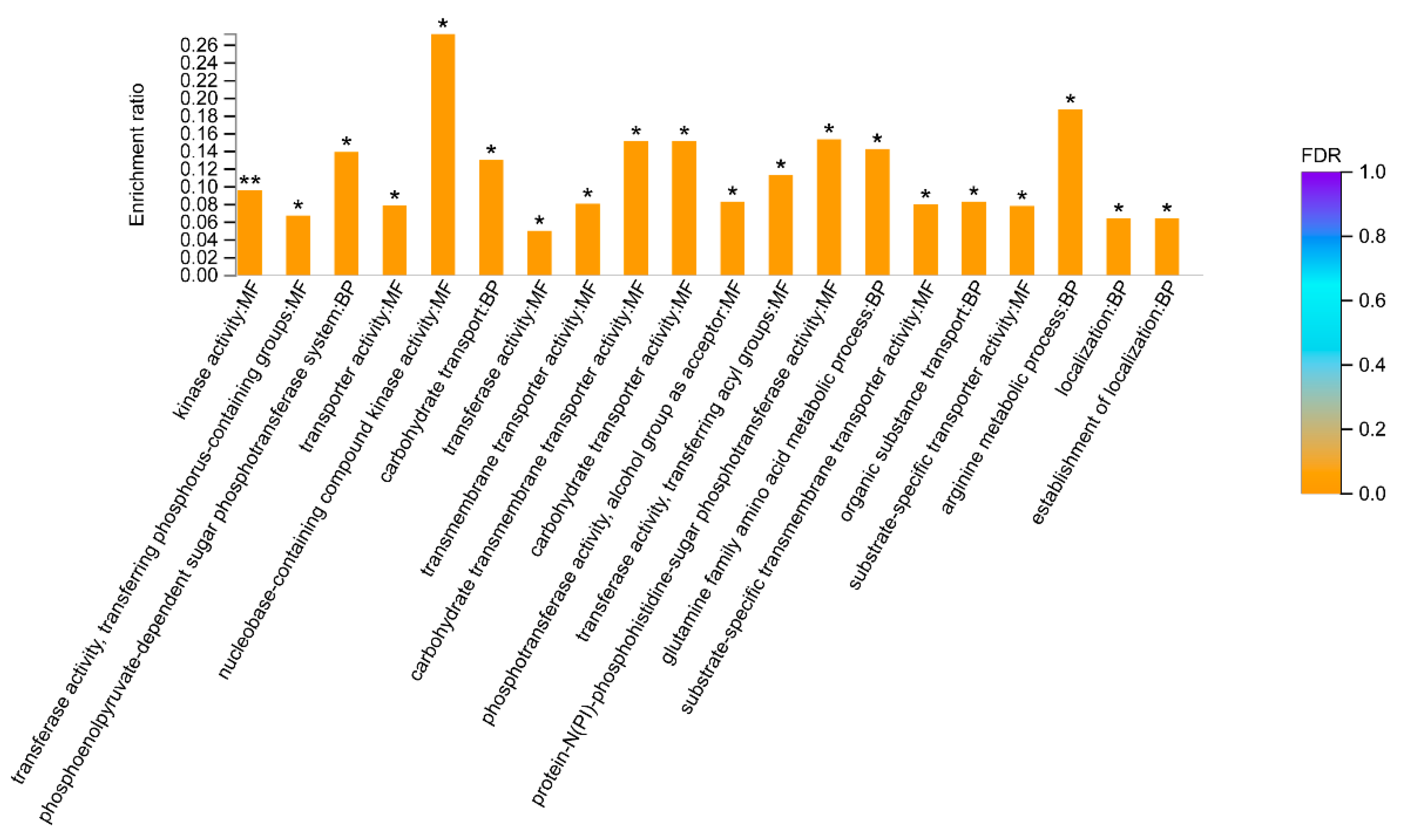
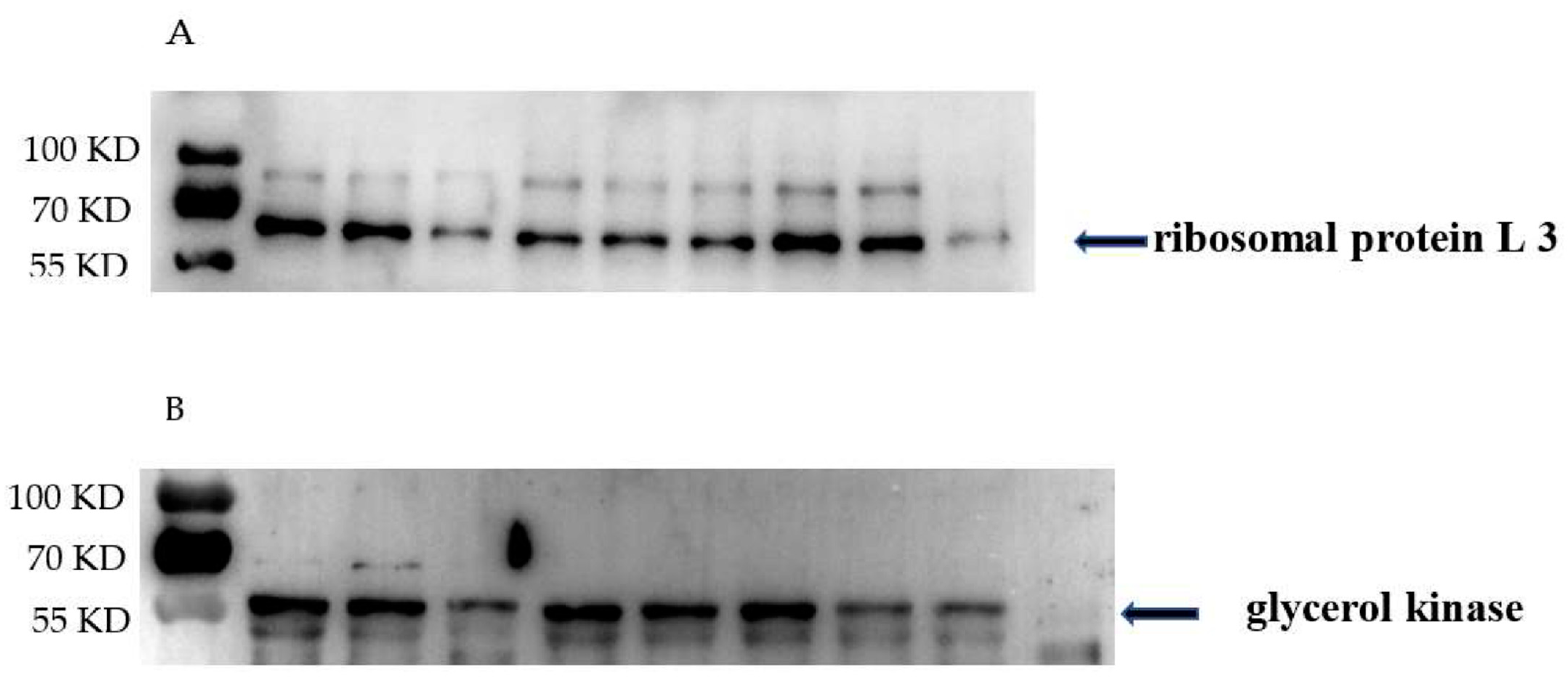
3.5. Proteomic Analysis Revealed Placental EVs Affect the Expression of Growth-Related Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vornhagen, J.; Waldorf, K.M.A.; Rajagopal, L. Perinatal Group B Streptococcal Infections: Virulence Factors, Immunity, and Prevention Strategies. Trends Microbiol. 2017, 25, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Verani, J.R.; Schrag, S.J. Group B Streptococcal Disease in Infants: Progress in Prevention and Continued Challenges. Clin. Perinatol. 2010, 37, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Desa, D.J.; Trevenen, C.L. Intrauterine infections with group B beta-haemolytic streptococci. Br. J. Obstet. Gynaecol. 1984, 91, 237–239. [Google Scholar] [CrossRef] [PubMed]
- McDonald, H.; Vigneswaran, R.; O’Loughlin, J.A. Group B Streptococcal Colonization and Preterm Labour. Aust. N. Z. J. Obstet. Gynaecol. 1989, 29, 291–293. [Google Scholar] [CrossRef]
- Seale, A.C.; Blencowe, H.; Bianchi-Jassir, F.; Embleton, N.; Bassat, Q.; Ordi, J.; Menéndez, C.; Cutland, C.; Briner, C.; Berkley, J.A.; et al. Stillbirth with Group B Streptococcus Disease Worldwide: Systematic Review and Meta-analyses. Clin. Infect. Dis. 2017, 65, S125–S132. [Google Scholar] [CrossRef]
- Sarker, S.; Scholz-Romero, K.; Perez, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E.; Salomon, C. Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy. J. Transl. Med. 2014, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Tong, M.; Chamley, L.W. Placental Extracellular Vesicles and Feto-Maternal Communication. Cold Spring Harb. Perspect. Med. 2015, 5, a023028. [Google Scholar] [CrossRef] [Green Version]
- Buca, D.; Bologna, G.; D’Amico, A.; Cugini, S.; Musca, F.; Febbo, M.; D’Arcangelo, D.; Buca, D.; Simeone, P.; Liberati, M.; et al. Extracellular Vesicles in Feto–Maternal Crosstalk and Pregnancy Disorders. Int. J. Mol. Sci. 2020, 21, 2120. [Google Scholar] [CrossRef] [Green Version]
- Sheller-Miller, S.; Choi, K.; Choi, C.; Menon, R. Cyclic-recombinase-reporter mouse model to determine exosome communication and function during pregnancy. Am. J. Obstet. Gynecol. 2019, 221, 502.e1–502.e12. [Google Scholar] [CrossRef]
- Chen, Q.; Stone, P.R.; McCowan, L.M.; Chamley, L. Phagocytosis of Necrotic but Not Apoptotic Trophoblasts Induces Endothelial Cell Activation. Hypertension 2006, 47, 116–121. [Google Scholar] [CrossRef]
- Shen, F.; Wei, J.; Snowise, S.; DeSousa, J.; Stone, P.; Viall, C.; Chen, Q.; Chamley, L. Trophoblast debris extruded from preeclamptic placentae activates endothelial cells: A mechanism by which the placenta communicates with the maternal endothelium. Placenta 2014, 35, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Kandzija, N.; Zhang, W.; Motta-Mejia, C.; Mhlomi, V.; McGowan-Downey, J.; James, T.; Cerdeira, A.S.; Tannetta, D.; Sargent, I.; Redman, C.W.; et al. Placental extracellular vesicles express active dipeptidyl peptidase IV; levels are increased in gestational diabetes mellitus. J. Extracell. Vesicles 2019, 8, 1617000. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, S.; Alam, S.; Jasti, S.; Hodes, H.; Nauser, T.; Gilliam, M.; Billstrand, C.; Hunt, J.; Petroff, M. Immunomodulatory molecules are released from the first trimester and term placenta via exosomes. Placenta 2012, 33, 982–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abumaree, M.; Chamley, L.; Badri, M.; El-Muzaini, M. Trophoblast debris modulates the expression of immune proteins in macrophages: A key to maternal tolerance of the fetal allograft? J. Reprod. Immunol. 2012, 94, 131–141. [Google Scholar] [CrossRef]
- Messerli, K.M.; May, S.R.; Hansson, H.; Schneider, W.; Holzgreve, S.; Hahn, C. Rusterholz, Feto-maternal interactions in preg-nancies: Placental microparticles activate peripheral blood monocytes. Placenta 2010, 31, 106–112. [Google Scholar] [CrossRef]
- Abumaree, M.; Stone, P.; Chamley, L. The effects of apoptotic, deported human placental trophoblast on macrophages: Possible consequences for pregnancy. J. Reprod. Immunol. 2006, 72, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Guo, F.; Jin, H.; Lau, S.; Stone, P.; Chamley, L. Phagocytosis of apoptotic trophoblastic debris protects endothelial cells against activation. Placenta 2012, 33, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Stanley, J.L.; Chen, Q.; Wise, M.R.; Stone, P.R.; James, J.L.; Chamley, L.W. In vivo targets of human placental extracellular vesicles. Reprod. Sci. 2016, 23, 153A. [Google Scholar]
- Chen, Q.; Rutten, V.; Cheng, W.-T.; Tong, M.; Wei, J.; Stone, P.; Ching, L.-M.; Chamley, L.W. Phagocytosis of Extracellular Vesicles Extruded From the Placenta by Ovarian Cancer Cells Inhibits Growth of the Cancer Cells. Int. J. Gynecol. Cancer 2018, 28, 545–552. [Google Scholar] [CrossRef]
- Tong, M.; Kleffmann, T.; Pradhan, S.; Johansson, C.L.; DeSousa, J.; Stone, P.R.; James, J.; Chen, Q.; Chamley, L. Proteomic characterization of macro-, micro- and nano-extracellular vesicles derived from the same first trimester placenta: Relevance for feto-maternal communication. Hum. Reprod. 2016, 31, 687–699. [Google Scholar] [CrossRef]
- Wei, J.; Lau, S.Y.; Blenkiron, C.; Chen, Q.; James, J.L.; Kleffmann, T.; Wise, M.; Stone, P.R.; Chamley, L.W. Trophoblastic debris modifies endothelial cell transcriptome in vitro: A mechanism by which fetal cells might control maternal responses to preg-nancy. Sci. Rep. 2016, 6, 30632. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Kang, M.; Wang, J.; Blenkiron, C.; Lee, A.; Wise, M.; Chamley, L.; Chen, Q. Estimation of the burden of human placental micro- and nano-vesicles extruded into the maternal blood from 8 to 12 weeks of gestation. Placenta 2018, 72–73, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.F.; Kothari, D. Comparison of antibiotic discs from different sources. J. Clin. Pathol. 1975, 28, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, W.; Gomez-Lopez, N.; Erez, O.; Romero, R.; Margolis, L. Extracellular vesicles generated by placental tissues ex vivo: A transport system for immune mediators and growth factors. Am. J. Reprod. Immunol. 2018, 80, e12860. [Google Scholar] [CrossRef] [PubMed]
- Stenqvist, A.-C.; Nagaeva, O.; Baranov, V.; Mincheva-Nilsson, L. Exosomes Secreted by Human Placenta Carry Functional Fas Ligand and TRAIL Molecules and Convey Apoptosis in Activated Immune Cells, Suggesting Exosome-Mediated Immune Privilege of the Fetus. J. Immunol. 2013, 191, 5515–5523. [Google Scholar] [CrossRef] [Green Version]
- Peiris, H.; Vaswani, K.; Almughlliq, F.; Koh, Y.; Mitchell, M. Review: Eicosanoids in preterm labor and delivery: Potential roles of exosomes in eicosanoid functions. Placenta 2017, 54, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ermini, L.; Ausman, J.; Melland-Smith, M.; Yeganeh, B.; Rolfo, A.; Litvack, M.L.; Todros, T.; Letarte, M.; Post, M.; Caniggia, I. A Single Sphingomyelin Species Promotes Exosomal Release of Endoglin into the Maternal Circulation in Preeclampsia. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Delorme-Axford, E.; Donker, R.B.; Mouillet, J.-F.; Chu, T.; Bayer, A.; Ouyang, Y.; Wang, T.; Stolz, D.B.; Sarkar, S.; Morelli, A.E.; et al. Human placental trophoblasts confer viral resistance to recipient cells. Proc. Natl. Acad. Sci. USA 2013, 110, 12048–12053. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Genet. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Scott, M.; Gunderson, C.W.; Mateescu, E.M.; Zhang, Z.; Hwa, T. Interdependence of Cell Growth and Gene Expression: Origins and Consequences. Science 2010, 330, 1099–1102. [Google Scholar] [CrossRef]
- Poehlsgaard, J.; Douthwaite, S. The bacterial ribosome as a target for antibiotics. Nat. Rev. Genet. 2005, 3, 870–881. [Google Scholar] [CrossRef]
- Vuilleumier, M.P.S.; Vuilleumier, S.; Pagni, M. The elusive roles of bacterial glutathione S- transferases: New lessons from genomes. Appl. Microbiol. Biotechnol. 2002, 58, 138–146. [Google Scholar] [CrossRef]
- Reizer, J.; Saier, M.H.; Deutscher, J.; Grenier, F.; Thompson, J.; Hengstenberg, W.; Dills, S.S. The phosphoenolpyruvate: Sugar phos-photransferase system in gram-positive bacteria: Properties, mechanism, and regulation. Crit. Rev. Microbiol. 1988, 15, 297–338. [Google Scholar] [CrossRef]
- Simon, J.P.; Stalon, V. Enzymes of agmatine degradation and the control of their synthesis in Streptococcus faecalis. J. Bacteriol. 1982, 152, 676–681. [Google Scholar] [CrossRef]
- Martínez-Botella, G.; Breen, J.N.; Duffy, J.E.S.; Dumas, J.; Geng, B.; Gowers, I.K.; Green, O.M.; Guler, S.; Hentemann, M.F.; Hernandez-Juan, F.A.; et al. Discovery of Selective and Potent Inhibitors of Gram-Positive Bacterial Thymidylate Kinase (TMK). J. Med. Chem. 2012, 55, 10010–10021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Antibiotics | Without EVs | With Micro-EVs | With Nano-EVs |
|---|---|---|---|
| Susceptible (Standard) | |||
| Ampicillin (10 µg) (≥24) | 29.3 | 28.3 | 30 |
| Penicillin (15 units) (≥24) | 31.3 | 31.3 | 29.3 |
| Linezolid (30 µg) (≥21) | 30 | 31 | 29 |
| Vancomycin (30 µg) (≥17) | 21 | 21 | 20 |
| Resistant (Standard) | |||
| Erythromycin (15 µg) (≤15) | 8 | 8 | 7.7 |
| Clindamycin (2 µg) (≤15) | 6 | 6 | 8 |
| Name of Antibiotics | Without EVs | With Micro-EVs | With Nano-EVs |
|---|---|---|---|
| Susceptible (Standard) | |||
| Fosfomycin (200 µg) (≥16) | 27 | 27 | 27 |
| Piperacillin/Tazobactam (100/10 µg) (≥21) | 29 | 26 | 27 |
| Meropenem (10 µg) (≥23) | 30 | 29 | 29 |
| Ceftazidime (30 µg) (≥21) | 26.5 | 26.7 | 26.7 |
| Ciprofloxacin (5 µg) (≥26) | 31.2 | 30.7 | 30.7 |
| Amikacin (30 µg) (≥17) | 22.4 | 22.4 | 22 |
| Resistant (Standard) | |||
| Ampicillin (10 µg) (≤13) | 6 | 6 | 6 |
| Piperacillin (100 µg) (≤17) | 10 | 10 | 13 |
| Ampicillin/Sulbactam (10/10 µg) (≤11) | 6 | 7 | 8.5 |
| Ceftriaxone (30 µg) (≤19) | 7.25 | 7.5 | 6.15 |
| E. coli | GBS | |
|---|---|---|
| Without placental EVs | 6 | 6 |
| With placental micro-EVs | 6 | 6 |
| With placental nano-EVs | 6 | 6 |
| Protein Accession | Regulation | Functional Description |
|---|---|---|
| Ribosome | ||
| A0A1C0BEJ0 | down | 50S ribosomal protein L21, rRNA binding |
| A0A4U3JET0 | down | 30S ribosomal protein S1, nucleic acid binding |
| A0A6A4TZH8 | down | 50S ribosomal protein L3, rRNA binding |
| Q3K0C9 | down | 50S ribosomal protein L35, structural constituent of ribosome |
| Q8DYG1 | down | 50S ribosomal protein L11, large ribosomal subunit rRNA binding |
| Q8DZZ2 | down | 30S ribosomal protein S20, rRNA binding |
| R4Z934 | down | 30S ribosomal protein S11, rRNA binding |
| V6YZ47 | down | 50S ribosomal protein L3, rRNA binding |
| V6YZ80 | down | 30S ribosomal protein S13, RNA/tRNA binding |
| V6Z128 | down | 30S ribosomal protein S9, structural constituent of ribosome |
| V6Z135 | down | 30S ribosomal protein S11, rRNA/mRNA 5′-UTR/small ribosomal subunit rRNA binding |
| V6Z275 | down | S1 RNA-binding protein, nucleic acid binding |
| V6Z2C6 | down | 50S ribosomal protein L7/L12, structural constituent of ribosome |
| V6Z6Z4 | down | Uncharacterised protein, nucleic acid binding |
| Q8E3E6 | down | 30S ribosomal protein S7, rRNA binding |
| V6Z0L9 | down | 50S ribosomal protein L10, large ribosomal subunit rRNA binding |
| Glutathione Transferase Activity | ||
| A0A0G2Z569 | up | Glutathione S-transferase, omega |
| S8FQZ0 | up | GST C-terminal domain-containing protein |
| V6Z179 | up | S-transferase |
| Oxidoreductase activity, acting on the CH-NH group of donors, NAD or NADP as acceptor | ||
| A0A0E1EHT5 | up | Nitroreductase |
| Q8E7I3 | up | Pyrroline-5-carboxylate reductase |
| S9AZA4 | up | Bifunctional protein FolD |
| Fatty Acid Biosynthetic Process | ||
| A0A0G9JEY8 | up | 3-oxoacyl-[acyl-carrier-protein] synthase 2 |
| A0A656G1D0 | up | 4′-phosphopantetheinyl transferase |
| Q8E5S2 | up | Uncharacterised protein |
| S8FH53 | up | Biotin carboxyl carrier protein of acetyl-CoA carboxylase |
| S9B2D4 | up | 3-oxoacyl-[acyl-carrier-protein] synthase 2 |
| V6Z187 | up | Biotin carboxylase |
| Protein Accession | Regulation | Functional Description |
|---|---|---|
| A0A0H1L4V8 | down | PTS beta-glucoside transporter subunit IIBCA |
| A0A0H1NG81 | down | Putative galactitol operon regulator (Transcriptional antiterminator), BglG family PTS system, mannitolfructose-specific IIA component |
| A0A0H1U8Y4 | down | Glycerol kinase |
| A0A380IIL8 | down | Carbamate kinase |
| A0A380IY32 | down | PTS system, sucrose-specific IIB component/PTS system, sucrose-specific IIC component/PTS system, sucrose-specific IIA component |
| A0A5N0LHT3 | down | Thymidine kinase |
| A0A656G179 | down | PTS system sucrose-specific IIBC component |
| P65204 | down | Adenylate kinase |
| Q8DX04 | down | Sensor histidine kinase |
| Q8E2P0 | down | Uncharacterised protein |
| V6Z3K8 | down | Thymidylate kinase |
| V6Z7B9 | down | Lipid kinase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Tang, Y.; Sun, X.; Chen, Q.; Peng, Y.; Tsai, C.J.-Y.; Chen, Q. Downregulation of Ribosomal Contents and Kinase Activities Is Associated with the Inhibitive Effect on the Growth of Group B Streptococcus Induced by Placental Extracellular Vesicles. Biology 2021, 10, 664. https://doi.org/10.3390/biology10070664
Gao J, Tang Y, Sun X, Chen Q, Peng Y, Tsai CJ-Y, Chen Q. Downregulation of Ribosomal Contents and Kinase Activities Is Associated with the Inhibitive Effect on the Growth of Group B Streptococcus Induced by Placental Extracellular Vesicles. Biology. 2021; 10(7):664. https://doi.org/10.3390/biology10070664
Chicago/Turabian StyleGao, Jing, Yunhui Tang, Xinyi Sun, Qiujing Chen, Yiqian Peng, Catherine Jia-Yun Tsai, and Qi Chen. 2021. "Downregulation of Ribosomal Contents and Kinase Activities Is Associated with the Inhibitive Effect on the Growth of Group B Streptococcus Induced by Placental Extracellular Vesicles" Biology 10, no. 7: 664. https://doi.org/10.3390/biology10070664
APA StyleGao, J., Tang, Y., Sun, X., Chen, Q., Peng, Y., Tsai, C. J.-Y., & Chen, Q. (2021). Downregulation of Ribosomal Contents and Kinase Activities Is Associated with the Inhibitive Effect on the Growth of Group B Streptococcus Induced by Placental Extracellular Vesicles. Biology, 10(7), 664. https://doi.org/10.3390/biology10070664