
Virulence Determinants of Colistin-Resistant K. pneumoniae High-Risk Clones

 , , , ,
, , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Data Collection
2.2. Microbiological and Molecular Studies
2.3. Phagocytosis Assays
2.4. Detection of Neutrophil Extracellular Traps
2.5. Statistical Analysis
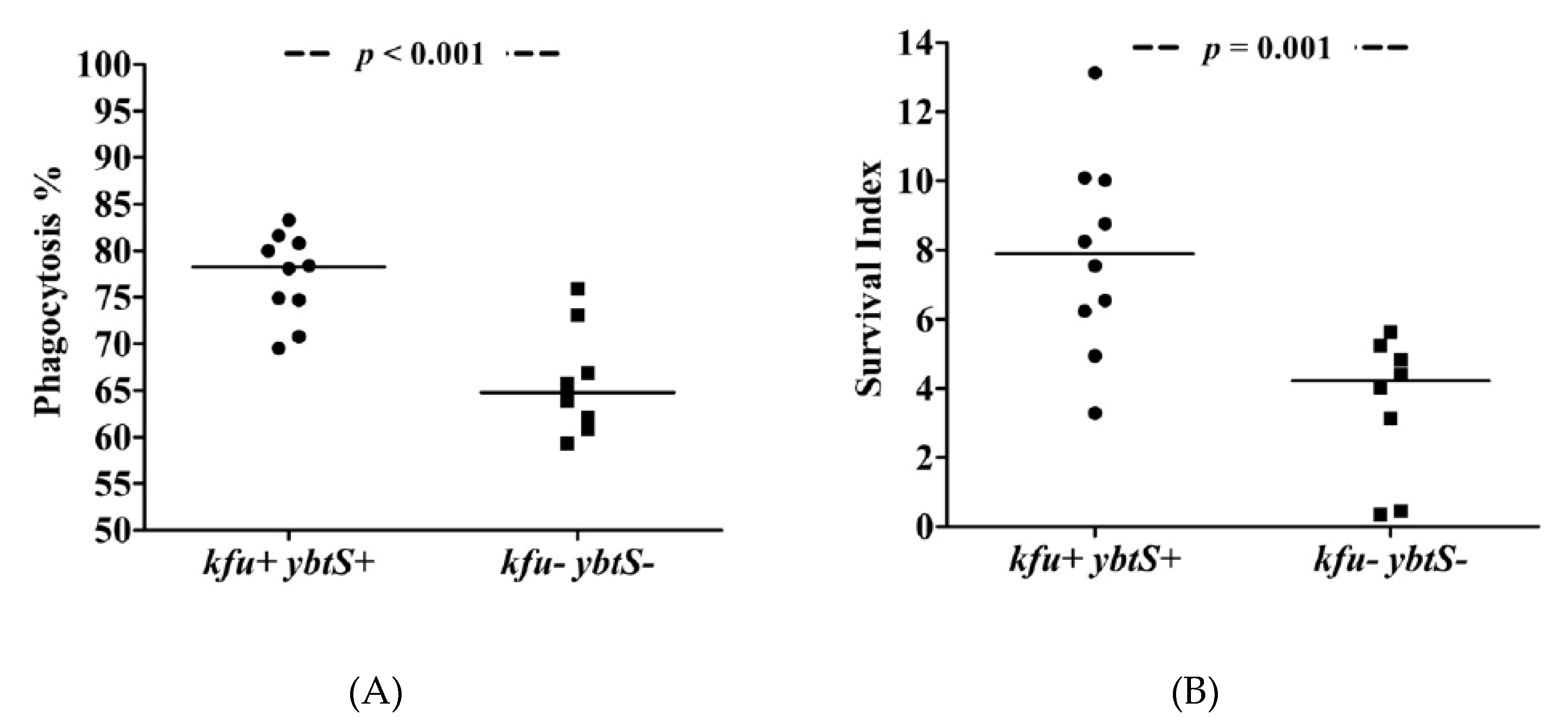
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, C.R.; Lee, J.H.; Park, K.S.; Jeon, J.H.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Antimicrobial Resistance of Hypervirulent Klebsiella pneumoniae: Epidemiology, Hypervirulence-Associated Determinants, and Resistance Mechanisms. Front. Cell Infect. Microbiol. 2017, 7, 483. [Google Scholar] [CrossRef]
- Lawlor, M.S.; O’Connor, C.; Miller, V.L. Yersiniabactin is a virulence factor for Klebsiella pneumoniae during pulmonary infection. Infect. Immun. 2007, 75, 1463–1472. [Google Scholar] [CrossRef]
- Di Tella, D.; Tamburro, M.; Guerrizio, G.; Fanelli, I.; Sammarco, M.L.; Ripabelli, G. Molecular Epidemiological Insights into Colistin-Resistant and Carbapenemases-Producing Clinical Klebsiella pneumoniae Isolates. Infect. Drug Resist. 2019, 12, 3783–3795. [Google Scholar] [CrossRef]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Mammina, C.; Bonura, C.; Aleo, A.; Fasciana, T.; Brunelli, T.; Pesavento, G.; Degl’Innocenti, R.; Nastasi, A. Sequence type 101 (ST101) as the predominant carbapenem-non-susceptible Klebsiella pneumoniae clone in an acute general hospital in Italy. Int. J. Antimicrob. Agents 2012, 39, 543–545. [Google Scholar] [CrossRef]
- Del Franco, M.; Paone, L.; Novati, R.; Giacomazzi, C.G.; Bagattini, M.; Galotto, C.; Montanera, P.G.; Triassi, M.; Zarrilli, R. Molecular epidemiology of carbapenem resistant Enterobacteriaceae in Valle d’Aosta region, Italy, shows the emergence of KPC-2 producing Klebsiella pneumoniae clonal complex 101 (ST101 and ST1789). BMC Microbiol. 2015, 15, 260. [Google Scholar] [CrossRef]
- Can, F.; Menekse, S.; Ispir, P.; Atac, N.; Albayrak, O.; Demir, T.; Karaaslan, D.C.; Karahan, S.N.; Kapmaz, M.; Kurt Azap, O.; et al. Impact of the ST101 clone on fatality among patients with colistin-resistant Klebsiella pneumoniae infection. J. Antimicrob. Chemother. 2018, 73, 1235–1241. [Google Scholar] [CrossRef]
- Avgoulea, K.; Di Pilato, V.; Zarkotou, O.; Sennati, S.; Politi, L.; Cannatelli, A.; Themeli-Digalaki, K.; Giani, T.; Tsakris, A.; Rossolini, G.M.; et al. Characterization of Extensively Drug-Resistant or Pandrug-Resistant Sequence Type 147 and 101 OXA-48-Producing Klebsiella pneumoniae Causing Bloodstream Infections in Patients in an Intensive Care Unit. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Aydin, M.; Ergonul, O.; Azap, A.; Bilgin, H.; Aydin, G.; Cavus, S.A.; Demiroglu, Y.Z.; Aliskan, H.E.; Memikoglu, O.; Menekse, S.; et al. Rapid emergence of colistin resistance and its impact on fatality among healthcare-associated infections. J. Hosp. Infect. 2018, 98, 260–263. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Oteo, J.; Perez-Vazquez, M.; Bautista, V.; Ortega, A.; Zamarron, P.; Saez, D.; Fernandez-Romero, S.; Lara, N.; Ramiro, R.; Aracil, B.; et al. The spread of KPC-producing Enterobacteriaceae in Spain: WGS analysis of the emerging high-risk clones of Klebsiella pneumoniae ST11/KPC-2, ST101/KPC-2 and ST512/KPC-3. J. Antimicrob. Chemother. 2016, 71, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Wand, M.E.; Bock, L.J.; Sutton, J.M. Retention of virulence following colistin adaptation in Klebsiella pneumoniae is strain-dependent rather than associated with specific mutations. J. Med. Microbiol. 2017, 66, 959–964. [Google Scholar] [CrossRef]
- Tzouvelekis, L.S.; Miriagou, V.; Kotsakis, S.D.; Spyridopoulou, K.; Athanasiou, E.; Karagouni, E.; Tzelepi, E.; Daikos, G.L. KPC-producing, multidrug-resistant Klebsiella pneumoniae sequence type 258 as a typical opportunistic pathogen. Antimicrob. Agents Chemother. 2013, 57, 5144–5146. [Google Scholar] [CrossRef]
- Holden, V.I.; Breen, P.; Houle, S.; Dozois, C.M.; Bachman, M.A. Klebsiella pneumoniae Siderophores Induce Inflammation, Bacterial Dissemination, and HIF-1alpha Stabilization during Pneumonia. MBio 2016, 7. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immun. Mol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef]
- Kobayashi, S.D.; Porter, A.R.; Dorward, D.W.; Brinkworth, A.J.; Chen, L.; Kreiswirth, B.N.; DeLeo, F.R. Phagocytosis and Killing of Carbapenem-Resistant ST258 Klebsiella pneumoniae by Human Neutrophils. J. Infect. Dis. 2016, 213, 1615–1622. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informationtional Supplement; CLSI Document M100-S20; Clinical and Laboratory Standards Institute: Pennsylvania, PA, USA, 2010; ISBN 1-56238-716-2. [Google Scholar]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Compain, F.; Babosan, A.; Brisse, S.; Genel, N.; Audo, J.; Ailloud, F.; Kassis-Chikhani, N.; Arlet, G.; Decre, D. Multiplex PCR for detection of seven virulence factors and K1/K2 capsular serotypes of Klebsiella pneumoniae. J. Clin. Microbiol. 2014, 52, 4377–4380. [Google Scholar] [CrossRef]
- Turton, J.F.; Perry, C.; Elgohari, S.; Hampton, C.V. PCR characterization and typing of Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and virulence gene targets. J. Med. Microbiol. 2010, 59, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.L.; Ko, W.C.; Cheng, K.C.; Lee, H.C.; Ke, D.S.; Lee, C.C.; Fung, C.P.; Chuang, Y.C. Association between rmpA and magA genes and clinical syndromes caused by Klebsiella pneumoniae in Taiwan. Clin. Infect. Dis. 2006, 42, 1351–1358. [Google Scholar] [CrossRef]
- Carniel, E. The Yersinia high-pathogenicity island: An iron-uptake island. Microbes. Infect. 2001, 3, 561–569. [Google Scholar] [CrossRef]
- Behnsen, J.; Raffatellu, M. Siderophores: More than Stealing Iron. MBio 2016, 7. [Google Scholar] [CrossRef]
- Bachman, M.A.; Oyler, J.E.; Burns, S.H.; Caza, M.; Lepine, F.; Dozois, C.M.; Weiser, J.N. Klebsiella pneumoniae Yersiniabactin Promotes Respiratory Tract Infection through Evasion of Lipocalin 2. Infect. Immun. 2011, 79, 3309–3316. [Google Scholar] [CrossRef]
- Gomez-Simmonds, A.; Uhlemann, A.C. Clinical Implications of Genomic Adaptation and Evolution of Carbapenem-Resistant Klebsiella pneumoniae. J. Infect. Dis. 2017, 215, S18–S27. [Google Scholar] [CrossRef]
- Shon, A.S.; Russo, T.A. Hypervirulent Klebsiella pneumoniae: The next superbug? Future Microbiol. 2012, 7, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Paauw, A.; Leverstein-van Hall, M.A.; van Kessel, K.P.; Verhoef, J.; Fluit, A.C. Yersiniabactin reduces the respiratory oxidative stress response of innate immune cells. PLoS ONE 2009, 4, e8240. [Google Scholar] [CrossRef]
- Chaturvedi, K.S.; Hung, C.S.; Giblin, D.E.; Urushidani, S.; Austin, A.M.; Dinauer, M.C.; Henderson, J.P. Cupric yersiniabactin is a virulence-associated superoxide dismutase mimic. ACS Chem. Biol. 2014, 9, 551–561. [Google Scholar] [CrossRef]
- Menegazzi, R.; Decleva, E.; Dri, P. Killing by neutrophil extracellular traps: Fact or folklore? Blood 2012, 119, 1214–1216. [Google Scholar] [CrossRef]
- Branzk, N.; Lubojemska, A.; Hardison, S.E.; Wang, Q.; Gutierrez, M.G.; Brown, G.D.; Papayannopoulos, V. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat. Immun. Mol. 2014, 15, 1017–1025. [Google Scholar] [CrossRef]
- Roe, C.C.; Vazquez, A.J.; Esposito, E.P.; Zarrilli, R.; Sahl, J.W. Diversity, Virulence, and Antimicrobial Resistance in Isolates from the Newly Emerging Klebsiella pneumoniae ST101 Lineage. Front. Microbiol. 2019, 10, 542. [Google Scholar] [CrossRef]
- Muggeo, A.; Guillard, T.; Klein, F.; Reffuveille, F.; Francois, C.; Babosan, A.; Bajolet, O.; Bertrand, X.; de Champs, C.; Grp, C. Spread of Klebsiella pneumoniae ST395 non-susceptible to carbapenems and resistant to fluoroquinolones in North-Eastern France. J. Glob. Antimicrob. Res. 2018, 13, 98–103. [Google Scholar] [CrossRef]
- Maida, C.M.; Bonura, C.; Geraci, D.M.; Graziano, G.; Carattoli, A.; Rizzo, A.; Torregrossa, M.V.; Vecchio, D.; Giuffre, M. Outbreak of ST395 KPC-Producing Klebsiella pneumoniae in a Neonatal Intensive Care Unit in Palermo, Italy. Infect. Cont. Hosp. Ep. 2018, 39, 496–498. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Patient | Total (n = 142) n (%) | ST101 n = 80 n (%) | ST395 n = 16 n (%) | Others * (n = 46) n (%) |
|---|---|---|---|---|
| Age Median (range) | 61 (0–91) | 63 (0–86) | 62 (30–84) | 53 (0–91) |
| Female gender | 60 (42) | 34 (43) | 7 (44) | 19 (41) |
| Bacteremia | 61 (43) | 33 (41) p = 0.852 | 10 (63) p = 0.147 | 18 (39) |
| VAP | 67 (47) | 44 (55) p = 0.009 | 9 (56) p = 0.079 | 14 (30) |
| 30-day mortality | 72 (51) | 46 (58) p = 0.005 | 12 (75) p = 0.003 | 14 (30) |
| Being in ICU | 119 (84) | 68 (86) p = 0.323 | 15 (94) p = 0.261 | 36 (78) |
| Mucoid Type and Capsule n (%) | Iron Metabolism n (%) | Adhesins n (%) | LPS n (%) | Allantoin n (%) | Carbapenemase n (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rmpA | magA | K2wzy | K5wzx | fyuA | kfu | IutA | ybtS | entB | mrkD | fimH | wabG | allS | OXA-48 | NDM-1 | |
| ST101 n = 80 | 71 (89) | 6 (8) | 31 (39) | 0 | 79 (99) | 79 (99) | 4 (5) | 79 (99) | 80 (100) | 79 (99) | 79 (99) | 80 (100) | 0 | 76 (95) | 3 (4) |
| p | 0.005 | 1 | 0.707 | - | 0.553 | <0.001 | 0.285 | <0.001 | 0.059 | 0.024 | 0.365 | - | p = 0.00 | p < 0.001 | |
| ST395 n = 16 | 14 (88) | 0 | 9 (56) | 0 | 15 (94) | 15 (94) | 1 (6) | 15 (94) | 15 (100) | 15 (94) | 16 (100) | 16 (100) | 0 | 5 (31) | 14 (88) |
| p | 0.194 | 0.565 | 0.401 | - | 1 | 0.012 | 1 | 0.012 | 1 | 0.315 | 1 | - | p = 0.002 | p < 0.001 | |
| Others n = 46 * | 32 (68) | 23 (49) | 20 (43) | 0 | 45 (96) | 28 (60) | 5 (11) | 28 (60) | 47 (100) | 43 (92) | 42 (89) | 46 (98) | 0 | 36 (77) | 14 (30) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dogan, O.; Vatansever, C.; Atac, N.; Albayrak, O.; Karahuseyinoglu, S.; Sahin, O.E.; Kilicoglu, B.K.; Demiray, A.; Ergonul, O.; Gönen, M.; et al. Virulence Determinants of Colistin-Resistant K. pneumoniae High-Risk Clones. Biology 2021, 10, 436. https://doi.org/10.3390/biology10050436
Dogan O, Vatansever C, Atac N, Albayrak O, Karahuseyinoglu S, Sahin OE, Kilicoglu BK, Demiray A, Ergonul O, Gönen M, et al. Virulence Determinants of Colistin-Resistant K. pneumoniae High-Risk Clones. Biology. 2021; 10(5):436. https://doi.org/10.3390/biology10050436
Chicago/Turabian StyleDogan, Ozlem, Cansel Vatansever, Nazli Atac, Ozgur Albayrak, Sercin Karahuseyinoglu, Ozgun Ekin Sahin, Bilge Kaan Kilicoglu, Atalay Demiray, Onder Ergonul, Mehmet Gönen, and et al. 2021. "Virulence Determinants of Colistin-Resistant K. pneumoniae High-Risk Clones" Biology 10, no. 5: 436. https://doi.org/10.3390/biology10050436
APA StyleDogan, O., Vatansever, C., Atac, N., Albayrak, O., Karahuseyinoglu, S., Sahin, O. E., Kilicoglu, B. K., Demiray, A., Ergonul, O., Gönen, M., & Can, F. (2021). Virulence Determinants of Colistin-Resistant K. pneumoniae High-Risk Clones. Biology, 10(5), 436. https://doi.org/10.3390/biology10050436