
Assessment of 16S rRNA Gene-Based Phylogenetic Diversity of Archaeal Communities in Halite-Crystal Salts Processed from Natural Saharan Saline Systems of Southern Tunisia


 , ,
, , 
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Site Description and Sampling
2.2. DNA Extraction from Environmental Samples and Archaeal Isolates
2.3. Isolation of Halophilic Archaea, PCR Amplification, and 16S rRNA Gene Sequencing
2.4. DGGE Fingerprinting
2.5. Full-Length 16S rRNA Archaeal Libraries
2.6. Sequence Analysis and Phylogenetic Assessment of Archaeal 16SrRNA Gene Sequences of Isolates and Clone Libraries
2.7. Illumina MiSeq Sequencing, Data Analysis, and Diversity Estimates
3. Nucleotide Sequence Accession Numbers
4. Results and Discussion
4.1. Isolation and Identification of Haloarchaeal Isolates
4.2. Total Archaeal Community Assessment
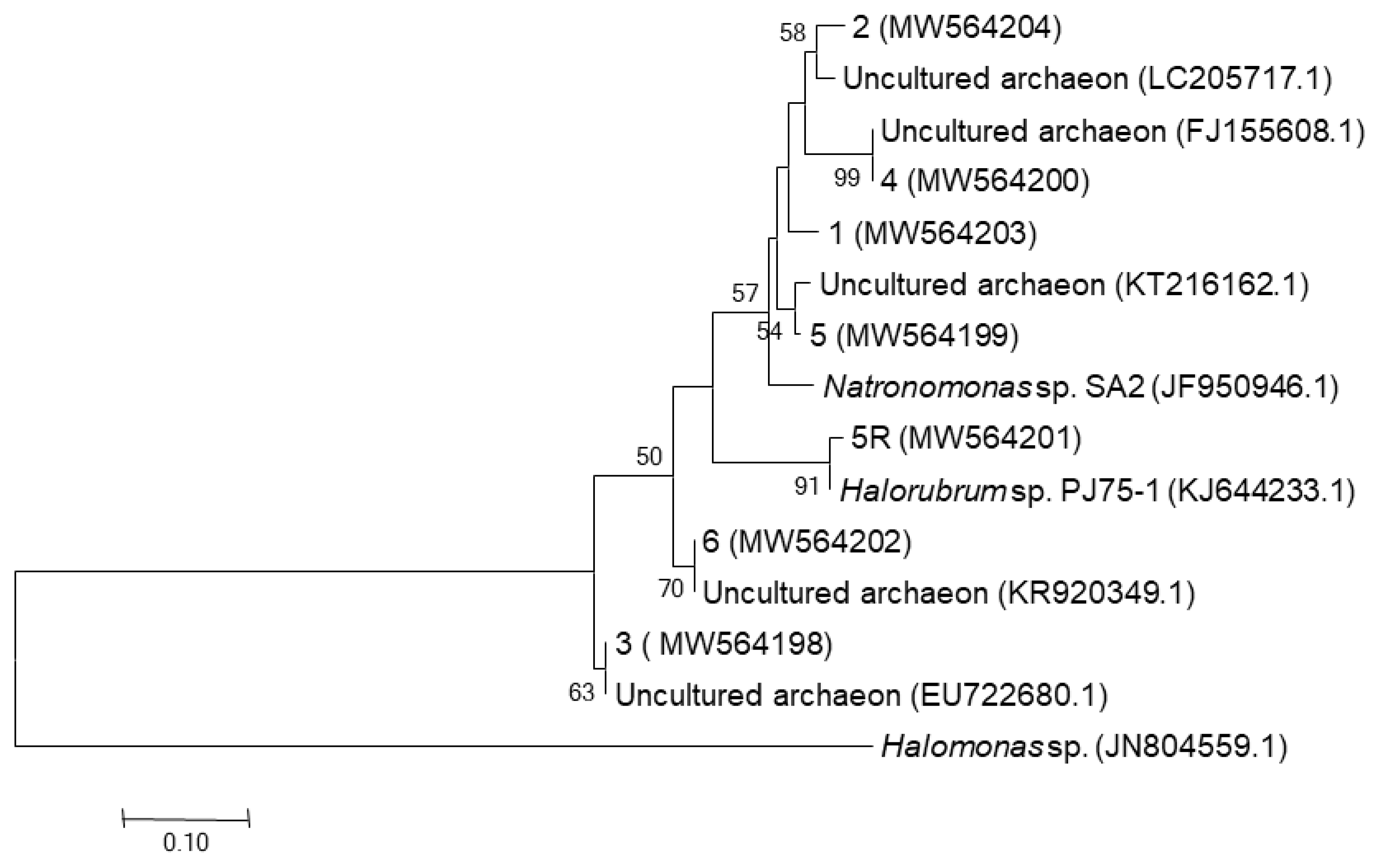
4.2.1. DGGE Fingerprint
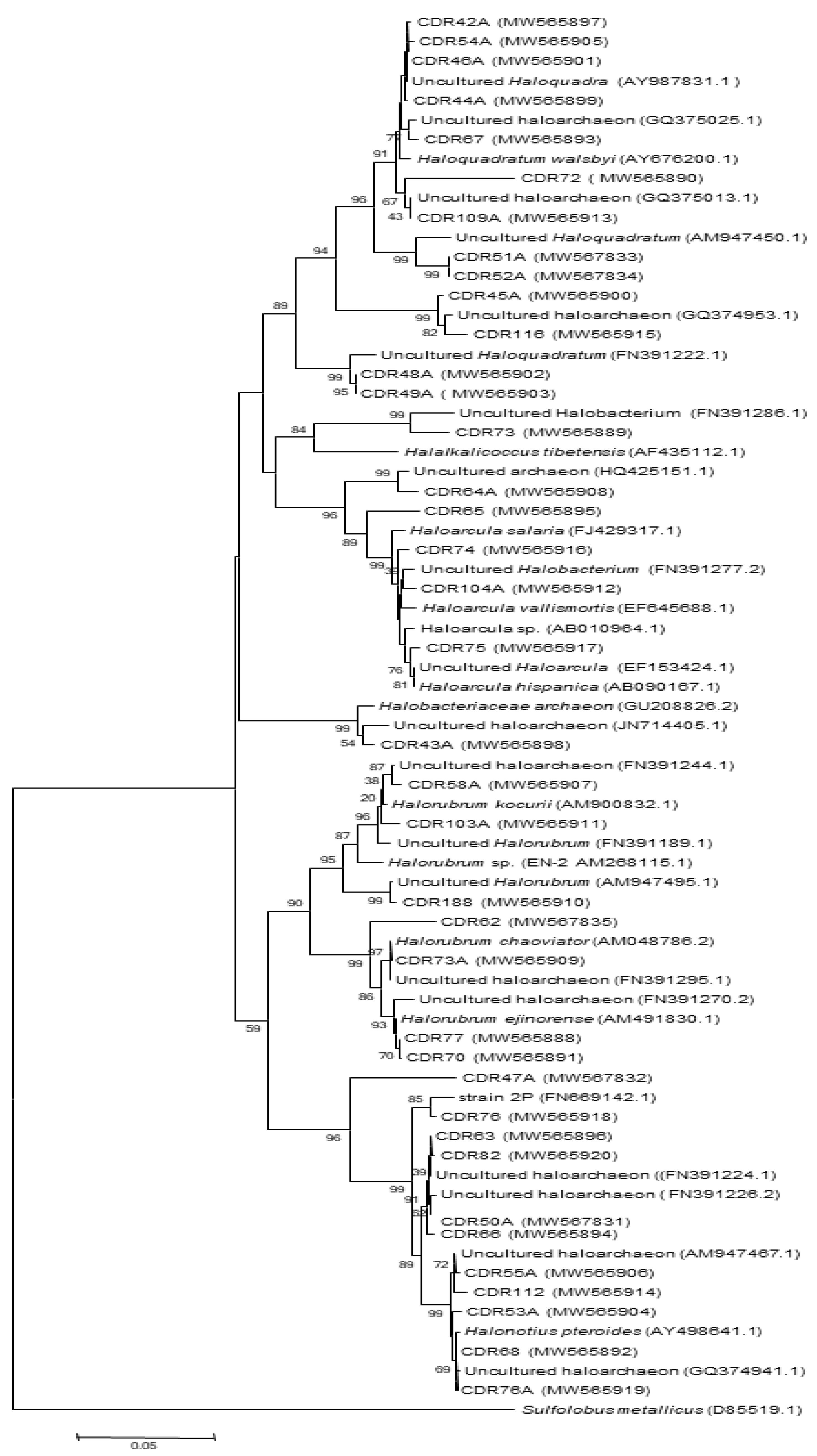
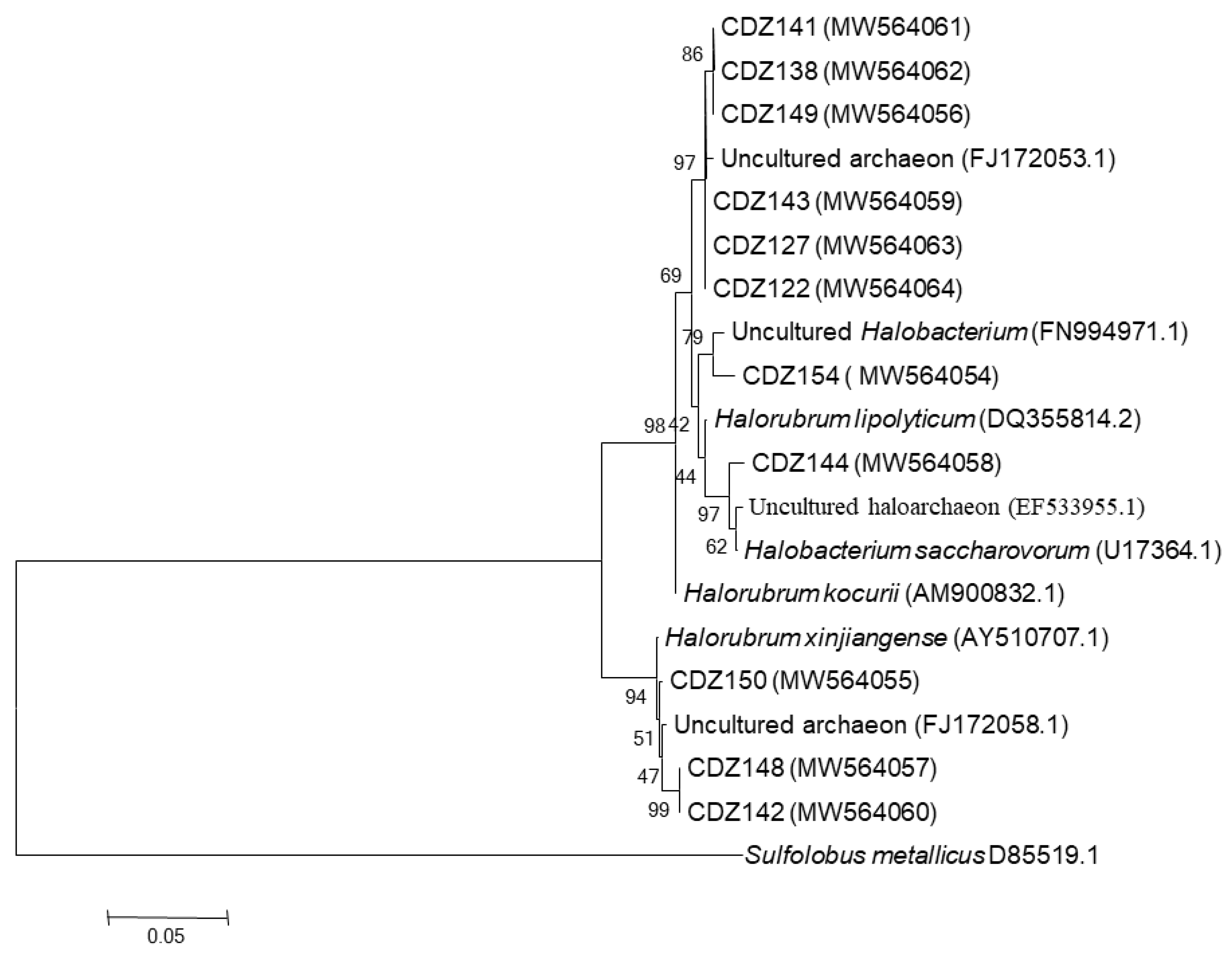
4.2.2. Full Length 16S rRNA Gene Libraries
Diversity and Richness Estimates
Taxonomic Assignment, Quantification, and Distribution
4.2.3. Archaeal Profiling Using MiSeq-Based High-Throughput 16S rRNA Sequencing Approach
Diversity and Richness Estimates
Class and Genus Levels Distribution
5. Haloarchaeal Community Structure between Sites Based on All Approaches
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stivaletta, N.; Barbieri, R.; Picard, C.; Bosco, M. Astrobiological Significance of the Sabkha Life and Environments of Southern Tunisia. Planet. Space Sci. 2009, 57, 597–605. [Google Scholar] [CrossRef]
- Javor, B. Hypersaline Environments Microbiology and Biogeochemistry; Springer: Berlin/Heidelberg, Germany, 1989; ISBN 978-3-642-74370-2. [Google Scholar]
- Kamekura, M. Diversity of Extremely Halophilic Bacteria. Extremophiles 1998, 2, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Halophilic Microorganisms and Their Environments; Springer: Berlin/Heidelberg, Germany, 2002; ISBN 978-0-306-48053-9. [Google Scholar]
- DasSarma, S.; DasSarma, P. Halophiles. In eLS; John Wiley & Sons, Ltd., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2012; p. a0000394.pub3. ISBN 978-0-470-01617-6. [Google Scholar]
- Youssef, N.H.; Savage-Ashlock, K.N.; McCully, A.L.; Luedtke, B.; Shaw, E.I.; Hoff, W.D.; Elshahed, M.S. Trehalose/2-Sulfotrehalose Biosynthesis and Glycine-Betaine Uptake Are Widely Spread Mechanisms for Osmoadaptation in the Halobacteriales. ISME J. 2014, 8, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Najjari, A.; Elshahed, M.S.; Cherif, A.; Youssef, N.H. Patterns and Determinants of Halophilic Archaea (Class Halobacteria) Diversity in Tunisian Endorheic Salt Lakes and Sebkhet Systems. Appl. Environ. Microbiol. 2015, 81, 4432–4441. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Siroosi, M.; Atashgahi, S.; Smidt, H.; Ventosa, A. Systematics of Haloarchaea and Biotechnological Potential of Their Hydrolytic Enzymes. Microbiology 2017, 163, 623–645. [Google Scholar] [CrossRef]
- Durán-Viseras, A.; Andrei, A.-S.; Ghai, R.; Sánchez-Porro, C.; Ventosa, A. New Halonotius Species Provide Genomics-Based Insights into Cobalamin Synthesis in Haloarchaea. Front. Microbiol. 2019, 10, 1928. [Google Scholar] [CrossRef]
- Grant, W.D.; Gemmell, R.T.; McGenity, T.J. Halobacteria: The Evidence for Longevity. Extremophiles 1998, 2, 279–287. [Google Scholar] [CrossRef]
- Gibbons, N.E. Halobacteriaceae. In Bergey’s Manual of Determinative Bacteriology; Buchanan, R.E., Gibbons, N.E., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1974; pp. 269–273. [Google Scholar]
- Anton, J.; Llobet-Brossa, E.; Rodriguez-Valera, F.; Amann, R. Fluorescence in Situ Hybridization Analysis of the Prokaryotic Community Inhabiting Crystallizer Ponds. Environ. Microbiol. 1999, 1, 517–523. [Google Scholar] [CrossRef]
- Ventosa, A. Halophilic Microorganisms; Springer: Berlin/Heidelberg, Germany, 2004; ISBN 978-3-662-07656-9. [Google Scholar]
- Oren, A.; Litchfield, C.D. A Procedure for the Enrichment and Isolation of Halobacterium. FEMS Microbiol. Lett. 1999, 173, 353–358. [Google Scholar] [CrossRef]
- Maturrano, L.; Santos, F.; Rosselló-Mora, R.; Antón, J. Microbial Diversity in Maras Salterns, a Hypersaline Environment in the Peruvian Andes. Appl. Environ. Microbiol. 2006, 72, 3887–3895. [Google Scholar] [CrossRef]
- George, R.Y. Extremophilic Life in Our Oceans as Models for Astrobiology. In Extremophiles as Astrobiological Models; Seckbach, J., Stan-Lotter, H., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 161–182. ISBN 978-1-119-59309-6. [Google Scholar]
- Huby, T.J.C.; Clark, D.R.; McKew, B.A.; McGenity, T.J. Extremely Halophilic Archaeal Communities Are Resilient to Short-term Entombment in Halite. Environ. Microbiol. 2020, 1462–2920. [Google Scholar] [CrossRef] [PubMed]
- Benlloch, S.; Acinas, S.G.; Martinez-Murcia, A.J.; Rodreguez-Valera, F. Description of Prokaryotic Biodiversity along the Salinity Gradient of a Multipond Solar Saltern by Direct PCR Amplification of 16S RDNA. Hydrobiologia 1996, 329, 19–31. [Google Scholar] [CrossRef]
- Javor, B. Industrial Microbiology of Solar Salt Production. J. Ind. Microbiol. Biotechnol. 2002, 28, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Tsiamis, G.; Katsaveli, K.; Ntougias, S.; Kyrpides, N.; Andersen, G.; Piceno, Y.; Bourtzis, K. Prokaryotic Community Profiles at Different Operational Stages of a Greek Solar Saltern. Res. Microbiol. 2008, 159, 609–627. [Google Scholar] [CrossRef]
- Baati, H.; Guermazi, S.; Amdouni, R.; Gharsallah, N.; Sghir, A.; Ammar, E. Prokaryotic Diversity of a Tunisian Multipond Solar Saltern. Extremophiles 2008, 12, 505–518. [Google Scholar] [CrossRef]
- Trigui, H.; Masmoudi, S.; Brochier-Armanet, C.; Maalej, S.; Dukan, S. Characterization of Halorubrum Sfaxense Sp. Nov., a New Halophilic Archaeon Isolated from the Solar Saltern of Sfax in Tunisia. Int. J. Microbiol. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boujelben, I.; Gomariz, M.; Martínez-García, M.; Santos, F.; Peña, A.; López, C.; Antón, J.; Maalej, S. Spatial and Seasonal Prokaryotic Community Dynamics in Ponds of Increasing Salinity of Sfax Solar Saltern in Tunisia. Antonie Leeuwenhoek 2012, 101, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Baricz, A.; Coman, C.; Andrei, A.Ş.; Muntean, V.; Keresztes, Z.G.; Păuşan, M.; Alexe, M.; Banciu, H.L. Spatial and Temporal Distribution of Archaeal Diversity in Meromictic, Hypersaline Ocnei Lake (Transylvanian Basin, Romania). Extremophiles 2014, 18, 399–413. [Google Scholar] [CrossRef]
- Oren, A. Life at High Salt Concentrations, Intracellular KCl Concentrations, and Acidic Proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef]
- Lefond, S.J. Handbook of World Salt Resources; Plenum Press: New York, NY, USA, 1969. [Google Scholar]
- Norton, C.F.; Grant, W.D. Survival of Halobacteria within Fluid Inclusions in Salt Crystals. Microbiology 1988, 134, 1365–1373. [Google Scholar] [CrossRef]
- Fendrihan, S.; Legat, A.; Pfaffenhuemer, M.; Gruber, C.; Weidler, G.; Gerbl, F.; Stan-Lotter, H. Extremely Halophilic Archaea and the Issue of Long-Term Microbial Survival. Rev. Environ. Sci. Biotechnol. 2006, 5, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Schubert, B.A.; Lowenstein, T.K.; Timofeeff, M.N.; Parker, M.A. Halophilic Archaea Cultured from Ancient Halite, Death Valley, California. Environ. Microbiol. 2010, 12, 440–454. [Google Scholar] [CrossRef]
- Schubert, B.A.; Lowenstein, T.K.; Timofeeff, M.N. Microscopic Identification of Prokaryotes in Modern and Ancient Halite, Saline Valley and Death Valley, California. Astrobiology 2009, 9, 467–482. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P.; Laye, V.J.; Schwieterman, E.W. Extremophilic Models for Astrobiology: Haloarchaeal Survival Strategies and Pigments for Remote Sensing. Extremophiles 2020, 24, 31–41. [Google Scholar] [CrossRef]
- DasSarma, S. Extreme halophiles are models for Astrobiology. Microbe 2006, 1, 120–126. [Google Scholar] [CrossRef][Green Version]
- Oren, A.; Pri-El, N.; Shapiro, O.; Siboni, N. Buoyancy studies in natural communities of square gas-vacuolate archaea in saltern crystallizer ponds. Saline Syst. 2006, 2, 4. [Google Scholar] [CrossRef]
- Soppa, J. From Genomes to Function: Haloarchaea as Model Organisms. Microbiology 2006, 152, 585–590. [Google Scholar] [CrossRef]
- Kharroub, K.; Quesada, T.; Ferrer, R.; Fuentes, S.; Aguilera, M.; Boulahrouf, A.; Ramos-Cormenzana, A.; Monteoliva-Sánchez, M. Halorubrum ezzemoulense Sp. Nov., a Halophilic Archaeon Isolated from Ezzemoul Sabkha, Algeria. Int. J. Syst. Evol. Microbiol. 2006, 56, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Kharroub, K.; Lizama, C.; Aguilera, M.; Boulahrouf, A.; Campos, V.; Ramos-Cormenzana, A.; Monteoliva-Sanchez, M. Halomicrobium katesii Sp. Nov., an Extremely Halophilic Archaeon. Int. J. Syst. Evol. Microbiol. 2008, 58, 2354–2358. [Google Scholar] [CrossRef] [PubMed]
- Luque, R.; González-Domenech, C.M.; Llamas, I.; Quesada, E.; Béjar, V. Diversity of Culturable Halophilic Archaea Isolated from Rambla Salada, Murcia (Spain). Extremophiles 2012, 16, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial Diversity and Biogeochemical Cycling in Soda Lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef]
- Williams, T.J.; Allen, M.A.; DeMaere, M.Z.; Kyrpides, N.C.; Tringe, S.G.; Woyke, T.; Cavicchioli, R. Microbial Ecology of an Antarctic Hypersaline Lake: Genomic Assessment of Ecophysiology among Dominant Haloarchaea. ISME J. 2014, 8, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, H.; Mossel, D.A.A.; Edberg, S.C.; Struijk, C.B. Advances in the Bacteriology of the Coliform Group: Their Suitability as Markers of Microbial Water Safety. Annu. Rev. Microbiol. 2001, 55, 201–234. [Google Scholar] [CrossRef]
- Youssef, N.H.; Ashlock-Savage, K.N.; Elshahed, M.S. Phylogenetic Diversities and Community Structure of Members of the Extremely Halophilic Archaea (Order Halobacteriales) in Multiple Saline Sediment Habitats. Appl. Environ. Microbiol. 2012, 78, 1332–1344. [Google Scholar] [CrossRef]
- Henriet, O.; Fourmentin, J.; Delincé, B.; Mahillon, J. Exploring the Diversity of Extremely Halophilic Archaea in Food-Grade Salts. Int. J. Food Microbiol. 2014, 191, 36–44. [Google Scholar] [CrossRef]
- Baati, H.; Guermazi, S.; Gharsallah, N.; Sghir, A.; Ammar, E. Novel Prokaryotic Diversity in Sediments of Tunisian Multipond Solar Saltern. Res. Microbiol. 2010, 161, 573–582. [Google Scholar] [CrossRef]
- Dillon, J.G.; Carlin, M.; Gutierrez, A.; Nguyen, V.; McLain, N. Patterns of Microbial Diversity along a Salinity Gradient in the Guerrero Negro Solar Saltern, Baja CA Sur, Mexico. Front. Microbiol. 2013, 4, 399. [Google Scholar] [CrossRef]
- Benlloch, S.; Acinas, S.G.; Antón, J.; López-López, A.; Luz, S.P.; Rodríguez-Valera, F. Archaeal Biodiversity in Crystallizer Ponds from a Solar Saltern: Culture versus PCR. Microb. Ecol. 2001, 41, 12–19. [Google Scholar] [CrossRef]
- Burns, D.G.; Camakaris, H.M.; Janssen, P.H.; Dyall-Smith, M.L. Combined Use of Cultivation-Dependent and Cultivation-Independent Methods Indicates That Members of Most Haloarchaeal Groups in an Australian Crystallizer Pond Are Cultivable. Appl. Environ. Microbiol. 2004, 70, 5258–5265. [Google Scholar] [CrossRef]
- Nicol, G.W.; Schleper, C. Ammonia-oxidising Crenarchaeota: Important players in the nitrogen cycle? Trends Microbiol. 2006, 14, 207–212. [Google Scholar] [CrossRef]
- Dyall-Smith, M. The Halohandbook: Protocols for Haloarchaeal Genetics. 2012. Available online: http://www.haloarchaea.com/resources/halohandbook/Halohandbook_2009_v7.2mds.pdf (accessed on 16 November 2012).
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Tekerlekopoulou, A.G.; Tsiamis, G.; Dermou, E.; Siozios, S.; Bourtzis, K.; Vayenas, D.V. The Effect of Carbon Source on Microbial Community Structure and Cr(VI) Reduction Rate. Biotechnol. Bioeng. 2010, 107, 478–487. [Google Scholar] [CrossRef]
- Ovreås, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of Bacterioplankton in Meromictic Lake Saelenvannet, as Determined by Denaturing Gradient Gel Electrophoresis of PCR-Amplified Gene Fragments Coding for 16S RRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [CrossRef]
- Chang, H.; Kim, K.; Nam, Y.; Roh, S.; Kim, M.; Jeon, C.; Oh, H.; Bae, J. Analysis of Yeast and Archaeal Population Dynamics in Kimchi Using Denaturing Gradient Gel Electrophoresis. Int. J. Food Microbiol. 2008, 126, 159–166. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of Complex Microbial Populations by Denaturing Gradient Gel Electrophoresis Analysis of Polymerase Chain Reaction-Amplified Genes Coding for 16S RRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef]
- Tajima, K.; Nagamine, T.; Matsui, H.; Nakamura, M.; Aminov, R.I. Phylogenetic Analysis of Archaeal 16S RRNA Libraries from the Rumen Suggests the Existence of a Novel Group of Archaea Not Associated with Known Methanogens. FEMS Microbiol. Lett. 2001, 200, 67–72. [Google Scholar] [CrossRef]
- Hartley, J.L.; Bowen, H. PEG precipitation for selective removal of small DNA fragments. Focus 2003, 25, 18. [Google Scholar]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved Alignments and New Tools for RRNA Analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism; Elsevier: Amsterdam, The Netherlands, 1969; pp. 21–132. ISBN 978-1-4832-3211-9. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for Reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Shahmohammadi, H.R.; Asgarani, E.; Terato, H.; Saito, T.; Ohyama, Y.; Gekko, K.; Yamamoto, O.; Ide, H. Protective Roles of Bacterioruberin and Intracellular KCl in the Resistance of Halobacterium salinarium against DNA-Damaging Agents. JRR 1998, 39, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Baxter, B.K. DNA Repair and Photoprotection: Mechanisms of Overcoming Environmental Ultraviolet Radiation Exposure in Halophilic Archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef]
- Pecher, W.T.; Martínez, F.L.; DasSarma, P.; Guzmán, D.; DasSarma, S. 16S RRNA Gene Diversity in the Salt Crust of Salar de Uyuni, Bolivia, the World’s Largest Salt Flat. Microbiol. Resour. Announc. 2020, 9, e00374-20. [Google Scholar] [CrossRef]
- Oren, A.; Gurevich, P.; Anati, D.A.; Barkan, E.; Luz, B. A Bloom of Dunaliella Parva in the Dead Sea in 1992: Biological and Biogeochemical Aspects. Hydrobiologia 1995, 297, 173–185. [Google Scholar] [CrossRef]
- Echigo, A.; Minegishi, H.; Shimane, Y.; Kamekura, M.; Itoh, T.; Usami, R. Halomicroarcula pellucida Gen. Nov., Sp. Nov., a Non-Pigmented, Transparent-Colony-Forming, Halophilic Archaeon Isolated from Solar Salt. Int. J. Syst. Evol. Microbiol. 2013, 63, 3556–3562. [Google Scholar] [CrossRef]
- Norton, C.F.; McGenity, T.J.; Grant, W.D. Archaeal halophiles (Halobacteria) from two British salt mines. Microbiology 1993, 139, 1077–1081. [Google Scholar] [CrossRef]
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D.; Stan-Lotter, H. Origins of Halophilic Microorganisms in Ancient Salt Deposits. Minireview. Environ. Microbiol. 2000, 2, 243–250. [Google Scholar] [CrossRef]
- Vreeland, R.H.; Jones, J.; Monson, A.; Rosenzweig, W.D.; Lowenstein, T.K.; Timofeeff, M.; Satterfield, C.; Cho, B.C.; Park, J.S.; Wallace, A.; et al. Isolation of Live Cretaceous (121–112 Million Years Old) Halophilic Archaea from Primary Salt Crystals. Geomicrobiol. J. 2007, 24, 275–282. [Google Scholar] [CrossRef]
- Park, J.S.; Vreeland, R.H.; Cho, B.C.; Lowenstein, T.K.; Timofeeff, M.N.; Rosenzweig, W.D. Haloarchaeal Diversity in 23, 121 and 419 MYA Salts. Geobiology 2009, 7, 515–523. [Google Scholar] [CrossRef]
- Lowenstein, T.K.; Schubert, B.A.; Timofeeff, M.N. Microbial Communities in Fluid Inclusions and Long-Term Survival in Halite. GSAT 2011, 21, 4–9. [Google Scholar] [CrossRef]
- Thombre, R.S.; Shinde, V.D.; Oke, R.S.; Dhar, S.K.; Shouche, Y.S. Biology and Survival of Extremely Halophilic Archaeon Haloarcula marismortui RR12 Isolated from Mumbai Salterns, India in Response to Salinity Stress. Sci. Rep. 2016, 6, 25642. [Google Scholar] [CrossRef]
- Gibtan, A.; Park, K.; Woo, M.; Shin, J.-K.; Lee, D.-W.; Sohn, J.H.; Song, M.; Roh, S.W.; Lee, S.-J.; Lee, H.-S. Diversity of Extremely Halophilic Archaeal and Bacterial Communities from Commercial Salts. Front. Microbiol. 2017, 8, 799. [Google Scholar] [CrossRef] [PubMed]
- Mormile, M.R.; Biesen, M.A.; Gutierrez, M.C.; Ventosa, A.; Pavlovich, J.B.; Onstott, T.C.; Fredrickson, J.K. Isolation of Halobacterium salinarum Retrieved Directly from Halite Brine Inclusions. Environ. Microbiol. 2003, 5, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular Signature of Hypersaline Adaptation: Insights from Genome and Proteome Composition of Halophilic Prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [PubMed]
- Tamez, P.; Lopez-Cortés, A. Interaction of Extreme Halophilic Archaea with the Evaporites of the Solar Salterns Guerrero Negro Baja California, Mexico; American Geophysical Union: Washington, DC, USA, 2008. [Google Scholar]
- Xiao, W.; Peng, Q.; Liu, H.; Wen, M.; Cui, X.; Yang, Y.; Duan, D.; Chen, W.; Deng, L.; Li, Q.; et al. Prokaryotic microbial diversity of the ancient salt deposits in the Kunming Salt Mine, China. Acta Microbiol. Sin. 2007, 47, 295–300. [Google Scholar]
- Gramain, A.; Díaz, G.C.; Demergasso, C.; Lowenstein, T.K.; McGenity, T.J. Archaeal Diversity along a Subterranean Salt Core from the Salar Grande (Chile): Archaeal Diversity along a Salt Core. Environ. Microbiol. 2011, 13, 2105–2121. [Google Scholar] [CrossRef]
- Elshahed, M.S.; Savage, K.N.; Oren, A.; Gutierrez, M.C.; Ventosa, A.; Krumholz, L.R. Haloferax sulfurifontis Sp. Nov., a Halophilic Archaeon Isolated from a Sulfide-and Sulfur-Rich Spring. Int. J. Syst. Evol. Microbiol. 2004, 54, 2275–2279. [Google Scholar] [CrossRef]
- Walsh, D.A.; Papke, R.T.; Doolittle, W.F. Archaeal Diversity along a Soil Salinity Gradient Prone to Disturbance. Environ. Microbiol. 2005, 7, 1655–1666. [Google Scholar] [CrossRef]
- Oueriaghli, N.; Béjar, V.; Quesada, E.; Martínez-Checa, F. Molecular Ecology Techniques Reveal Both Spatial and Temporal Variations in the Diversity of Archaeal Communities within the Athalassohaline Environment of Rambla Salada, Spain. Microb. Ecol. 2013, 66, 297–311. [Google Scholar] [CrossRef]
- Oh, D.; Porter, K.; Russ, B.; Burns, D.; Dyall-Smith, M. Diversity of Haloquadratum and Other Haloarchaea in Three, Geographically Distant, Australian Saltern Crystallizer Ponds. Extremophiles 2010, 14, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhaxybayeva, O.; Stepanauskas, R.; Mohan, N.R.; Papke, R.T. Cell Sorting Analysis of Geographically Separated Hypersaline Environments. Extremophiles 2013, 17, 265–275. [Google Scholar] [CrossRef]
- Han, R.; Zhang, X.; Liu, J.; Long, Q.; Chen, L.; Liu, D.; Zhu, D. Microbial Community Structure and Diversity within Hypersaline Keke Salt Lake Environments. Can. J. Microbiol. 2017, 63, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Bidle, K.; Amadio, W.; Oliveira, P.; Paulish, T.; Hicks, S.; Earnest, C. Research Article: A Phylogenetic Analysis of Haloarchaea Found in a Solar Saltern. BIOS 2005, 76, 89–96. [Google Scholar] [CrossRef]
- Bowman, J.P.; McCammon, S.A.; Rea, S.M.; McMeekin, T.A. The Microbial Composition of Three Limnologically Disparate Hypersaline Antarctic Lakes. FEMS Microbiol. Lett. 2000, 183, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Podell, S.; Emerson, J.B.; Jones, C.M.; Ugalde, J.A.; Welch, S.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. Seasonal Fluctuations in Ionic Concentrations Drive Microbial Succession in a Hypersaline Lake Community. ISME J. 2014, 8, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Çınar, S.; Mutlu, M.B. Comparative Analysis of Prokaryotic Diversity in Solar Salterns in Eastern Anatolia (Turkey). Extremophiles 2016, 20, 589–601. [Google Scholar] [CrossRef]
- Burns, D.G.; Janssen, P.H.; Itoh, T.; Kamekura, M.; Li, Z.; Jensen, G.; Rodríguez-Valera, F.; Bolhuis, H.; Dyall-Smith, M.L. Haloquadratum walsbyi Gen. Nov., Sp. Nov., the Square Haloarchaeon of Walsby, Isolated from Saltern Crystallizers in Australia and Spain. Int. J. Syst. Evol. Microbiol. 2007, 57, 387–392. [Google Scholar] [CrossRef]
- Walsby, A.E. A Square Bacterium. Nature 1980, 283, 69–71. [Google Scholar] [CrossRef]
- Bolhuis, H.; Poele, E.M.T.; Rodriguez-Valera, F. Isolation and Cultivation of Walsby’s Square Archaeon. Environ. Microbiol. 2004, 6, 1287–1291. [Google Scholar] [CrossRef]
- Burns, D.; Camakaris, H.; Janssen, P.; Dyallsmith, M. Cultivation of Walsby’S Square Haloarchaeon. FEMS Microbiol. Lett. 2004, 238, 469–473. [Google Scholar] [CrossRef]
- Burns, D.G.; Janssen, P.H.; Itoh, T.; Kamekura, M.; Echigo, A.; Dyall-Smith, M.L. Halonotius pteroides Gen. Nov., Sp. Nov., an Extremely Halophilic Archaeon Recovered from a Saltern Crystallizer. Int. J. Syst. Evol. Microbiol. 2010, 60, 1196–1199. [Google Scholar] [CrossRef][Green Version]
- Oxley, A.P.A.; Lanfranconi, M.P.; Würdemann, D.; Ott, S.; Schreiber, S.; McGenity, T.J.; Timmis, K.N.; Nogales, B. Halophilic Archaea in the Human Intestinal Mucosa: Haloarchaea in the Intestinal Mucosa. Environ. Microbiol. 2010, 12, 2398–2410. [Google Scholar] [CrossRef] [PubMed]
- McGenity, T.J.; Grant, W.D. Transfer of Halobacterium saccharovorum, Halobacterium sodomense, Halobacterium trapanicum NRC 34021 and Halobacterium lacusprofundi to the Genus halorubrum Gen. Nov., as Halorubrum saccharovorum Comb. Nov., Halorubrum sodomense Comb. Nov., Halorubrum trapanicum Comb. Nov., and Halorubrum lacusprofundi Comb Nov. Syst. Appl. Microbiol. 1995, 18, 237–243. [Google Scholar] [CrossRef]
- Hu, L.; Pan, H.; Xue, Y.; Ventosa, A.; Cowan, D.A.; Jones, B.E.; Grant, W.D.; Ma, Y. Halorubrum luteum Sp. Nov., Isolated from Lake Chagannor, Inner Mongolia, China. Int. J. Syst. Evol. Microbiol. 2008, 58, 1705–1708. [Google Scholar] [CrossRef][Green Version]
- Papke, R.; Corral, P.; Ram-Mohan, N.; Haba, R.; Sánchez-Porro, C.; Makkay, A.; Ventosa, A. Horizontal Gene Transfer, Dispersal and Haloarchaeal Speciation. Life 2015, 5, 1405–1426. [Google Scholar] [CrossRef]
- Kemp, B.; Tabish, E.; Wolford, A.; Jones, D.; Butler, J.; Baxter, B. The Biogeography of Great Salt Lake Halophilic Archaea: Testing the Hypothesis of Avian Mechanical Carriers. Diversity 2018, 10, 124. [Google Scholar] [CrossRef]
- Gupta, R.S.; Naushad, S.; Baker, S. Phylogenomic Analyses and Molecular Signatures for the Class Halobacteria and Its Two Major Clades: A Proposal for Division of the Class Halobacteria into an Emended Order Halobacteriales and Two New Orders, Haloferacales Ord. Nov. and Natrialbales Ord. Nov., Containing the Novel Families Haloferacaceae Fam. Nov. and Natrialbaceae Fam. Nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1050–1069. [Google Scholar] [CrossRef]
- Cui, H.-L.; Gao, X.; Yang, X.; Xu, X.-W. Halolamina pelagica Gen. Nov., Sp. Nov., a New Member of the Family Halobacteriaceae. Int. J. Syst. Evol. Microbiol. 2011, 61, 1617–1621. [Google Scholar] [CrossRef]
- Cui, H.-L.; Mou, Y.-Z.; Yang, X.; Zhou, Y.-G.; Liu, H.-C.; Zhou, P.-J. Halorubellus salinus Gen. Nov., Sp. Nov. and Halorubellus litoreus Sp. Nov., Novel Halophilic Archaea Isolated from a Marine Solar Saltern. Syst. Appl. Microbiol. 2012, 35, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Tazi, L.; Breakwell, D.P.; Harker, A.R.; Crandall, K.A. Life in Extreme Environments: Microbial Diversity in Great Salt Lake, Utah. Extremophiles 2014, 18, 525–535. [Google Scholar] [CrossRef]
- Kakhki, A.M.; Amoozegar, M.A.; Khaledi, E.M. Diversity of Hydrolytic Enzymes in Haloarchaeal Strains Isolated from Salt Lake. Int. J. Environ. Sci. Technol. 2011, 8, 705–714. [Google Scholar] [CrossRef]
- Pašić, L.; Bartual, S.G.; Ulrih, N.P.; Grabnar, M.; Velikonja, B.H. Diversity of Halophilic Archaea in the Crystallizers of an Adriatic Solar Saltern. FEMS Microbiol. Ecol. 2005, 54, 491–498. [Google Scholar] [CrossRef]
- Fish, S.A.; Shepherd, T.J.; McGenity, T.J.; Grant, W.D. Recovery of 16S Ribosomal RNA Gene Fragments from Ancient Halite. Nature 2002, 417, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Narasingarao, P.; Podell, S.; Ugalde, J.A.; Brochier-Armanet, C.; Emerson, J.B.; Brocks, J.J.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. De Novo Metagenomic Assembly Reveals Abundant Novel Major Lineage of Archaea in Hypersaline Microbial Communities. ISME J. 2012, 6, 81–93. [Google Scholar] [CrossRef]
- Meng, J.; Xu, J.; Qin, D.; He, Y.; Xiao, X.; Wang, F. Genetic and Functional Properties of Uncultivated MCG Archaea Assessed by Metagenome and Gene Expression Analyses. ISME J. 2014, 8, 650–659. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Gelsinger, D.R.; Ma, B.; Wierzchos, J.; Ravel, J.; Davila, A.; Casero, M.C.; DiRuggiero, J. Functional Interactions of Archaea, Bacteria and Viruses in a Hypersaline Endolithic Community: Halite Metagenome. Environ. Microbiol. 2016, 18, 2064–2077. [Google Scholar] [CrossRef]
- McGonigle, J.M.; Bernau, J.A.; Bowen, B.B.; Brazelton, W.J. Robust Archaeal and Bacterial Communities Inhabit Shallow Subsurface Sediments of the Bonneville Salt Flats. mSphere 2019, 4, e00378-19. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.K.; Chaudhary, C.; Singh, L.; Sidhu, C.; Siddhardha, B.; Prasad, S.E.; Thakur, K.G. Isolation and Taxonomic Characterization of Novel Haloarchaeal Isolates from Indian Solar Saltern: A Brief Review on Distribution of Bacteriorhodopsins and V-Type ATPases in Haloarchaea. Front. Microbiol. 2020, 11, 554927. [Google Scholar] [CrossRef] [PubMed]
- Casanueva, A.; Galada, N.; Baker, G.C.; Grant, W.D.; Heaphy, S.; Jones, B.; Yanhe, M.; Ventosa, A.; Blamey, J.; Cowan, D.A. Nanoarchaeal 16S RRNA Gene Sequences Are Widely Dispersed in Hyperthermophilic and Mesophilic Halophilic Environments. Extremophiles 2008, 12, 651–656. [Google Scholar] [CrossRef]
- Gareeb, A.; Setani, M. Assessment of alkaliphilic haloarchaeal diversity in Sua pan evaporator ponds in Botswana. Afr. J. Biotechnol. 2009, 8, 259–267. [Google Scholar]
- Ghai, R.; Pašić, L.; Fernández, A.B.; Martin-Cuadrado, A.-B.; Mizuno, C.M.; McMahon, K.D.; Papke, R.T.; Stepanauskas, R.; Rodriguez-Brito, B.; Rohwer, F.; et al. New Abundant Microbial Groups in Aquatic Hypersaline Environments. Sci. Rep. 2011, 1, 135. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, M.; Santos, F.; Moreno-Paz, M.; Parro, V.; Antón, J. Unveiling Viral–Host Interactions within the Microbial Dark Matter. Nat. Commun. 2014, 5, 4542. [Google Scholar] [CrossRef] [PubMed]
- Vavourakis, C.D.; Ghai, R.; Rodriguez-Valera, F.; Sorokin, D.Y.; Tringe, S.G.; Hugenholtz, P.; Muyzer, G. Metagenomic Insights into the Uncultured Diversity and Physiology of Microbes in Four Hypersaline Soda Lake Brines. Front. Microbiol. 2016, 7, 211. [Google Scholar] [CrossRef] [PubMed]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.-F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the Phylogeny and Coding Potential of Microbial Dark Matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef]
- Williams, T.A.; Embley, T.M. Archaeal “Dark Matter” and the Origin of Eukaryotes. Genome Biol. Evol. 2014, 6, 474–481. [Google Scholar] [CrossRef]
- Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Hug, L.A.; Brown, C.T.; Wilkins, M.J.; Frischkorn, K.R.; Tringe, S.G.; Singh, A.; Markillie, L.M.; et al. Genomic Expansion of Domain Archaea Highlights Roles for Organisms from New Phyla in Anaerobic Carbon Cycling. Curr. Biol. 2015, 25, 690–701. [Google Scholar] [CrossRef]
- Stan-Lotter, H.; Fendrihan, S. Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life 2015, 5, 1487–1496. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Maleki Zanjani, B.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of Carotenoid Production by Halorubrum Sp. TBZ126: An Extremely Halophilic Archeon from Urmia Lake. Adv. Pharm. Bull. 2014, 4, 61. [Google Scholar] [CrossRef]
- Naghoni, A.; Emtiazi, G.; Amoozegar, M.A.; Rasooli, M.; Etemadifar, Z.; Shahzadeh Fazeli, S.A.; Minegishi, H.; Ventosa, A. Natronoarchaeum persicum Sp. Nov., a Haloarchaeon Isolated from a Hypersaline Lake. Int. J. Syst. Evol. Microbiol. 2017, 67, 3339–3344. [Google Scholar] [CrossRef]
- Bachran, M.; Kluge, S.; Lopez-Fernandez, M.; Cherkouk, A. Microbial Diversity in an Arid, Naturally Saline Environment. Microb. Ecol. 2019, 78, 494–505. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Origin of Isolation | Strain Code (Acession Number) | Colony Morphology (Number of Colonies Per Morphology) | pH Range | Closest 16S rRNA (a) (Accession Number) | Similarity |
|---|---|---|---|---|---|---|
| Chott Douz (CDZ) | White salt crust | CDZ60 (MW565884) | Light red and small (1) | 6–9 | Uncultured Halorubrum sp. (FN391189) | 99% |
| CDZ56 (MW565880) | Orange and big (2) | 6–9 | Halorubrum xinjiangense (AY510707) | 99% | ||
| CDZ43 (MW565881) | Light orange and small (2) | 6–9 | Halorubrum chaoviator (AM048786) | 99% | ||
| Chott DJerid (CDW) | White halite crystal salt | CDW18.2.2 (MW565877) | Mucoid pink and big (3) | 6–8 | Haloferax mediterranei (D11107) | 99% |
| CDW18.2.3(A) (MW565876) | Red and small (4) | 6–9 | Halorubrum chaoviator (AM048786) | 99% | ||
| CDW63 (MW565886) | White and big (4) | 6–9 | Uncultured haloarchaeon (GQ374937) | 99% | ||
| CDW66 (MW565885) | Red (5) | 6–9 | Uncultured Halorubrum sp. (FN391188) | 99% | ||
| CDW12 (MW565879) | Light orange (3) | 6–9 | Haloarcula vallismortis (EF645688) | 99% | ||
| Chott DJerid (CDR) | Pink halite crystal salt | CDR49 (MW565883) | Red small (4) | 6–9 | Uncultured Haloarcula sp. (EF153424) | 99% |
| CDR50 (MW565887) | Red (4) | 5–8 | Uncultured haloarchaeon (FN391270) | 99% | ||
| CDR52 (MW565878) | Orange and small (3) | 6–8 | Halogeometricum borinquense (AF002984) | 99% | ||
| CDR16 (MW565882) | Orange and big (2) | 6–9 | Haloarcula vallismortis (EF645688) | 99% | ||
| CDR36 (MW565874) | Red and big (3) | 6–8 | Halorubrum chaoviator (AM048786) | 99% | ||
| CDR04 (MW565875) | Small red (4) | 6–8 | Halorubrum chaoviator (AM048786) | 98% |
| Origin of Samples | DGGE Bands (Accession no.) | Closest RDP II Match (Accession no.)_Genus (a) | Similarity Percentage |
|---|---|---|---|
| CDR | 5R (MW564201) | Halorubrum sp. PJ75-1 (KJ644233)_Halorubrum | 97.97% |
| 5 (MW564199) | uncultured archaeon (KT216162)_unclassified_Halobacteriaceae | 98.17% | |
| 6 (MW564202) | uncultured archaeon (KR920349) unclassified halobacteria | 99.57% | |
| CDW | 4 (MW564200) | uncultured archaeon (FJ155608)_ unclassified_Halobacteriaceae | 100% |
| 3 (MW564198) | uncultured archaeon (EU722680) unclassified_Halobacteriaceae | 100% | |
| CDZ | 1 (MW564203) | Natronomonas sp. SA2 (JF950946)_Natronomonas | 95% |
| 2 (MW564204) | Uncultured archaeon LC205717_unclassified_Halobacteriaceae | 95.95% |
| Clones ID | Closest Relatives (Accession no.) | Similarity (%) | Classification (a) | ||
|---|---|---|---|---|---|
| (Accession no. Deposited) | |||||
| Sampling Site: CDR | |||||
| Total OTUs (0.03 Cut-Off) | Phylum_Family | Genus | |||
| OTU1 | CDR48A (MW565902), CDR49A (MW565903) | Uncultured Haloquadratum (FN391222) | 98 | Euryarchaeota_Halobacteriaceae | Uncultured Haloquadratum |
| OTU2 | CDR72 (MW565890) | Uncultured haloarchaeon (GQ375023) | 94 | Euryarchaeota_Halobacteriaceae | Haloquadratum |
| OTU3 | CDR54A (MW565905), CDR67 (MW565893) | Uncultured haloarchaeon (Q375025.1) | 99 | Euryarchaeota_Halobacteriaceae | Haloquadratum |
| CDR42A(MW565897), CDR46A(MW565901), CDR44A (MW565899) | Uncultured Haloquadra (AY987831.1) | 98–99 | Euryarchaeota_Halobacteriaceae | Haloquadratum | |
| CDR109A (MW565913) | Uncultured haloarchaeon (GQ375013.1) | 99 | |||
| OTU4 | CDR43A (MW565898) | Uncultured haloarchaeon (JN714405.1) | 99 | Euryarchaeota_Halobacteriaceae | Halolamina |
| OTU5 | CDR73A (MW565909) | Halorubrum chaoviator (FN391295.1) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| CDR70 (MW565891), CDR77 (MW565888) | Uncultured haloarchaeon (FN391270.2) | 98–99 | |||
| OTU6 | CDR62 (MW567835) | Strain SFH1H061 (FN391295) | 95 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| OTU7 | CDR188 (MW565910) | Uncultured Halorubrum (AM947495.1) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| OTU8 | CDR103A (MW565911) | Uncultured Halorubrum (FN391189.1) | 98 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| CDR58A (MW565907) | Uncultured haloarchaeon (FN391244) | 99 | |||
| OTU9 | CDR55A (MW565906), CDR53A (MW565904), | Strain SFG1E101 (AM947467) | 99 | Euryarchaeota_Halobacteriaceae | Halonotius |
| CDR68 (MW565892),CDR76A (MW565919) | Uncultured haloarchaeon (GQ374941.1) | 99 | |||
| OTU10 | CDR76 (MW565918) | Uncultured haloarchaeon (FN669142.1) | 98 | Euryarchaeota_Halobacteriaceae | Halonotius |
| CDR66 (MW565894), CDR63 (MW565896), | Uncultured haloarchaeon (FN391224) | 99 | |||
| CDR82 (MW565920), CDR50A (MW567831) | Uncultured haloarchaeon (FN391226) | 98 | |||
| OTU11 | CDR112 (MW565914) | Uncultured haloarchaeon (AM947467) | 99 | Euryarchaeota_Halobacteriaceae | Halonotius |
| OTU12 | CDR64A (MW565908) | Uncultured archaeon (HQ425151.1) | 98 | Euryarchaeota_Halobacteriaceae | Haloarcula |
| OTU13 | CDR75 ((MW565917) | Uncultured Haloarcula (EF153424.1) | 99 | Euryarchaeota_Halobacteriaceae | Haloarcula |
| CDR104A (MW565912), CDR74 (MW565916) | Uncultured Halobacterium (FN391277.2) | 99 | |||
| OTU14 | CDR73 (MW565889), CDR47A (MW567832) | Uncultured Halobacterium (FN391286.1) | 94 | Euryarchaeota_Halobacteriaceae | Halorubellus |
| OTU15 | Uncultured haloarchaeon (FN391224.1) | 91 | Euryarchaeota_Halobacteriaceae | Unclasssified halobacteriaceae | |
| OTU16 | CDR51A (MW567833), CDR52A (MW567834) | Uncultured Haloquadratum (AM947450.1) | 95 | Euryarchaeota_Halobacteriaceae | Unclasssified halobacteriaceae |
| OTU17 | CDR116 (MW565915), CDR45A (MW565900), | Uncultured haloarchaeon (GQ374953) | 98–99 | Euryarchaeota_Halobacteriaceae | unclassified_Halobacteriaceae |
| OTU18 | CDR65 (MW565895) | Uncultured haloarchaeon (GQ375025) | 99 | Euryarchaeota_Halobacteriaceae | Unclasssified halobacteriaceae |
| Operational Taxonomic Units (OTUs) classification and affiliation of 16S rRNA gene clone sequences of halite-salt crystals samples based on RDP II database. | |||||
| OTU1 | CDW222 (W563901), CDW208 (MW563889), CDW218 (MW563897), CDW221 (MW563900), CDW209 (MW563890), CDW211 (MW563892), CDW198 (MW563880) | Uncultured Haloquadratum sp. (FN994951.1) | 98–100 | Euryarchaeota_Halobacteriaceae | Haloquadratum |
| OTU2 | CDW 182 (MW563870), CDW181 (MW563869) | Uncultured Haloquadra sp. (AY987831.1) | 94–96 | Euryarchaeota_Halobacteriaceae | Haloquadratum |
| OTU3 | CDW 184 (MW563871) | Uncultured haloarchaeon clone (GQ374987.1) | 96 | Euryarchaeota_Halobacteriaceae | Haloquadratum |
| OTU4 | CDW195 (MW563877), CDW190 (MW563874) | Uncultured haloarchaeon (FN391230) | 99–100 | Euryarchaeota_Halobacteriaceae | Unclassified_Haloferacaceae |
| CDW197 (MW563879) | Uncultured Halorubrum sp. (FN391188.1) | 99–100 | Euryarchaeota_Halobacteriaceae | Unclassified_Haloferacaceae | |
| OTU5 | CDW202 MW563884), CDW193 (MW563876) | Uncultured haloarchaeon clone (GQ374985.1) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| OTU6 | CDW206 (MW567775), CDW194 (MW567774) | Uncultured haloarchaeon clone (AM947463) | 91 | Euryarchaeota_Halobacteriaceae | Halonotius |
| OTU7 | CDW203 (MW563885), CDW199 (MW563881) | Uncultured haloarchaeon clone (AM947467.1) | 97–98 | Euryarchaeota_Halobacteriaceae | Halonotius |
| CDW215 (MW563895), CDW216 (MW563896) | Uncultured haloarchaeon clone (AM947465) | 99 | |||
| CDW 219 (MW563898), CDW210 (MW563891) | Uncultured haloarchaeon clone (FN391225.1) | 98–99 | |||
| CDW 212 (MW563893), CDW196 (MW563878) | Uncultured haloarchaeon clone (FN391223.1) | 99 | |||
| CDW 204 (MW563886) | Uncultured haloarchaeon clone (FN391238.1) | 99 | |||
| CDW 207 (MW563888) | Uncultured haloarchaeon clone (FN391224) | 99 | |||
| OTU8 | CDW220 (MW563899), CDW205 (MW563887) | Uncultured haloarchaeon clone (FN391270.2) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| CDW214 (MW563894), CDW200 (MW563882) | Uncultured haloarchaeon clone (FN391295.1) | 98 | |||
| OTU9 | CDW224 (MW563902) | Uncultured Halobacterium clone (FN391277.2) | 97 | Euryarchaeota_Halobacteriaceae | Haloarcula |
| CDW189 (MW563873), CDW185 (MW563872) | Uncultured archaeon (HQ425180) | 99 | |||
| OTU10 | CDW228 (MW563905) | Uncultured haloarchaeon clone (JN714408.1) | 95 | Euryarchaeota_Halobacteriaceae | Unclassified_Halobacteriaceae |
| CDW229 (MW563906) | Uncultured Halobacterium clone (FN391290.1) | 98 | |||
| OTU11 | CDW192 (MW563875), CDW226 (MW563903) | Uncultured archaeon clone (EU722673.1)| | 95–98 | Euryarchaeota_Halobacteriaceae | Unclassified_Halobacteriaceae |
| OTU12 | CDW227 (MW563904), CDW201 ( MW563883) | Uncultured Halobacterium sp. (AM947502) | 98–99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| CDW197 (MW563879) | Uncultured Halobacterium sp. (FN391188.1) | 99 | |||
| Operational Taxonomic Units (OTUs) classification and affiliation of 16S rRNA gene clone sequences of halite-salt crystals samples based on RDP II database. | |||||
| OTU1 | CDZ122 (MW564064), CDZ149 ((MW564056), CDZ141 (MW564061), CDZ127 (MW564063), CDZ138 (MW564062), CDZ143 (MW564059) | Uncultured archaeon (FJ172053) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| CDZ144 (MW564058) | Uncultured haloarchaeon (EF533955.1) | 99 | Euryarchaeota_Halobacteriaceae | ||
| CDZ154 (MW564054) | Uncultured Halobacterium (FN994971.1) | 99 | Euryarchaeota_Halobacteriaceae | ||
| OTU2 | CDZ142 (MW564060), CDZ148 (MW564057), CDZ150 (MW564055) | Uncultured archaeon (FJ172058) | 99 | Euryarchaeota_Halobacteriaceae | Halorubrum |
| Alpha Diversity | |||||
|---|---|---|---|---|---|
| Samples and Approaches | Total of Cloned Sequences/Valid Reads | Total OTU0.03 a | Shannon Index | Chao Index | Good’s Coverage |
| 16S rRNA Clones libraries approach | |||||
| CDR | 38 | 18 | 2.67 | 2.0 | 76.31 |
| CDW | 40 | 12 | 2.25 | 12.16 | 95 |
| CDZ | 11 | 0.2 | 0.58 | 25.2 | 100 |
| Amplicon analysis | |||||
| CDZ | 731 | 10 | 1.245 | 11 | 99.73 |
| CDW | 2119 | 49 | 3.255 | 49.4 | 99.9 |
| CDR | 3198 | 131 | 3.559 | 134.7 | 99.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najjari, A.; Stathopoulou, P.; Elmnasri, K.; Hasnaoui, F.; Zidi, I.; Sghaier, H.; Ouzari, H.I.; Cherif, A.; Tsiamis, G. Assessment of 16S rRNA Gene-Based Phylogenetic Diversity of Archaeal Communities in Halite-Crystal Salts Processed from Natural Saharan Saline Systems of Southern Tunisia. Biology 2021, 10, 397. https://doi.org/10.3390/biology10050397
Najjari A, Stathopoulou P, Elmnasri K, Hasnaoui F, Zidi I, Sghaier H, Ouzari HI, Cherif A, Tsiamis G. Assessment of 16S rRNA Gene-Based Phylogenetic Diversity of Archaeal Communities in Halite-Crystal Salts Processed from Natural Saharan Saline Systems of Southern Tunisia. Biology. 2021; 10(5):397. https://doi.org/10.3390/biology10050397
Chicago/Turabian StyleNajjari, Afef, Panagiota Stathopoulou, Khaled Elmnasri, Faten Hasnaoui, Ines Zidi, Haitham Sghaier, Hadda Imene Ouzari, Ameur Cherif, and George Tsiamis. 2021. "Assessment of 16S rRNA Gene-Based Phylogenetic Diversity of Archaeal Communities in Halite-Crystal Salts Processed from Natural Saharan Saline Systems of Southern Tunisia" Biology 10, no. 5: 397. https://doi.org/10.3390/biology10050397
APA StyleNajjari, A., Stathopoulou, P., Elmnasri, K., Hasnaoui, F., Zidi, I., Sghaier, H., Ouzari, H. I., Cherif, A., & Tsiamis, G. (2021). Assessment of 16S rRNA Gene-Based Phylogenetic Diversity of Archaeal Communities in Halite-Crystal Salts Processed from Natural Saharan Saline Systems of Southern Tunisia. Biology, 10(5), 397. https://doi.org/10.3390/biology10050397