
Supplementation with >Your< Iron Syrup Corrects Iron Status in a Mouse Model of Diet-Induced Iron Deficiency



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
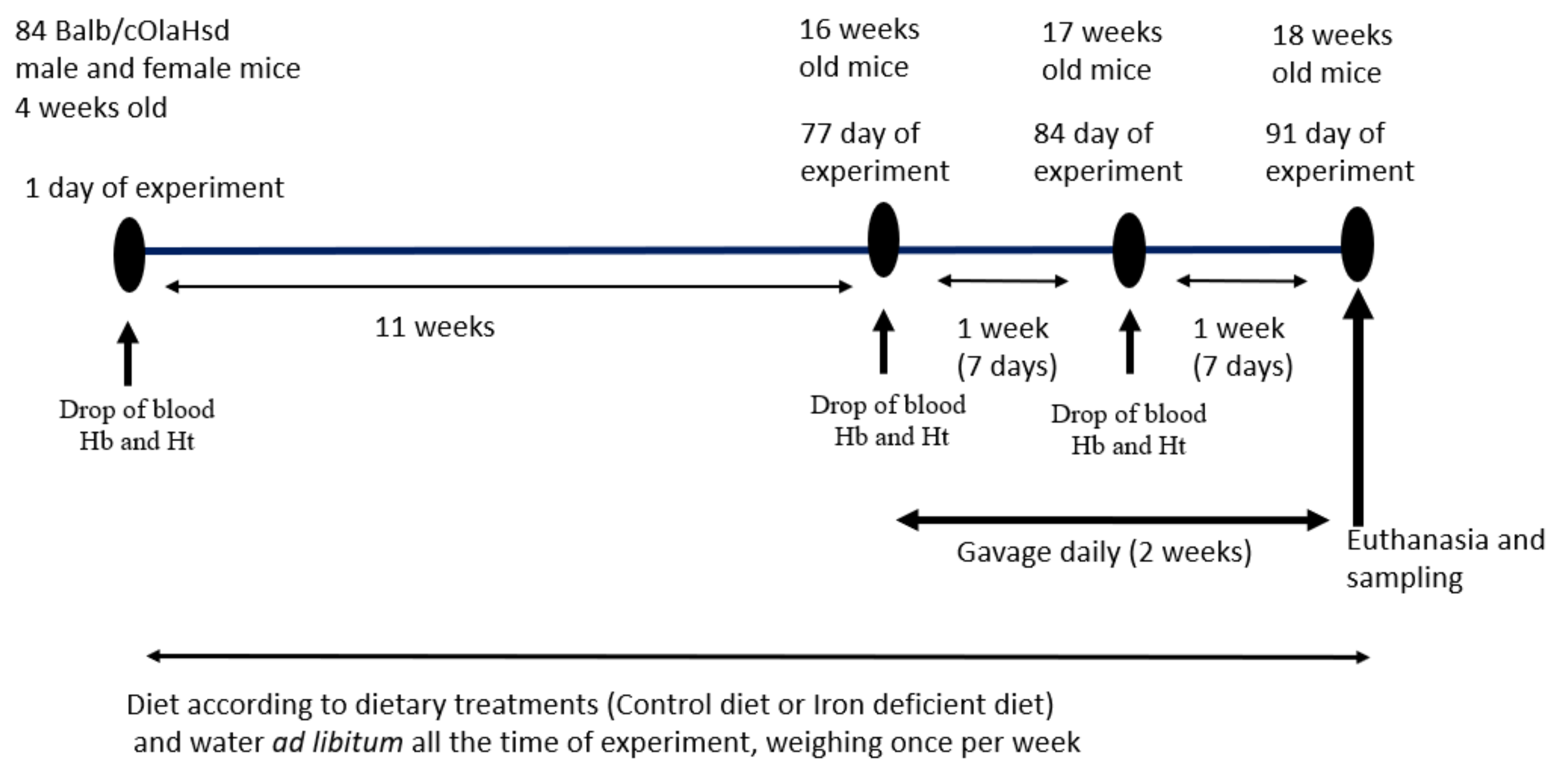
2.1. Study Design
2.2. Diets
2.3. Experimental Products
2.4. Animals and Gavage Feeding Regime
2.5. Haemoglobin and Haematocrit Determination
2.6. Iron Determination in Blood Serum and Liver
2.7. Ferritin Determination in Blood Serum and Liver
2.8. qPCR Analyses of Hamp (Hepcidin Antimicrobial Peptide) and Selected Liver Inflammatory Marker Genes
2.9. Statistical Analysis
3. Results
3.1. Chemical Composition of the Diets and of >Your< Iron Syrup
3.2. Food Intake, Growth Traits and Organ Weights
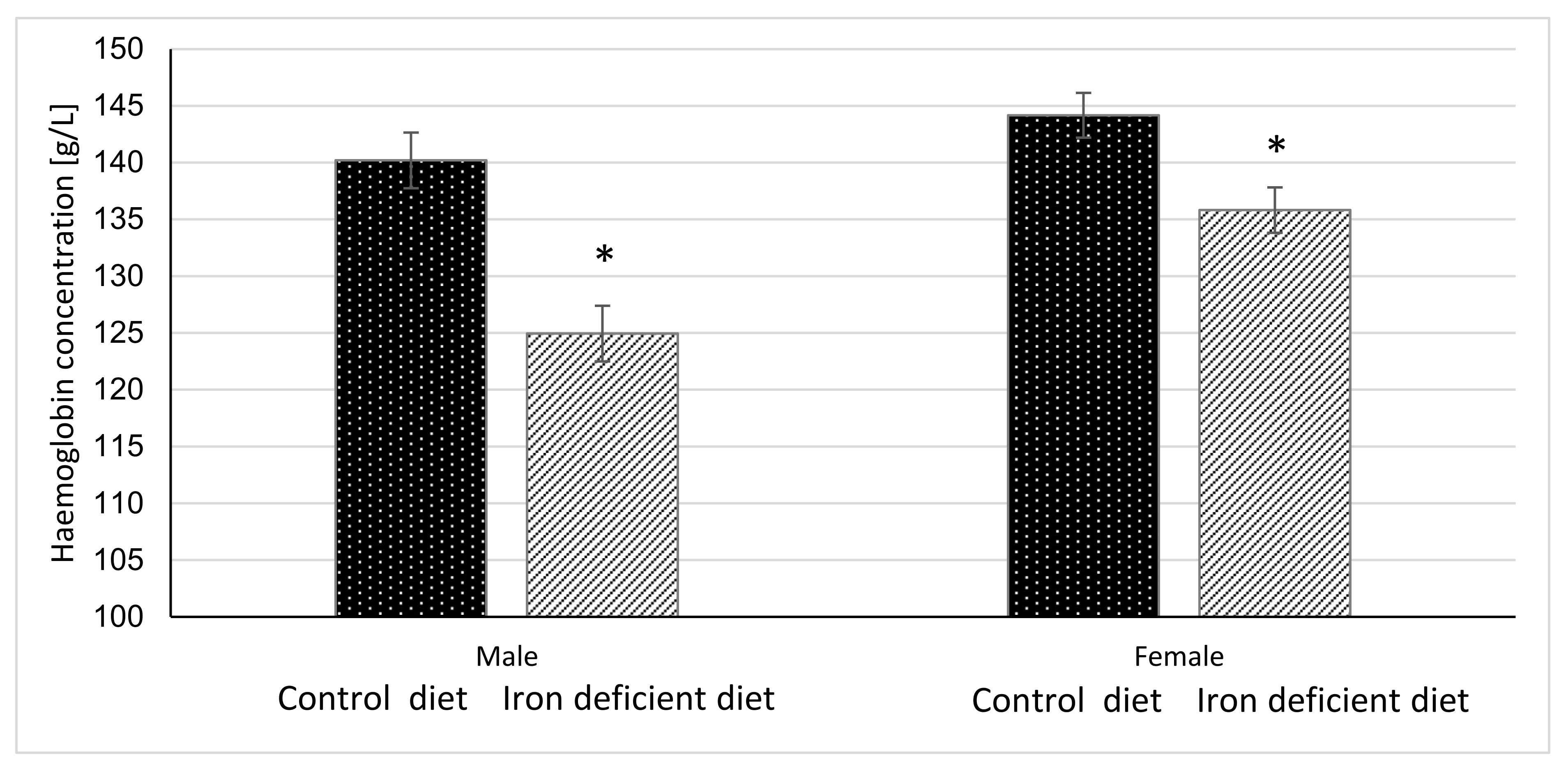
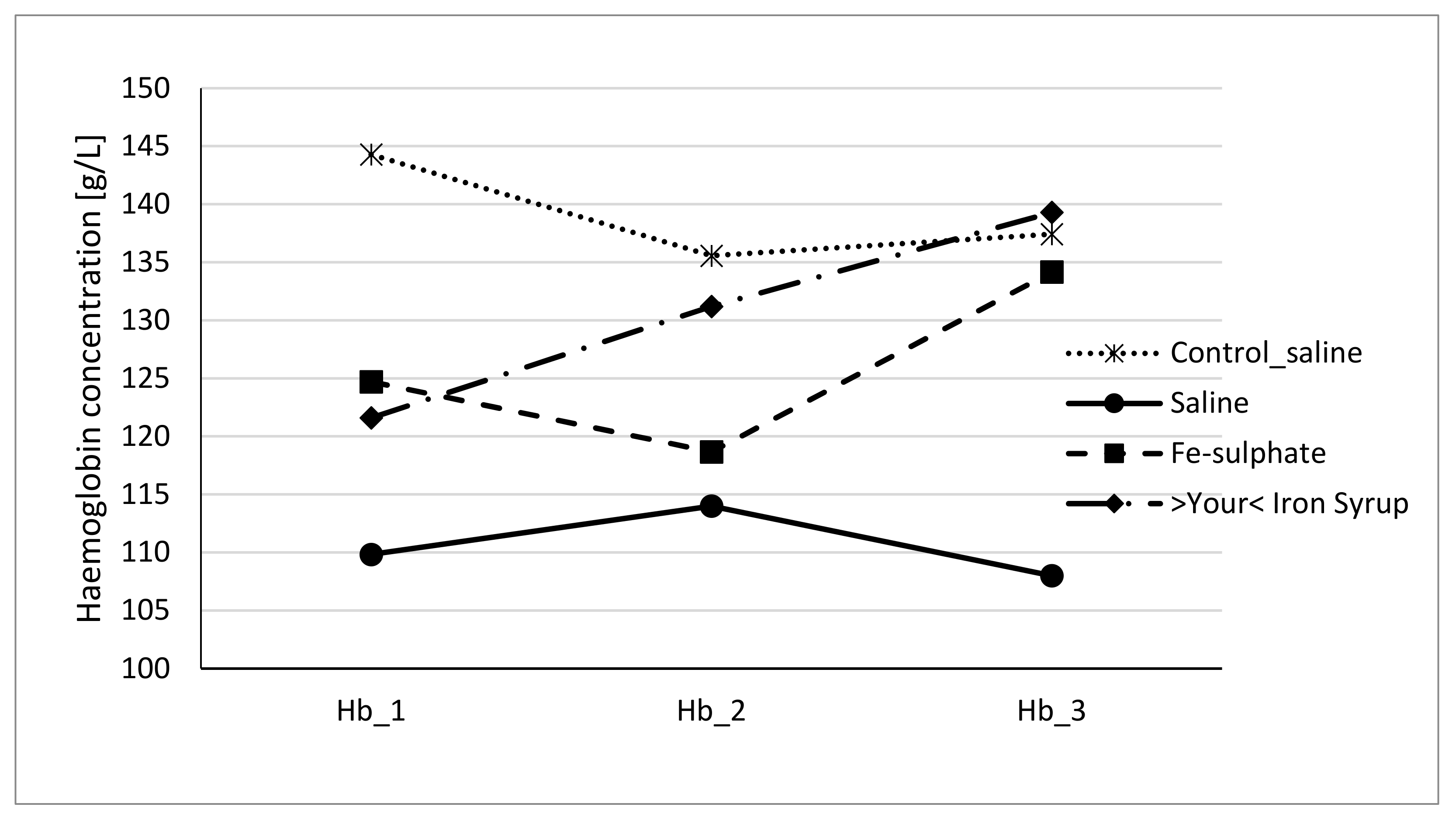
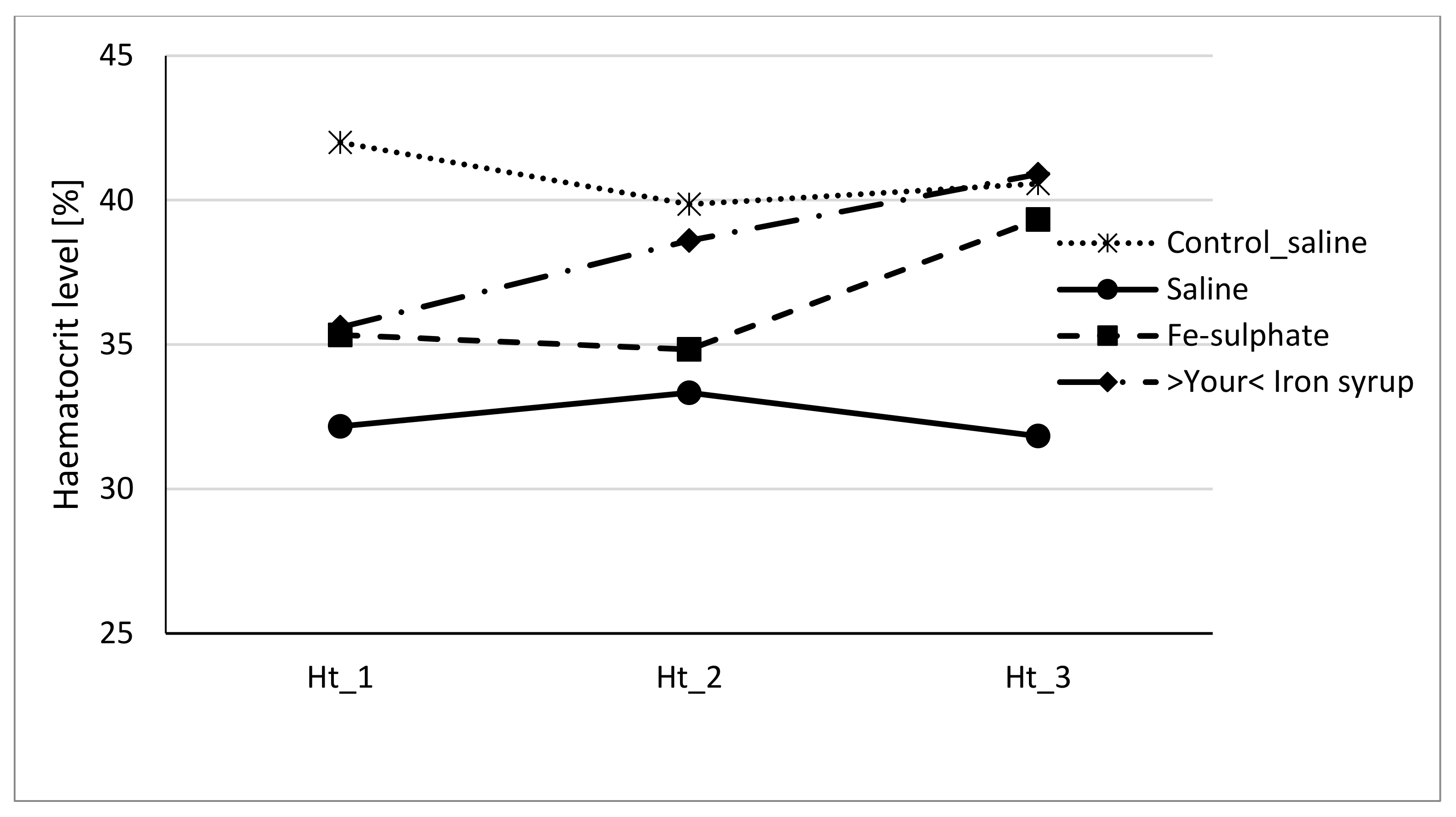
3.3. Haemoglobin and Haematocrit Level
3.4. Iron Levels in Serum and Liver
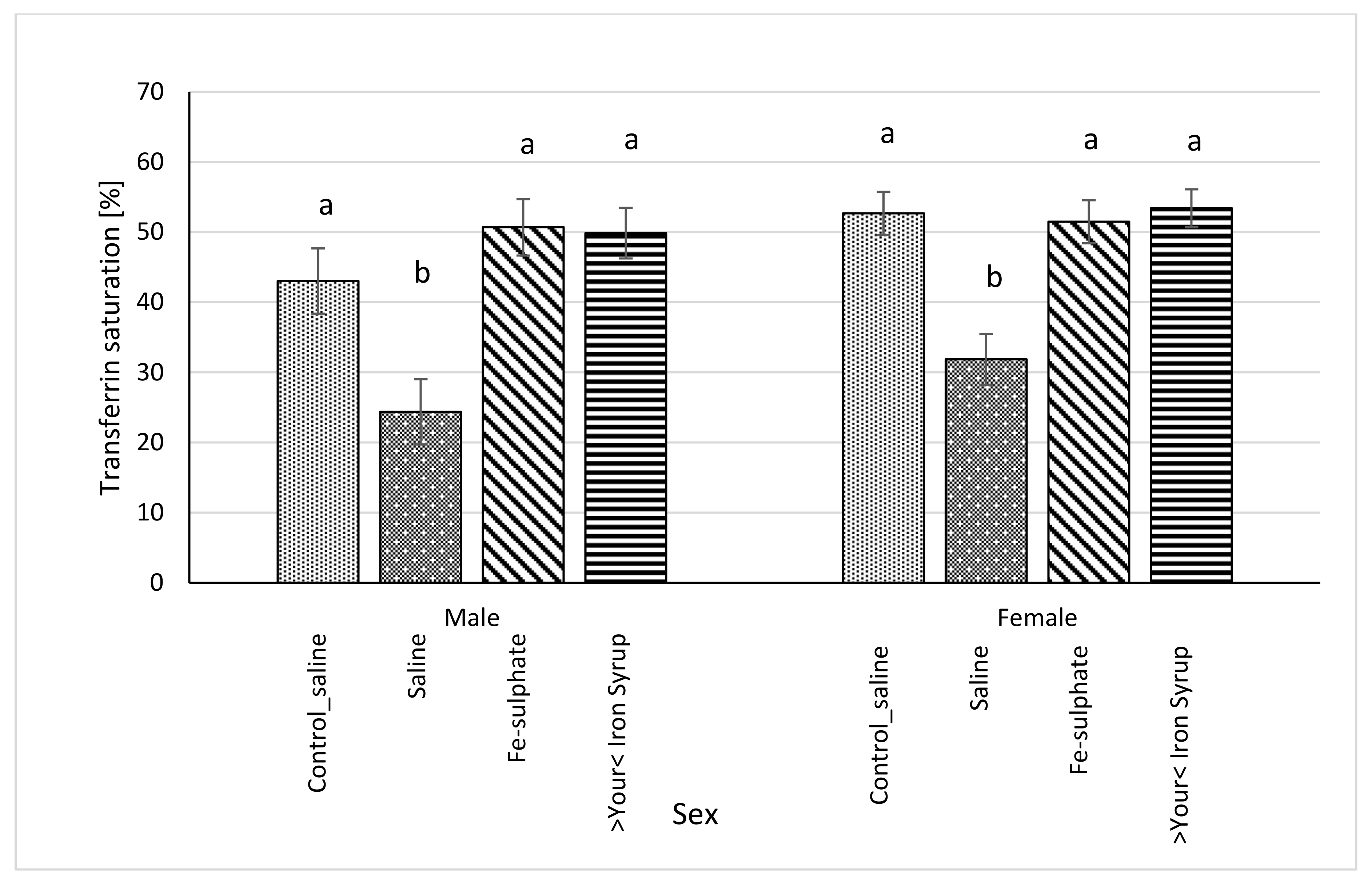
3.5. Ferritin Level in Serum and Liver
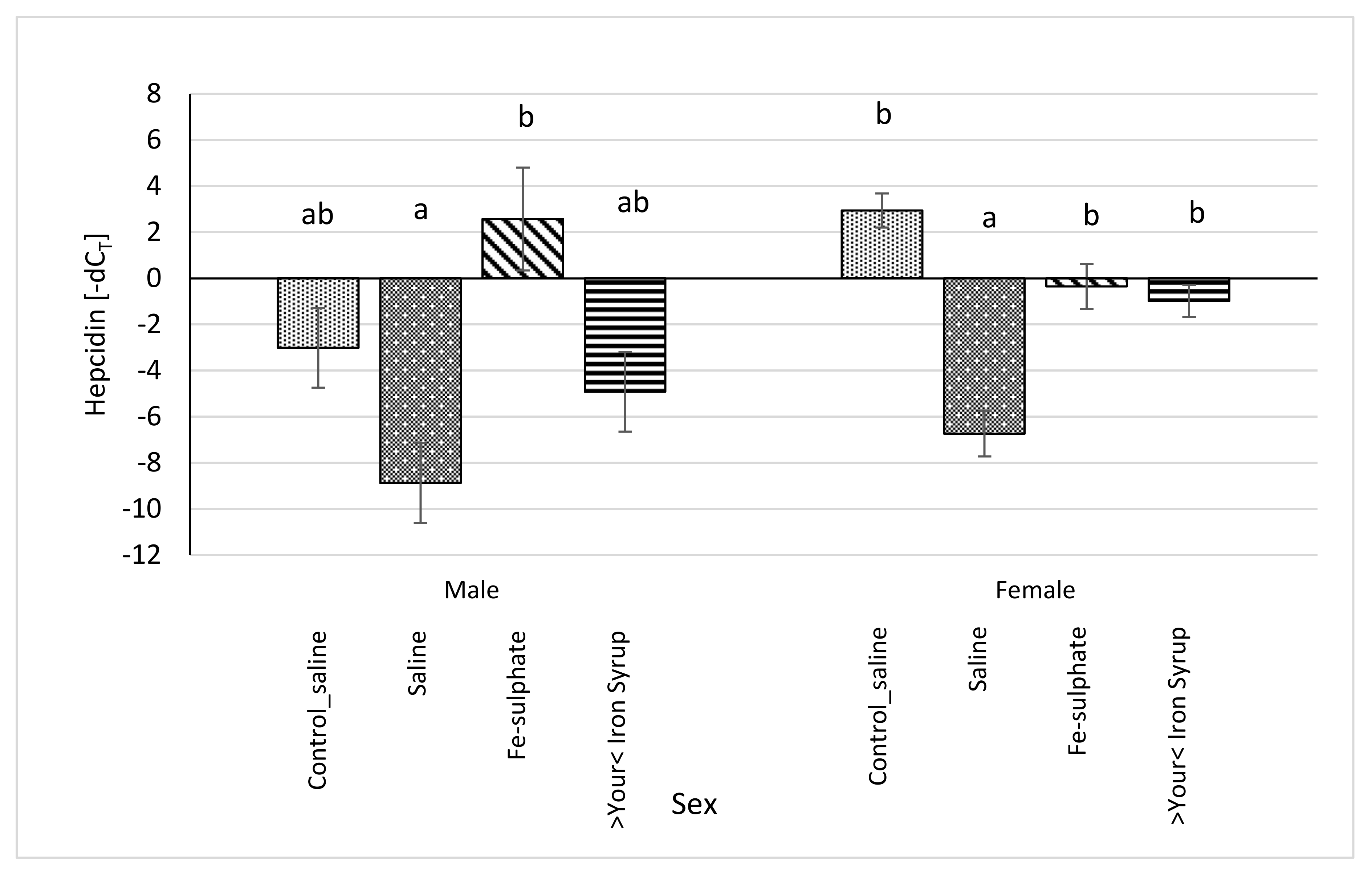
3.6. mRNA Expression of Hamp and Inflammatory Markers in the Liver
4. Discussion
4.1. Efficacy of Iron Deficient Diet in Inducing Iron Deficiency in Mice
4.2. Effects of Fe-Sulphate and >Your< Iron Syrup
4.3. Expression of Hepcidin (Hamp) and Inflammatory Marker mRNAs in the Liver
4.4. Sex Differences
4.5. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Girelli, D.; Ugolini, S.; Busti, F.; Marchi, G.; Castagna, A. Modern iron replacement therapy: Clinical and pathophysiological insights. Int. J. Hematol. 2018, 107, 16–30. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. An overview of evidence for a causal relation between iron deficiency during development and deficits in cognitive or behavioral function. Am. J. Clin. Nutr. 2007, 85, 931–945. [Google Scholar] [CrossRef]
- Georgieff, M.K. Long-term brain and behavioral consequences of early iron deficiency. Nutr. Rev. 2011, 69, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Yiannikourides, A.; Latunde-Dada, G. A Short Review of Iron Metabolism and Pathophysiology of Iron Disorders. Medicines 2019, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Pirman, T.; Orešnik, A. Fe bioavailability from Fe-enriched yeast biomass in growing rats. Animal 2012, 6, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Szudzik, M.; Starzyński, R.R.; Jończy, A.; Mazgaj, R.; Lenartowicz, M.; Lipiński, P. Iron supplementation in suckling piglets: An ostensibly easy therapy of neonatal iron deficiency anemia. Pharmaceuticals 2018, 11, 128. [Google Scholar] [CrossRef] [PubMed]
- WHO. Nutritional Anaemias: Tools for Effective Prevention and Control. World Health Organization: Geneva, Switzerland, 2017; pp. 1–83. [Google Scholar]
- Kassebaum, N.J.; Fleming, T.D.; Flaxman, A.; Phillips, D.E.; Steiner, C.; Barber, R.M.; Hanson, S.W.; Moradi-Lakeh, M.; Coffeng, L.E.; Haagsma, J.; et al. The Global Burden of Anemia. Hematol. Oncol. Clin. North Am. 2016, 30, 247–308. [Google Scholar] [CrossRef]
- Tolkien, Z.; Stecher, L.; Mander, A.P.; Pereira, D.I.A.; Powell, J.J. Ferrous sulfate supplementation causes significant gastrointestinal side-effects in adults: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0117383. [Google Scholar] [CrossRef]
- Churio, O.; Durán, E.; Guzmán-Pino, S.A.; Valenzuela, C. Use of encapsulation technology to improve the efficiency of an iron oral supplement to prevent anemia in suckling pigs. Animals 2019, 9, 1. [Google Scholar] [CrossRef]
- Wegmüller, R.; Zimmermann, M.B.; Moretti, D.; Arnold, M.; Langhans, W.; Hurrell, R.F. Particle size reduction and encapsulation affect the bioavailability of ferric pyrophosphate in rats. J. Nutr. 2004, 134, 3301–3304. [Google Scholar] [CrossRef]
- Fidler, M.C.; Walczyk, T.; Davidsson, L.; Zeder, C.; Sakaguchi, N.; Juneja, L.R.; Hurrell, R.F. A micronised, dispersible ferric pyrophosphate with high relative bioavailability in man. Br. J. Nutr. 2004, 91, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Rojo, R.; Pérez-Granados, A.M.; Toxqui, L.; González-Vizcayno, C.; Delgado, M.A.; Vaquero, M.P. Efficacy of a microencapsulated iron pyrophosphate-fortified fruit juice: A randomised, double-blind, placebo-controlled study in Spanish iron-deficient women. Br. J. Nutr. 2011, 105, 1652–1659. [Google Scholar] [CrossRef] [PubMed]
- Navas-Carretero, S.; Pérez-Granados, A.M.; Sarriá, B.; Vaquero, M.P. Iron absorption from meat pate fortified with ferric pyrophosphate in iron-deficient women. Nutrition 2009, 25, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D.; Zimmermann, M.B.; Muthayya, S.; Thankachan, P.; Lee, T.C.; Kurpad, A.V.; Hurrell, R.F. Extruded rice fortified with micronized ground ferric pyrophosphate reduces iron deficiency in Indian schoolchildren: A double-blind randomized controlled trial1-3. Am. J. Clin. Nutr. 2006, 84, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Roe, M.A.; Collings, R.; Hoogewerff, J.; Fairweather-Tait, S.J. Relative bioavailability of micronized, dispersible ferric pyrophosphate added to an apple juice drink. Eur. J. Nutr. 2009, 48, 115–119. [Google Scholar] [CrossRef]
- Miller, D. Iron fortification of the food supply: A balancing act between bioavailability and iron-catalyzed oxidation reactions. In Nutritional Biotechnology in the Feed and Food Industries: Proceedings of Alltech’s 18th Annual Symposium: From Niche Markets to Mainstream, Lexington, KY, USA, 13–15 May 2002; Lyons, T.P., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 2002; pp. 239–244. [Google Scholar]
- Stoltzfus, R.J.; Dreyfuss, M.L. Guidelines for the Use of Iron Supplements to Prevent and Treat Iron Deficiency Anaemia; International Nutritional Anaemia Group: Washington, DC, USA, 1998; ISBN 1578810205. [Google Scholar]
- Zimmermann, M.B. The potential of encapsulated iron compounds in food fortification: A review. Int. J. Vitam. Nutr. Res. 2004, 74, 453–461. [Google Scholar] [CrossRef]
- McDowell, L.R. Minerals in Animal and Human Nutrition, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Asperti, M.; Gryzik, M.; Brilli, E.; Castagna, A.; Corbella, M.; Gottardo, R.; Girelli, D.; Tarantino, G.; Arosio, P.; Poli, M. Sucrosomial® iron supplementation in mice: Effects on blood parameters, hepcidin, and inflammation. Nutrients 2018, 10, 1349. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 21st ed.; Horwith, W., Ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Kappel, S.; Hawkins, P.; Mendl, M.T. To group or not to group? Good practice for housing male laboratory mice. Animals 2017, 7, 88. [Google Scholar] [CrossRef]
- Zidar, J.; Weber, E.M.; Ewaldsson, B.; Tjäder, S.; Lilja, J.; Mount, J.; Svensson, C.; Svensk, E.; Udén, E.; Törnqvist, E. Group and single housing of male mice: Collected experiences from research facilities in sweden. Animals 2019, 9, 1010. [Google Scholar] [CrossRef]
- Hurst, J.L.; West, R.S. Taming anxiety in laboratory mice. Nat. Methods 2010, 7, 825–826. [Google Scholar] [CrossRef]
- Meikle, M.N.; Arévalo, A.P.; Schlapp, G.; Fernández-Graña, G.; Menchaca, A.; Crispo, M. Long-Term Effect of Environmental Enrichment on Reproductive Performance of Swiss Webster Mice and Their Female Offspring. Animals 2020, 10, 1438. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, Scientific Research Publishing: Vienna, Austria, 2018.
- Lou, D.Q.; Nicolas, G.; Lesbordes, J.C.; Viatte, L.; Grimber, G.; Szajnert, M.F.; Kahn, A.; Vaulont, S. Functional differences between hepcidin 1 and 2 in transgenic mice. Blood 2004, 103, 2816–2821. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [PubMed]
- Giulietti, A.; Overbergh, L.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. An overview of real-time quantitative PCR: Applications to quantify cytokine gene expression. Methods 2001, 25, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Langford, D.J.; Bailey, A.L.; Chanda, M.L.; Clarke, S.E.; Drummond, T.E.; Echols, S.; Glick, S.; Ingrao, J.; Klassen-Ross, T.; Lacroix-Fralish, M.L.; et al. Coding of facial expressions of pain in the laboratory mouse. Nat. Methods 2010, 7, 447–449. [Google Scholar] [CrossRef]
- Hohlbaum, K.; Corte, G.M.; Humpenöder, M.; Merle, R.; Thöne-Reineke, C. Reliability of the Mouse Grimace Scale in C57BL/6JRj Mice. Animals 2020, 10, 1648. [Google Scholar] [CrossRef]
- Liu, Z.; Mohan, S.; Yakar, S. Does the GH/IGF-1 axis contribute to skeletal sexual dimorphism? Evidence from mouse studies. Growth Horm. IGF Res. 2016, 27, 7–17. [Google Scholar] [CrossRef]
- Chen, X.; McClusky, R.; Chen, J.; Beaven, S.W.; Tontonoz, P.; Arnold, A.P.; Reue, K. The number of X chromosomes causes sex differences in adiposity in mice. PLoS Genet. 2012, 8, e1002709. [Google Scholar] [CrossRef]
- Clothier, B.; Robinson, S.; Akhtar, R.A.; Francis, J.E.; Peters, T.J.; Raja, K.; Smith, A.G. Genetic variation of basal iron status, ferritin and iron regulatory protein in mice: Potential for modulation of oxidative stress. Biochem. Pharmacol. 2000, 59, 115–122. [Google Scholar] [CrossRef]
- Bechensteen, A.G.; Halvorsen, S. Parenteral iron increases serum erythropoietin concentration during the “early anaemia’’ of 10-20-day-old mice. Br. J. Haematol. 1996, 94, 529–532. [Google Scholar] [CrossRef]
- Chaudhury, C.; Kim, J.; Mehnaz, S.; Wani, M.A.; Oberyszyn, T.M.; Bronson, C.L.; Mohanty, S.; Hayton, W.L.; Robinson, J.M.; Anderson, C.L. Accelerated transferrin degradation in HFE-deficient mice is associated with increased transferrin saturation. J. Nutr. 2006, 136, 2993–2998. [Google Scholar] [CrossRef][Green Version]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef]
- Punnonen, K.; Irjala, K.; Rajamäki, A. Serum transferrin receptor and its ratio to serum ferritin in the diagnosis of iron deficiency. Blood 1997, 89, 1052–1057. [Google Scholar] [CrossRef]
- Porter, J.B.; El-Alfy, M.; Viprakasit, V.; Giraudier, S.; Chan, L.L.; Lai, Y.; El-Ali, A.; Han, J.; Cappellini, M.D. Utility of labile plasma iron and transferrin saturation in addition to serum ferritin as iron overload markers in different underlying anemias before and after deferasirox treatment. Eur. J. Haematol. 2016, 96, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Ambrosy, A.P.; Fitzpatrick, J.K.; Tabada, G.H.; Gurwitz, J.H.; Artz, A.; Schrier, S.L.; Rao, S.V.; Reynolds, K.; Smith, D.H.; Peterson, P.N.; et al. A reduced transferrin saturation is independently associated with excess morbidity and mortality in older adults with heart failure and incident anemia. Int. J. Cardiol. 2020, 309, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Babitt, J.L. Liver iron sensing and body iron homeostasis. Blood 2019, 133, 18–29. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Porteu, A.; Mativet, S.; Beaumont, C.; Grandchamp, B.; Sirito, M.; Sawadogo, M.; Kahn, A.; Vaulont, S. Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc. Natl. Acad. Sci. USA 2002, 99, 4596–4601. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loréal, O. A New Mouse Liver-specific Gene, Encoding a Protein Homologous to Human Antimicrobial Peptide Hepcidin, Is Overexpressed during Iron Overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef]
- Nicolas, G.; Chauvet, C.; Viatte, L.; Danan, J.L.; Bigard, X.; Devaux, I.; Beaumont, C.; Kahn, A.; Vaulont, S. The gene encoding the iron regulatory peptide hepcidin is regulated by anemia, hypoxia, and inflammation. J. Clin. Investig. 2002, 110, 1037–1044. [Google Scholar] [CrossRef]
- Hahn, P.; Song, Y.; Ying, G.S.; He, X.; Beard, J.; Dunaief, J.L. Age-dependent and gender-specific changes in mouse tissue iron by strain. Exp. Gerontol. 2009, 44, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Arnold, A.P.; Reue, K. A Guide for the Design of Pre-clinical Studies on Sex Differences in Metabolism. Cell Metab. 2017, 25, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Clayton, J.A.; Collins, F.S. NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef]
- Quintero-Gutiérrez, A.G.; González-Rosendo, G.; Sánchez-Muñoz, J.; Polo-Pozo, J.; Rodríguez-Jerez, J.J. Bioavailability of heme iron in biscuit filling using piglets as an animal model for humans. Int. J. Biol. Sci. 2008, 4, 58–62. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet (D11112201) | Iron Deficient Diet(D15071503) | |
|---|---|---|
| Casein | 200 | 200 |
| L-Cystine | 3 | 3 |
| Corn Starch | 381 | 381 |
| Maltodextrin 10 | 110 | 110 |
| Dextrose | 150 | 150 |
| Cellulose, BW200 | 75 | 75 |
| Inulin | 25 | 25 |
| Soybean Oil | 70 | 70 |
| Mineral Mix S10026 1 | 10 | |
| Mineral Mix S18708 (no added iron) 2 | 10 | |
| Dicalcium Phosphate | 13 | |
| Dicalcium Phosphate, Chemically Pure | 13 | |
| Calcium Carbonate | 5.5 | |
| Calcium Carbonate, Reagent Grade | 5.5 | |
| Potassium Citrate, 1 H2O | 16.5 | 16.5 |
| Vitamin Mix V10001 3 | 10 | 10 |
| Choline Bitartrate | 2 | 2 |
| Yellow Dye #5, FD&C | 0.025 | |
| Blue Dye #1, FD&C | 0.025 | 0.05 |
| Total | 1071.05 | 1071.05 |
| Iron Deficient Diet Groups | Control Diet Groups | ||
|---|---|---|---|
| Saline | Iron deficient diet12th and 13th week each day 150 µL 0.9% saline solution | Saline | Control diet12th and 13th week each day 150 µL 0.9% saline solution |
| Fe-sulphate | Iron deficient diet12th and 13th week each day 1 mg iron (from Ferrous sulphate)/kg body mass dissolved in 150 µL 0.9% saline solution | Fe-sulphate | Control diet12th and 13th week each day 1 mg iron (from Ferrous sulphate)/kg body mass dissolved in 150 µL 0.9% saline solution |
| >Your< Iron Syrup | Iron deficient diet12th and 13th week each day 1 mg iron (from >Your< Iron Syrup)/kg body mass—9 µL of >Your< Iron Syrup was mixed with 141 µL 0.9% saline solution | >Your< Iron Syrup | Control diet12th and 13th week each day 1 mg iron (from >Your< Iron Syrup)/kg body mass- 9 µL of Your Iron Syrup was mixed with 141 µL 0.9% saline solution |
| Target Gene | Primer Sequence | Amplicon Size (bp) | Primer Source |
|---|---|---|---|
| Actb | F: 5′-GTGACGTTGACATCCGTAAAGA-3′ | 245 | PrimerBank [29] ID = 145966868c1 |
| R: 5′-GCCGGACTCATCGTACTCC -3′ | |||
| Gapdh | F: 5′-TCACCACCATGGAGAAGGC-3′ | 168 | Giulietti et al. [30] |
| R: 5′-GCTAAGCAGTTGGTGGTGCA-3′ | |||
| Hamp | F: 5′-AAGCAGGGCAGACATTGCGAT-3′ | 141 | Asperti et al. [21] |
| R: 5′-CAGGATGTGGCTCTAGGCTATGT-3′ | |||
| Crp | F: 5′-GCTACTCTGGTGCCTTCTGATCA-3′ | 135 | Asperti et al. [21] |
| R: 5′-GGCTTCTTTGACTCTGCTTCCA-3′ | |||
| Il6 | F: 5′-CTCTGCAAGAGACTTCCATCCAGT-3′ | 58 | Asperti et al. [21] |
| R: 5′-CGTGGTTGTCACCAGCATCA-3′ | |||
| Saa1 | F: 5′-AGAGGACATGAGGACACCAT-3′ | 85 | Asperti et al. [21] |
| R: 5′-CAGGAGGTCTGTAGTAATTGG-3′ | |||
| Socs3 | F: 5′-TTAAATGCCCTCTGTCCCAGG-3′ | 51 | Asperti et al. [21] |
| R: 5′-TGTTTGGCTCCTTGTGTGCC-3′ |
| Control Diet (D11112201) | Iron Deficient Diet (D15071503) | |
|---|---|---|
| Dry matter (g/kg) | 914.8 | 919.3 |
| Crude fibre (g) | 71.7 | 72.3 |
| Crude fats (g) | 69.5 | 69.4 |
| Crude protein (g) | 171.6 | 177.7 |
| Crude ash (g) | 34.8 | 35.5 |
| Non-nitrogen extractives (g) | 645.1 | 652.3 |
| Minerals | ||
| Na (g) | 1.17 | 1.16 |
| K (g) | 6.92 | 6.31 |
| Ca (g) | 6.23 | 5.96 |
| Mg (g) | 0.60 | 0.61 |
| P (g) | 4.49 | 4.35 |
| Fe (mg) | 51.42 | 14.76 |
| Zn (mg) | 80.29 | 72.48 |
| Cu (mg) | 7.53 | 7.10 |
| Mn (mg) | 58.36 | 58.25 |
| Fe in Serum (µMol/L) | Fe in Liver (µg/g Liver) | |||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Control_saline | 31.93 ± 2.74 ab | 39.89 ± 2.99 ab | 124.34 ± 7.01 a | 158.37 ± 9.64 a |
| Iron deficient diet groups | ||||
| Saline | 21.03 ± 2.74 b | 32.41 ± 2.99 b | 55.54 ± 7.01 c | 68.96 ± 9.64 b |
| Fe-sulphate | 37.85 ± 2.56 a | 42.24 ± 2.99 ab | 89.80 ± 6.56 b | 139.06 ± 9.64 a |
| Your Iron Syrup | 39.27 ± 2.29 a | 44.40 ± 2.50 a | 63.74 ± 5.86 bc | 81.42 ± 8.06 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirman, T.; Lenardič, A.; Nemec Svete, A.; Horvat, S. Supplementation with >Your< Iron Syrup Corrects Iron Status in a Mouse Model of Diet-Induced Iron Deficiency. Biology 2021, 10, 357. https://doi.org/10.3390/biology10050357
Pirman T, Lenardič A, Nemec Svete A, Horvat S. Supplementation with >Your< Iron Syrup Corrects Iron Status in a Mouse Model of Diet-Induced Iron Deficiency. Biology. 2021; 10(5):357. https://doi.org/10.3390/biology10050357
Chicago/Turabian StylePirman, Tatjana, Ajda Lenardič, Alenka Nemec Svete, and Simon Horvat. 2021. "Supplementation with >Your< Iron Syrup Corrects Iron Status in a Mouse Model of Diet-Induced Iron Deficiency" Biology 10, no. 5: 357. https://doi.org/10.3390/biology10050357
APA StylePirman, T., Lenardič, A., Nemec Svete, A., & Horvat, S. (2021). Supplementation with >Your< Iron Syrup Corrects Iron Status in a Mouse Model of Diet-Induced Iron Deficiency. Biology, 10(5), 357. https://doi.org/10.3390/biology10050357