
Synthetic Scaffold Systems for Increasing the Efficiency of Metabolic Pathways in Microorganisms



Abstract
Simple Summary
Abstract
1. Introduction
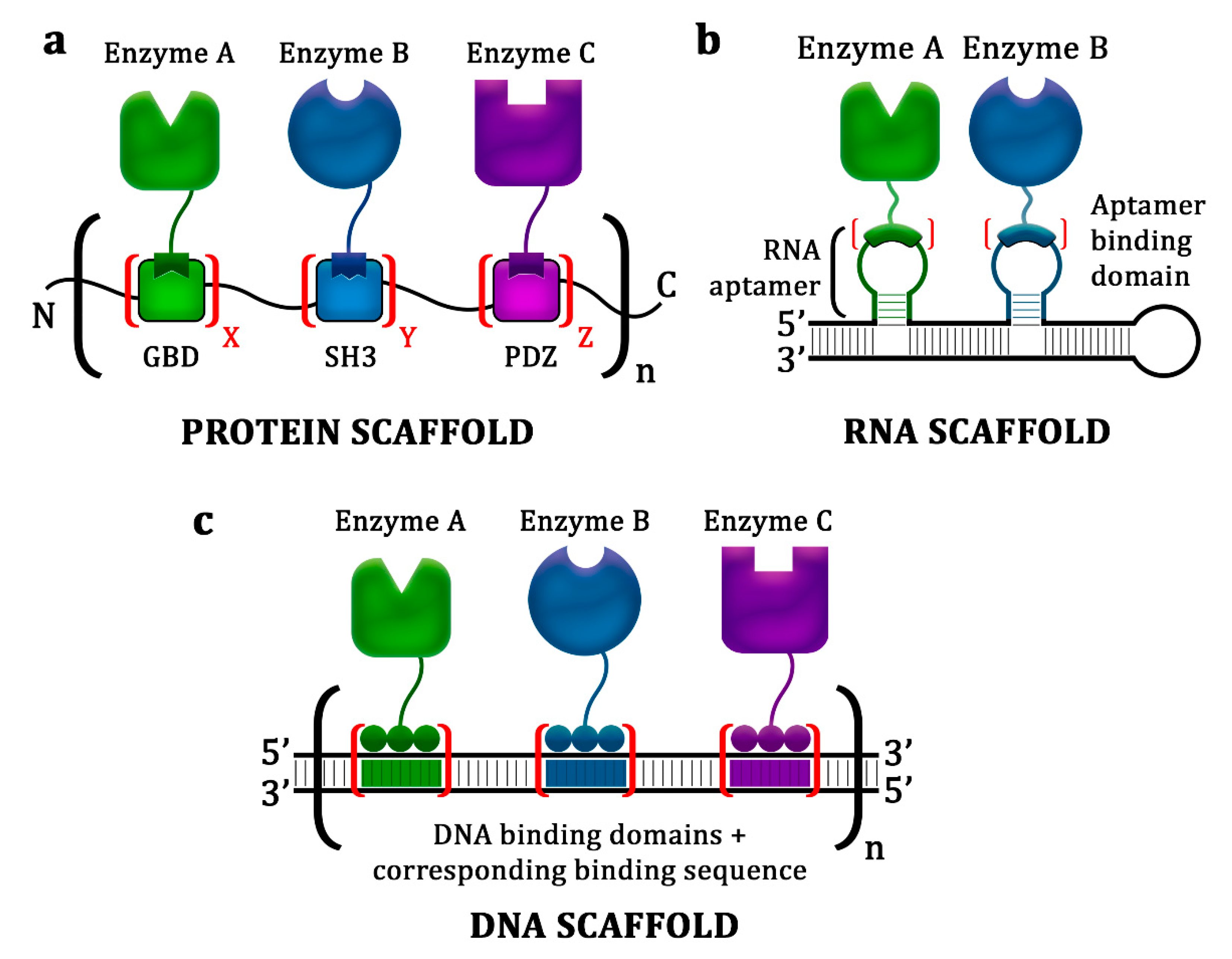
2. Protein Scaffolds
3. Nucleic Acid Scaffolds
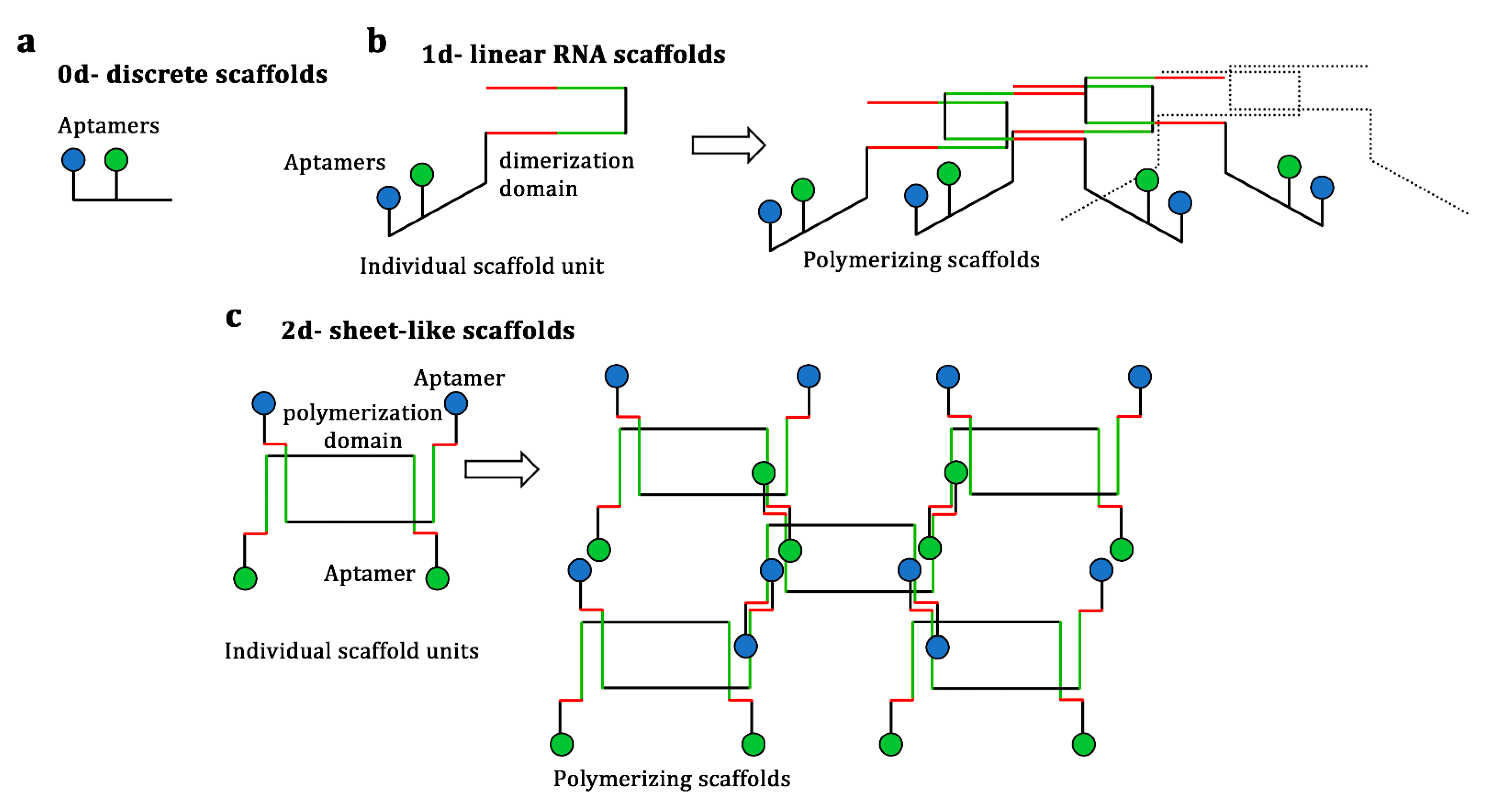
3.1. RNA Scaffolds
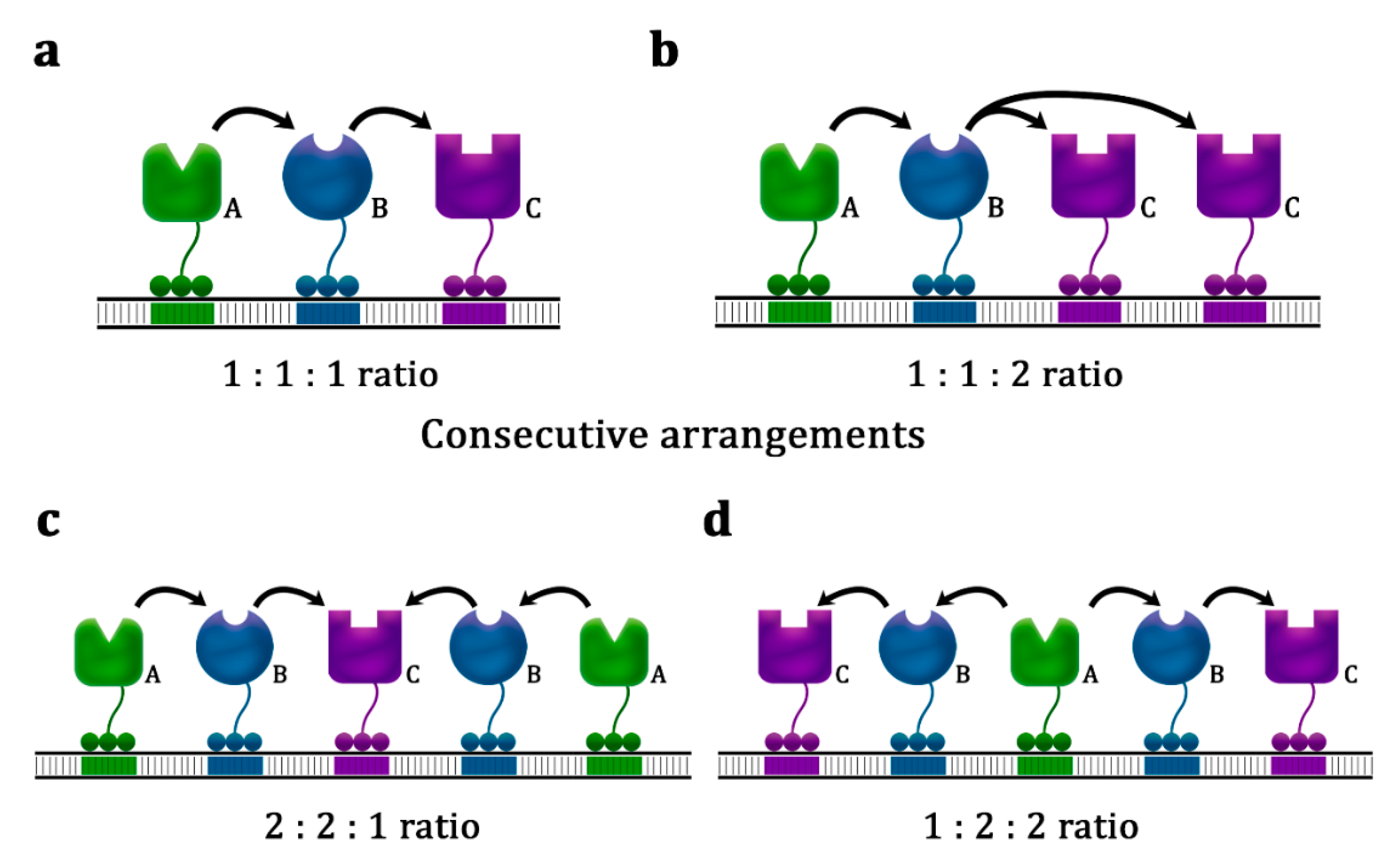
3.2. DNA Scaffolds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Santos, C.N.S.; Xiao, W.; Stephanopoulos, G. Rational, combinatorial, and genomic approaches for engineering L-tyrosine production in Escherichia coli. Proc. Natl. Acad. Sci. USA 2012, 109, 13538–13543. [Google Scholar] [CrossRef]
- Yadav, D.; Tanveer, A.; Malviya, N.; Yadav, S. Overview and Principles of Bioengineering; Elsevier Inc.: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Keasling, J.D. Manufacturing molecules through metabolic engineering. Science 2010, 330, 1355–1358. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Toparlak, T.D.; Koffas, M.A.G. Metabolic pathway balancing and its role in the production of biofuels and chemicals. Curr. Opin. Biotechnol. 2015, 33, 52–59. [Google Scholar] [CrossRef]
- Harder, B.J.; Bettenbrock, K.; Klamt, S. Model-based metabolic engineering enables high yield itaconic acid production by Escherichia coli. Metab. Eng. 2016, 38, 29–37. [Google Scholar] [CrossRef]
- Jensen, M.K.; Keasling, J.D. Recent applications of synthetic biology tools for yeast metabolic engineering. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Kim, E.; Moore, B.S.; Yoon, Y.J. Reinvigorating natural product combinatorial biosynthesis with synthetic biology. Nat. Chem. Biol. 2015, 11, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.H.; Chen, R.P.; Sun, Q.; Chen, L.; Tsai, S.L.; Chen, W. Synthetic scaffolds for pathway enhancement. Curr. Opin. Biotechnol. 2015, 36, 98–106. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, S.C.; Bui, L.M.; Kang, K.H.; Song, J.J.; Kim, S.C. Improved production of l-threonine in Escherichia coli by use of a DNA scaffold system. Appl. Environ. Microbiol. 2013, 79, 774–782. [Google Scholar] [CrossRef]
- Chen, A.H.; Silver, P.A. Designing biological compartmentalization. Trends Cell Biol. 2012, 22, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.L.; Zhu, J.; Wheeldon, I. Synthetic protein scaffolds for biosynthetic pathway colocalization on lipid droplet membranes. ACS Synth. Biol. 2017, 6, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Benčina, M.; Mori, J.; Gaber, R.; Jerala, R. Metabolic channeling Using DNA as a Scaffold. In Synthetic Biology; Lee, S.Y., Nielsen, J., Stephanopoulos, G., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 237–259. [Google Scholar]
- Pröschel, M.; Detsch, R.; Boccaccini, A.R.; Sonnewald, U. Engineering of metabolic pathways by artificial enzyme channels. Front. Bioeng. Biotechnol. 2015, 3, 1–13. [Google Scholar] [CrossRef]
- Aon, M.A.; Cortassa, S. Function of metabolic and organelle networks in crowded and organized media. Front. Physiol. 2015, 5, 523. [Google Scholar] [CrossRef] [PubMed]
- Polka, J.K.; Hays, S.G.; Silver, P.A. Building Spatial synthetic biology with compartments, scaffolds, and communities. Cold Spring Harb. Perspect. Biol. 2016, 8, a024018. [Google Scholar] [CrossRef]
- Fang, Y.; Huang, F.; Faulkner, M.; Jiang, Q.; Dykes, G.F.; Yang, M.; Liu, L.-N. Engineering and Modulating functional cyanobacterial CO2-fixing organelles. Front. Plant Sci. 2018, 9, 739. [Google Scholar] [CrossRef]
- Cameron, J.C.; Wilson, S.C.; Bernstein, S.L.; Kerfeld, C.A. Biogenesis of a bacterial organelle: The carboxysome assembly pathway. Cell 2013, 155, 1131–1140. [Google Scholar] [CrossRef]
- Jakobson, C.M.; Tullman-Ercek, D.; Mangan, N.M. Spatially organizing biochemistry: Choosing a strategy to translate synthetic biology to the factory. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Greening, C.; Lithgow, T. Formation and function of bacterial organelles. Nat. Rev. Microbiol. 2020, 18, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Agapakis, C.M.; Boyle, P.M.; Silver, P.A. Natural strategies for the spatial optimization of metabolism in synthetic biology. Nat. Chem. Biol. 2012, 8, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Na, D.; Park, J.M.; Lee, J.; Choi, S.; Lee, S.Y. Systems Metabolic engineering of microorganisms for natural and non-natural chemicals. Nat. Chem. Biol. 2012, 8, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Conrado, R.J.; Varner, J.D.; DeLisa, M.P. Engineering the spatial organization of metabolic enzymes: Mimicking nature’s synergy. Curr. Opin. Biotechnol. 2008, 19, 492–499. [Google Scholar] [CrossRef]
- Quin, M.B.; Wallin, K.K.; Zhang, G.; Schmidt-Dannert, C. Spatial organization of multi-enzyme biocatalytic cascades. Org. Biomol. Chem. 2017, 15, 4260–4271. [Google Scholar] [CrossRef]
- Qiu, X.Y.; Xie, S.S.; Min, L.; Wu, X.M.; Zhu, L.Y.; Zhu, L. Spatial Organization of enzymes to enhance synthetic pathways in microbial chassis: A systematic review. Microb. Cell Fact. 2018, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.; Sung, B.H.; Yi, J.Y.; Bui, L.M.; Lee, J.H.; Kim, S.C. Enhanced production of n-alkanes in Escherichia coli by spatial organization of biosynthetic pathway enzymes. J. Biotechnol. 2014, 192, 187–191. [Google Scholar] [CrossRef]
- Wang, C.; Yoon, S.H.; Jang, H.J.; Chung, Y.R.; Kim, J.Y.; Choi, E.S.; Kim, S.W. Metabolic engineering of Escherichia coli for alpha-farnesene production. Metab. Eng. 2011, 13, 648–655. [Google Scholar] [CrossRef]
- Fujiwara, R.; Noda, S.; Tanaka, T.; Kondo, A. Muconic acid production using gene-level fusion proteins in Escherichia coli. ACS Synth. Biol. 2018, 7, 2698–2705. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.Z.; Li, J.; Pan, X.; Cahoon, R.E.; Jaworski, J.G.; Wang, X.; Jez, J.M.; Chen, F.; Yu, O. Using Unnatural protein fusions to engineer resveratrol biosynthesis in yeast and mammalian cells. J. Am. Chem. Soc. 2006, 128, 13030–13031. [Google Scholar] [CrossRef]
- Chen, R.; Chen, Q.; Kim, H.; Siu, K.H.; Sun, Q.; Tsai, S.L.; Chen, W. Biomolecular Scaffolds for enhanced signaling and catalytic efficiency. Curr. Opin. Biotechnol. 2014, 28, 59–68. [Google Scholar] [CrossRef]
- Liu, F.; Banta, S.; Chen, W. Functional assembly of a multi-enzyme methanol oxidation cascade on a surface-displayed trifunctional scaffold for enhanced NADH production. Chem. Commun. 2013, 49, 3766–3768. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hahn, J.S. Synthetic scaffold based on a cohesin-dockerin interaction for improved production of 2,3-butanediol in saccharomyces cerevisiae. J. Biotechnol. 2014, 192, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Dueber, J.E.; Wu, G.C.; Malmirchegini, G.R.; Moon, T.S.; Petzold, C.J.; Ullal, A.V.; Prather, K.L.J.; Keasling, J.D. Synthetic protein scaffolds provide modular control over metabolic flux. Nat. Biotechnol. 2009, 27, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.S.; Dueber, J.E.; Shiue, E.; Prather, K.L.J. Use of modular, synthetic scaffolds for improved production of glucaric acid in engineered E. coli. Metab. Eng. 2010, 12, 298–305. [Google Scholar] [CrossRef]
- Agapakis, C.M.; Ducat, D.C.; Boyle, P.M.; Wintermute, E.H.; Way, J.C.; Silver, P.A. Insulation of a Synthetic hydrogen metabolism circuit in bacteria. J. Biol. Eng. 2010, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, O. Synthetic Scaffolds increased resveratrol biosynthesis in engineered yeast cells. J. Biotechnol. 2012, 157, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.M.; Mazumdar, S.; Lee, S.W.; Jung, M.Y.; Lim, J.H.; Seo, S.W.; Jung, G.Y.; Oh, M.K. Butyrate production in engineered escherichia coli with synthetic scaffolds. Biotechnol. Bioeng. 2013, 110, 2790–2794. [Google Scholar] [CrossRef]
- Le Vo, T.D.; Ko, J.S.; Park, S.J.; Lee, S.H.; Hong, S.H. Efficient gamma-aminobutyric acid bioconversion by employing synthetic complex between glutamate decarboxylase and glutamate/GABA Antiporter in engineered Escherichia coli. J. Ind. Microbiol. Biotechnol. 2013, 40, 927–933. [Google Scholar] [CrossRef]
- Zhao, S.; Jones, J.A.; Lachance, D.M.; Bhan, N.; Khalidi, O.; Venkataraman, S.; Wang, Z.; Koffas, M.A.G. Improvement of catechin production in Escherichia coli through combinatorial metabolic engineering. Metab. Eng. 2015, 28, 43–53. [Google Scholar] [CrossRef]
- Tran, K.-N.T.; Somasundaram, S.; Eom, G.T.; Hong, S.H. Efficient itaconic acid production via protein–protein scaffold introduction between GltA, AcnA, and CadA in recombinant Escherichia Coli. Biotechnol. Prog. 2019, 35, e2799. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, S.; Jeong, J.; Irisappan, G.; Kim, T.W.; Hong, S.H. Enhanced production of malic acid by co-localization of phosphoenolpyruvate carboxylase and malate dehydrogenase using synthetic protein scaffold in Escherichia coli. Biotechnol. Bioprocess Eng. 2020, 25, 39–44. [Google Scholar] [CrossRef]
- Han, G.H.; Seong, W.; Fu, Y.; Yoon, P.K.; Kim, S.K.; Yeom, S.J.; Lee, D.H.; Lee, S.G. Leucine zipper-mediated targeting of multi-enzyme cascade reactions to inclusion bodies in Escherichia Coli for enhanced production of 1-butanol. Metab. Eng. 2017, 40, 41–49. [Google Scholar] [CrossRef]
- Ye, L.; Zhu, X.; Wu, T.; Wang, W.; Zhao, D.; Bi, C.; Zhang, X. Optimizing the localization of astaxanthin enzymes for improved productivity. Biotechnol. Biofuels 2018, 11, 278. [Google Scholar] [CrossRef]
- Haslinger, K.; Prather, K.L.J. Heterologous caffeic acid biosynthesis in Escherichia Coli is affected by choice of tyrosine ammonia lyase and redox partners for bacterial cytochrome P450. Microb. Cell Fact. 2020, 19, 26. [Google Scholar] [CrossRef]
- Whitaker, W.R.; Dueber, J.E. Chapter nineteen–Metabolic Pathway Flux Enhancement by Synthetic Protein Scaffolding. In Synthetic Biology, Part A; Voigt, C., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 497, pp. 447–468. [Google Scholar]
- Horn, A.H.C.; Sticht, H. Synthetic protein scaffolds based on peptide motifs and cognate adaptor domains for improving metabolic productivity. Front. Bioeng. Biotechnol. 2015, 3, 1–7. [Google Scholar] [CrossRef]
- Singleton, C.; Howard, T.P.; Smirnoff, N. Synthetic metabolons for metabolic engineering. J. Exp. Bot. 2014, 65, 1947–1954. [Google Scholar] [CrossRef]
- Conrado, R.J.; Wu, G.C.; Boock, J.T.; Xu, H.; Chen, S.Y.; Avbelj, M.; Gaber, R.; Lebar, T.; Turns, J.; Hodnik, V.; et al. DNA–guided assembly of biosynthetic pathways promotes improved catalytic efficiency. Nucleic Acids Res. 2012, 40, 1879–1889. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA websuite. Nucleic Acids Res. 2008, 36, 70–74. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Delebecque, C.J.; Silver, P.A.; Lindner, A.B. Designing and using RNA scaffolds to assemble proteins in Vivo. Nat. Protoc. 2012, 7, 1797–1807. [Google Scholar] [CrossRef]
- Sachdeva, G.; Garg, A.; Godding, D.; Way, J.C.; Silver, P.A. In Vivo co-localization of enzymes on RNA Scaffolds increases metabolic production in a geometrically dependent manner. Nucleic Acids Res. 2014, 42, 9493–9503. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.M.; Geraldi, A.; Nguyen, T.T.; Lee, J.H.; Lee, J.Y.; Cho, B.-K.; Kim, S.C. MRNA Engineering for the efficient chaperone-mediated co-translational folding of recombinant proteins in Escherichia coli. Int. J. Mol. Sci. 2019, 20, 3163. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Qiu, X.; Zhu, L.; Wu, X.; Zhang, Y.; Zhu, Q.; Fan, D.; Zhu, C.; Zhang, D. Spatial organization of heterologous metabolic system in Vivo based on TALE. Sci. Rep. 2016, 6, 26065. [Google Scholar] [CrossRef]
- Xie, S.; Qiu, X.; Zhu, L.; Zhu, C.; Liu, C.; Wu, X.; Zhu, L.; Zhang, D. Assembly of TALE-based DNA scaffold for the enhancement of exogenous multi-enzymatic pathway. J. Biotechnol. 2019, 296, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rossi, J. Aptamers as Targeted therapeutics: Current potential and challenges. Nat. Rev. Drug Discov. 2017, 16, 181–202. [Google Scholar] [CrossRef]
- Chen, M. RNA Scaffold: Designed to co-localize enzymes. Methods Mol. Biol. 2015, 1316, 105–112. [Google Scholar] [CrossRef]
- Delebecque, C.J.; Lindner, A.B.; Silver, P.A.; Aldaye, F.A. Organization of Intracellular reactions with rationally designed RNA assemblies. Science 2011, 333, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.S. A Guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Zalatan, J.G.; Lee, M.E.; Almeida, R.; Gilbert, L.A.; Whitehead, E.H.; La Russa, M.; Tsai, J.C.; Weissman, J.S.; Dueber, J.E.; Qi, L.S.; et al. Engineering complex synthetic transcriptional programs with CRISPR RNA scaffolds. Cell 2015, 160, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, D.; Cox, T.; Platt, R.J.; Zhang, F. Therapeutic genome editing: Prospects and challenges. Nat. Med. 2015, 21, 121–131. [Google Scholar] [CrossRef]
- Hong, F.; Zhang, F.; Liu, Y.; Yan, H. DNA Origami: Scaffolds for Creating Higher Order Structures. Chem. Rev. 2017, 117, 1258–12640. [Google Scholar] [CrossRef]
- Lin, C.X.; Rinker, S.; Wang, X.; Liu, Y.; Seeman, N.C.; Yan, H. In Vivo Cloning of artificial DNA nanostructures. Proc. Natl. Acad. Sci. USA 2008, 105, 17626–17631. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Scaffolded Enzyme | Domains | Host | Maximum Fold Increase 1 | Reference |
|---|---|---|---|---|---|
| Domains and Ligands from Metazoan Cells | |||||
| Mevalonate | Acetoacetyl-CoA thiolase, hydroxymethylglutaryl-CoA synthase, hydroxymethylglutaryl-CoA reductase | GBD, SH3, PDZ | Escherichia coli | 77 | [32] |
| d-Glucaric acid | Myo-inositol-1-phosphate synthase, myo-inositol oxygenase | SH3, PDZ | E. coli | 3 | [32] |
| d-Glucaric acid | Myo-inositol-1-phosphate synthase, myo-inositol oxygenase, uronate dehydrogenase | GBD, SH3, PDZ | E. coli | 5 | [33] |
| Hydrogen (H2) | [Fe-Fe]-hydrogenase, ferredoxin | SH3, PDZ | E. coli | 3 | [34] |
| Resveratrol | 4-Coumarate- CoA ligase, stilbene synthase | SH3, PDZ | Saccharomyces cerevisiae | 5 | [35] |
| Butyrate | 3-Hydroxybutyryl-CoA dehydrogenase, 3-hydroxybutyryl-CoA dehydratase, trans-enoyl-coenzyme A reductase, acyl-CoA thioesterase II | GBD, SH3, PDZ | E. coli | 3 | [36] |
| Gamma-aminobutyric acid (GABA) | Glutamate decarboxylase, glutamate/GABA antiporter | SH3 | E. coli | 2.5 | [37] |
| Catechin | Flavanone 3-hydroxylase, dihydroflavonol 4-reductase, leucoanthocyanidin reductase | GBD, SH3, PDZ | E. coli | 1.3 | [38] |
| Itaconic acid | Citrate synthase, acotinase, cis-acotinic acid decarboxylase | GBD, SH3, PDZ | E. coli | 3.8 | [39] |
| Malic acid | Phosphoenolpyruvate carboxylase, malate dehydrogenase | SH3 | E. coli | 3.6 | [40] |
| Indigoidine | Glutamine synthetase, indigoidine synthase (indC), helper protein of IndC (IndB) | GBD, SH3, PDZ | E. coli | 2.1 | |
| Cohesin and dockerin domains | |||||
| NADH | Alcohol dehydrogenase, formaldehyde dehydrogenase, formate dehydrogenase | Cohesin–dockerin pairs from Clostridium cellulolyticum, Clostridium thermocellum, and Ruminococcus flavefaciens | S. cerevisiae | 5 | [30] |
| 2,3-Butanediol | Acetolactate synthase, acetolactate decarboxylase, 2,3-butanediol dehydrogenase | Cohesin–dockerin pair from C. thermocellum | S. cerevisiae | 1.4 | [31] |
| Other domains | |||||
| 1-Butanol | 3-Hydroxybutyryl-CoA dehydrogenase, crotonase, butyryl-CoA dehydrogenase, butylaldehyde/butanol dehydrogenase | Leucine zipper domains | E. coli | 1.5 | [41] |
| Astaxanthin | β-Carotene ketolase, β-carotene hydroxylase | Glycerol uptake facilitator protein | E. coli | 2.2 | [42] |
| Caffeic acid | ferredoxin, ferredoxin reductase, cytochrome P450 enzyme | PCNA of Sulfolobus solfataricus P2 | E. coli | 8 | [43] |
| Product | Scaffolded Enzyme/Protein | Nucleic Acid-Binding Protein | Host | Maximum Fold Increase 1/Other Result | Reference |
|---|---|---|---|---|---|
| RNA Scaffold Systems | |||||
| Hydrogen (H2) | [Fe-Fe]-hydrogenase, ferredoxin | PP7, MS2 | Escherichia coli | 48 | [50] |
| Pentadecane | Acyl-ACP reductase, aldehyde deformylating oxygenase | BIV-Tat, PP7 | E. coli | 1.8 | [51] |
| Succinate | Carbonic anhydrase, pyruvate carboxylase, malate dehydrogenase, NAD-formate dehydrogenase | RevR11Q, PP7, BIV-Tat, Lambda N | E. coli | 2.6 | [51] |
| E. coli UDP-glucose dehydrogenase (UGD), Anti-Ras single chain variable fragment (anti-Ras ScFv) | DnaJ chaperone | KH3 | E. coli | 90% of expressed UGD and 80% of expressed anti-Ras ScFv were soluble | [52] |
| DNA scaffold systems | |||||
| trans- Resveratrol | 4-Coumarate–CoA ligase, stilbene synthase | Zif268, PBSII (3 fingers ZFP) | E. coli | 3 | [47] |
| 1,2- Propanediol | Methylglyoxal synthase, 2,5-diketo-d-gluconic acid reductase, glycerol dehydrogenase | ZFa, ZFb, ZFc (3 fingers ZFP) | E. coli | 4 | [47] |
| Mevalonate | Acetoacetyl-CoA thiolase, hydroxymethylglutaryl- CoA synthase, hydroxymethylglutaryl-CoA reductase | ZFa, ZFb, ZFc (3 fingers ZFP) | E. coli | 3 | [47] |
| l-Threonine | Homoserine dehydrogenase, homoserine kinase, threonine synthase | ADB1, ADB2, ADB3 (4 fingers ZFP) | E. coli | Reduced production time by more than 50% | [9] |
| n-Alkane | Acyl-ACP reductase, aldehyde deformylating oxygenase | ADB2, ADB4 (4 fingers ZFP) | E. coli | 8.8 | [25] |
| Indole-3-acetic acid | Tryptophan-2-mono-oxygenase, indole-3-acetamide hydrolase | TALE1, TALE2 | E. coli | 9.6 | [53] |
| Mevalonate | Acetoacetyl-CoA thiolase, hydroxymethylglutaryl- CoA synthase, hydroxymethylglutaryl-CoA reductase | TALE1, TALE2, TALE3 | E. coli | 3.7 | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geraldi, A.; Khairunnisa, F.; Farah, N.; Bui, L.M.; Rahman, Z. Synthetic Scaffold Systems for Increasing the Efficiency of Metabolic Pathways in Microorganisms. Biology 2021, 10, 216. https://doi.org/10.3390/biology10030216
Geraldi A, Khairunnisa F, Farah N, Bui LM, Rahman Z. Synthetic Scaffold Systems for Increasing the Efficiency of Metabolic Pathways in Microorganisms. Biology. 2021; 10(3):216. https://doi.org/10.3390/biology10030216
Chicago/Turabian StyleGeraldi, Almando, Fatiha Khairunnisa, Nadya Farah, Le Minh Bui, and Ziaur Rahman. 2021. "Synthetic Scaffold Systems for Increasing the Efficiency of Metabolic Pathways in Microorganisms" Biology 10, no. 3: 216. https://doi.org/10.3390/biology10030216
APA StyleGeraldi, A., Khairunnisa, F., Farah, N., Bui, L. M., & Rahman, Z. (2021). Synthetic Scaffold Systems for Increasing the Efficiency of Metabolic Pathways in Microorganisms. Biology, 10(3), 216. https://doi.org/10.3390/biology10030216