Climate Mitigation through Biological Conservation: Extensive and Valuable Blue Carbon Natural Capital in Tristan da Cunha’s Giant Marine Protected Zone

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
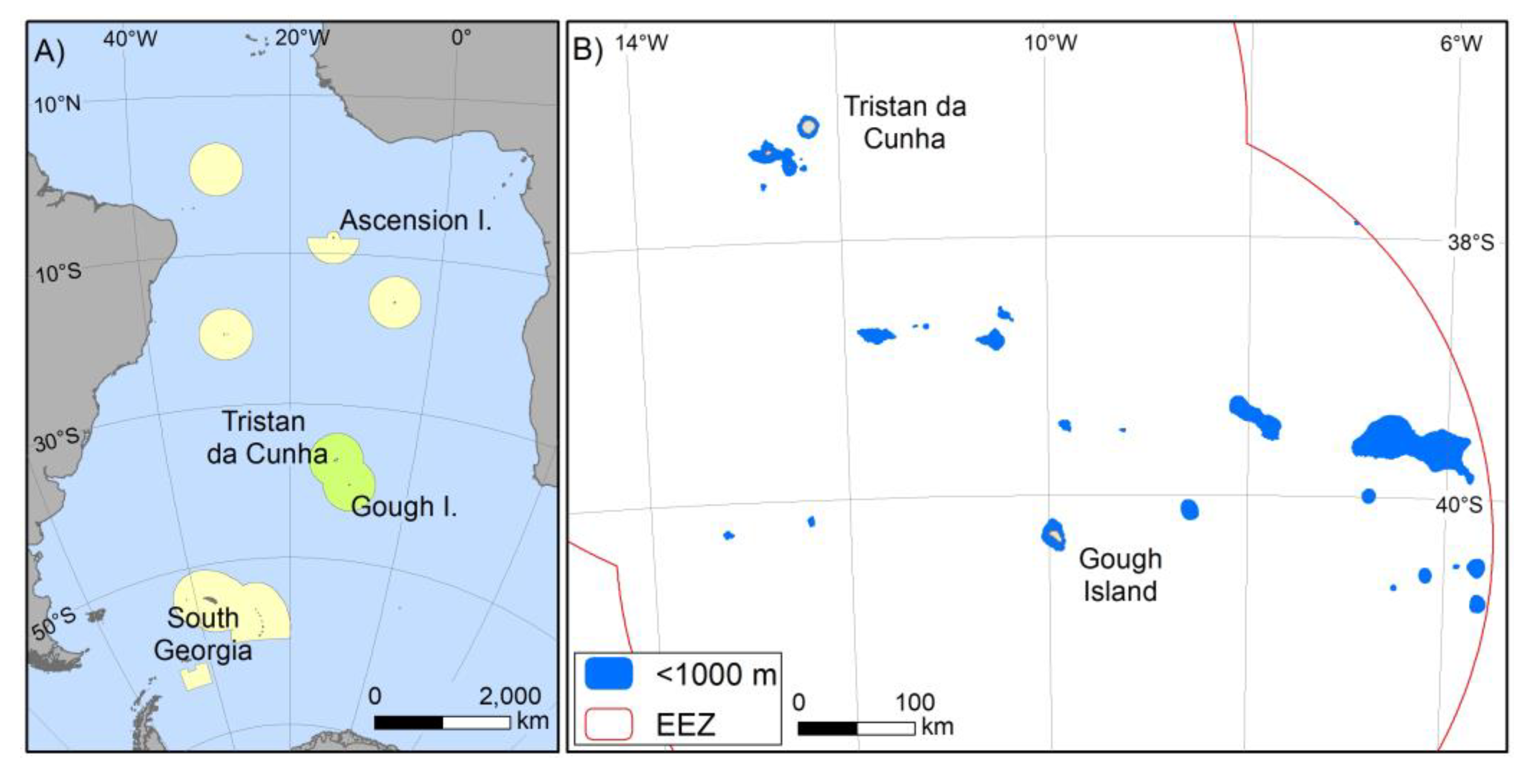
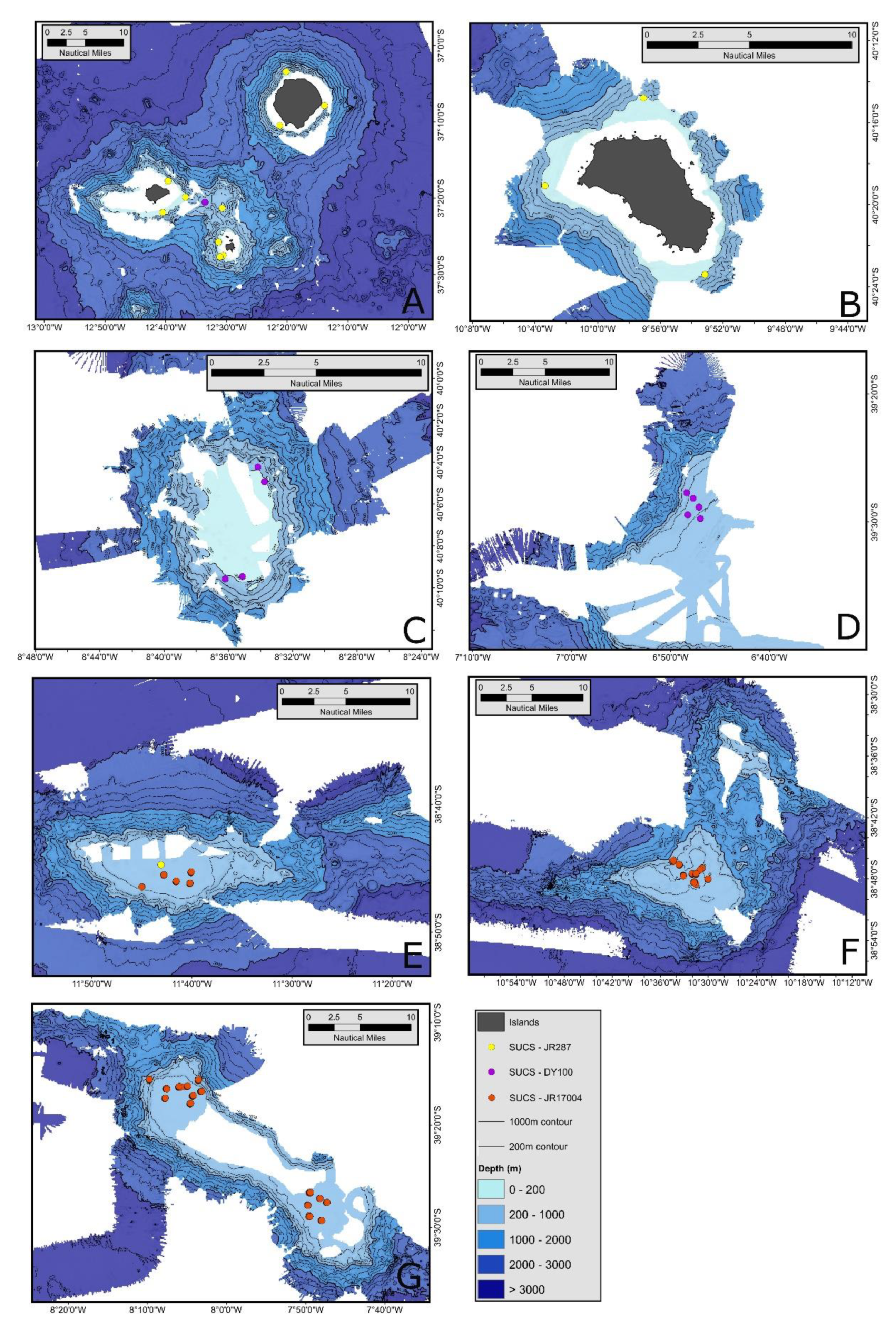
2. Materials and Methods
3. Results
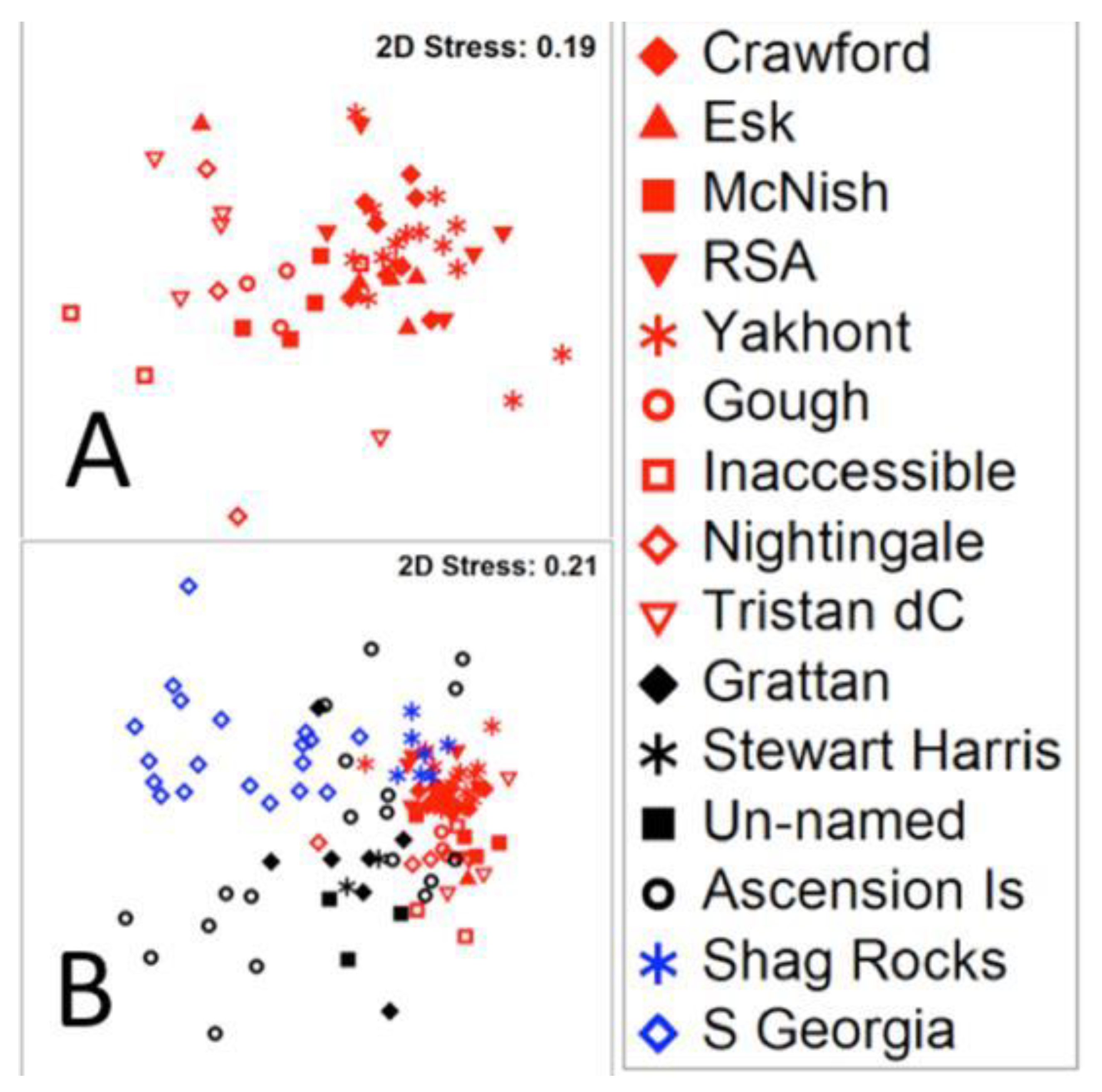
3.1. Variability of Seabed Biological Carbon Storage and Influences on It
3.2. Valuing Tristan da Cunha EEZ/MPA Carbon Sequestration
4. Discussion
4.1. How and Why Does Mid Ocean Blue Carbon Natural Capital Vary?
4.2. Ongoing Blue Carbon Ecosystem Services and Sequestration
4.3. What Now for VLMPAs and Mid Ocean Archipelagos?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Constanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- De Groot, R.; Brander, R.; van der Ploeg, S.; Costanza, R.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst. Serv. 2012, 1, 50–61. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E.; Arneth, A.; Bai, X.; Barnes, D.; Burrows, M.; Chan, L.; Cheung, W.L.; et al. IPBES-IPCC Co-Sponsored Workshop Report on Biodiversity and Climate Change; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES): Bonn, Germany; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2021. [Google Scholar]
- Sala, E.; Mayorga, J.; Bradley, D.; Cabral, R.B.; Atwood, T.B.; Auber, A.; Cheung, W.; Costello, C.; Ferretti, F.; Friedlander, A.M.; et al. Protecting the global ocean for biodiversity, food and climate. Nature 2021, 592, 397–402. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global declines in human-driven mangrove loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Duarte, C.M. Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience 2016, 9, 737–742. [Google Scholar] [CrossRef]
- Queirós, A.M.; Stephens, N.; Widdicombe, S.; Tait, K.; McCoy, S.J.; Ingels, J.; Rühl, S.; Airs, R.; Beesley, A.; Carnovale, G.; et al. Connected macroalgal-sediment Syst.: Blue carbon and food webs in the deep coastal ocean. Ecol. Monogr. 2019, 89, e01366. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Wernberg, T. Substantial blue carbon in overlooked Australian kelp forests. Sci. Rep. 2020, 10, 12341. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Wheeler, A.J.; Freiwald, A.; Cairns, S.J. Cold-Water Corals. The Biology and Geology of Deep-Sea Coral Habitats; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- La Bianca, G.; Tillin, H.; Hodgson, B.; Erni-Cassola, G.; Howell, K.; Rees, S. Ascension Island Natural Capital Assessment: Marine Ecosystem Services Report; Natural Capital in the UK’s Overseas Territories Report Series—Supplementary Report (South Atlantic Region); JNCC: Peterborough, UK, 2018. [Google Scholar]
- Bridges, A.E.; Barnes, D.K.A.; Bell, J.B.; Ross, R.E.; Howell, K.L. Benthic community composition of South Atlantic seamounts. Front. Mar. Sci. 2021, 8, 1530. [Google Scholar] [CrossRef]
- Foley, N.; van Rensburg, T.; Armstrong, C. The ecological and economic value of cold-water coral ecosystems. Ocean Coast. Manag. 2010, 53, 313–326. [Google Scholar] [CrossRef]
- Jobstvogt, N.; Townsend, M.; Witte, U.; Hanley, N. How Can we identify and communicate the ecological value of deep-sea ecosystem services? PLoS ONE 2014, 9, e100646. [Google Scholar] [CrossRef]
- Rogers, A.; Baco, A.; Griffiths, H.; Hart, T.; Hall-Spencer, J. Corals on seamounts. In Seamounts: Ecology, Fisheries & Conservation; Pitcher, T.J., Morato, T., Hart, P.J.B., Clark, M.R., Haggan, N., Santos, R.S., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 1–527. [Google Scholar] [CrossRef]
- Kenchington, E.; Murillo, F.J.; Lirette, C.; Sacau, M.; Koen-Alonso, M.; Kenny, A.; Ollerhead, N.; Wareham, V.; Beazley, L. Kernel density surface modelling as a means to identify significant concentrations of vulnerable marine ecosystem indicators. PLoS ONE 2014, 9, e109365. [Google Scholar] [CrossRef]
- Kenny, A.J.; Cambell, N.; Koen-Alonso, M.; Pepin, P.; Diz, D. Delivering sustainable fisheries through adoption of a risk-based framework as part of an ecosystem approach to fisheries management. Mar. Policy 2018, 93, 232–240. [Google Scholar] [CrossRef]
- Kaiser, M.J. Fish in deep-water coral habitats. Science 2004, 304, 1595. [Google Scholar] [CrossRef][Green Version]
- Bax, N.; Sands, C.; Gogarty, B.; Downey, R.V.; Moreau, C.V.E.; Moreno, B.; Held, C.; Lund Paulsen, M.; McGee, J.; Haward, M.; et al. Perspective: Increasing blue carbon around Antarctica is an ecosystem service of considerable societal and economic value worth protecting. Glob. Chang. Biol. 2021, 27, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.B.; Guijarro-Garcia, E.; Kenny, A. Demersal fishing in areas beyond national jurisdiction: A comparative analysis of regional fisheries management organizations. Front. Mar. Sci. 2019, 6, 596. [Google Scholar] [CrossRef]
- Pham, C.K.; Vandeperre, F.; Menezes, G.; Porterio, F.; Isidro, E.; Morato, T. The importance of deep-sea vulnerable marine ecosystems for demersal fish in the Azores. Deep Sea Res. Part I 2015, 96, 80–88. [Google Scholar] [CrossRef]
- McLeod, G.L.; Chmura, S.; Bouillon, R.; Salm, M.; Bjork, C.M.; Duarte, C.E.; Lovelock, W.; Schlesinger, W.H.; Silliman, B.R. A blue print for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Environ. Ecol. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Wylie, L.; Sutton-Grier, A.E.; Moore, A. Keys to successful blue carbon projects: Lessons learned from global case studies. Mar. Policy 2016, 65, 76–84. [Google Scholar] [CrossRef]
- White, M.; Bashmachnikov, I.; Aristegui, J.; Martins, A. Physical processes and seamount productivity. Fish Aquat. Res. Ser. 2007, 12, 65–84. [Google Scholar]
- Barnes, D.K.A.; Sands, C.J. Functional group diversity is key to Southern Ocean benthic carbon pathways. PLoS ONE 2017, 12, e0179735. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.A.; Sands, C.J.; Richardson, A.; Smith, N. Extremes in benthic ecosystem services; Blue Carbon natural capital shallower than 1000 m in isolated, small and young Ascension island’s EEZ. Front. Mar. Sci. 2019, 6, 663. [Google Scholar] [CrossRef]
- Souster, T.; Barnes, D.K.A.; Hopkins, J. Variation in zoobenthic blue carbon in the Arctic’s Barents Sea shelf sediments. Phil. Trans. R. Soc. A 2020, 378, 20190362. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Langenkämper, D.; Zurowietz, M.; Schoening, T.; Nattkemper, T.W. BIIGLE 2.0—Browsing and annotating large marine image collections. Front. Mar. Sci. 2017, 4, 83. [Google Scholar] [CrossRef]
- Caselle, J.E.; Hamilton, S.L.; Davis, K.; Thompson, C.D.; Turchik, A.; Jenkinson, R.; Simpson, D.; Sala, E. First quantification of subtidal community structure at Tristan da Cunha Islands in the remote South Atlantic: From kelp forests to the deep sea. PLoS ONE 2018, 13, e0195167. [Google Scholar] [CrossRef] [PubMed]
- North, W.J. Growth of individual fronds of the mature giant kelp Macrocystis. In The Biology of Giant Kelp Beds (Macrocystis) in California; North, W.J., Lehre, J., Eds.; Cramer: Lehre, Germany, 1971; pp. 123–168. [Google Scholar]
- Diekmann, G.S. Aspects of Growth and Production of Laminaria pallida off the Cape Peninsula. Master’s Thesis, University of Capetown, Capetown, South Africa, 1978. [Google Scholar]
- Wickham, S.B.; Darimont, C.T.; Reynolds, J.D.; Staromski, B.M. Species-specific wet-dry mass calibrations for dominant Northeastern Pacific Ocean macroalgae and seagrass. Aquat. Bot. 2019, 152, 27–31. [Google Scholar] [CrossRef]
- Mora-Soto, A.; Palacios, M.; Macaya, E.C.; Gómez, I.; Huovinen, P.; Pérez-Matus, A.; Young, M.; Golding, N.; Toro, M.; Yaqub, M.; et al. A high-resolution global map of giant Kelp (Macrocystis pyrifera) forests and intertidal Green Algae (Ulvophyceae) with sentinel-2 imagery. Remote Sens. 2020, 12, 694. [Google Scholar] [CrossRef]
- Krumhansl, K.; Scheibling, R. Production and fate of kelp detritus. Mar. Ecol. Prog. Ser. 2012, 467, 281–302. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Lavery, P.; Serrano, O.; Marbà, N.; Masque, P.; Duarte, C.M. Sequestration of macroalgal carbon: The elephant in the Blue Carbon room. Biol. Lett. 2018, 14, 142018023620180236. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.; Bellgrove, A.; Macreadie, P.I.; Petrou, K.; Beardall, J.; Steven, A.; Ralph, P.J. Can macroalgae contribute to blue carbon? An Australian perspective. Limnol. Oceanogr. 2015, 60, 1689–1706. [Google Scholar] [CrossRef]
- De Clippele, L.H.; Rovelli, L.; Ramiro-Sánchez, B.; Kazanidis, G.; Vad, J.; Turner, S.; Glud, R.N.; Roberts, J.M. Mapping cold-water coral biomass: An approach to derive ecosystem functions. Coral Reefs 2021, 40, 215–231. [Google Scholar] [CrossRef]
- Carbon Pricing Leadership Coalition. Report of the High-Level Commission on Carbon Pricing and Competitiveness; World Bank: Washington, DC, USA, 2019; Available online: https://openknowledge.worldbank.org/handle/10986/32419 (accessed on 24 November 2021).
- Vecsei, A. A new estimate of global reefal carbonate production including the fore-reefs. Glob. Planet. Chang. 2004, 43, 1–18. [Google Scholar] [CrossRef]
- Roberts, C.M. Deep impact: The rising toll of fishing in the deep sea. Trends Ecol. Evol. 2002, 17, 242–245. [Google Scholar] [CrossRef]
- Santillo, D.; Johnston, P.A. Marine protected areas (MPAs) as management tools to conserve seamount ecosystems. In Proceedings of the International Conference on Governance and Management of Deep-Sea Fisheries, Queenstown, New Zealand, 1–5 December 2003; FAO Fisheries Proceedings. FAO: Rome, Italy, 2005; pp. 560–578. [Google Scholar]
- Howell, K.L.; Davies, J.S.; Narayanaswamy, B.E. Identifying deep-sea megafaunal epibenthic assemblages for use in habitat mapping and marine protected area network design. J. Mar. Biol. Assoc. UK 2010, 90, 33–68. [Google Scholar] [CrossRef]
- Clark, M.R.; Bowden, D.A. Seamount biodiversity: High variability both within and between seamounts in the Ross Sea region of Antarctica. Hydrobiology 2015, 761, 161–180. [Google Scholar] [CrossRef]
- Ross, R.E.; Howell, K.L. Use of predictive habitat modelling to assess the distribution and extent of the current protection of ‘listed’ deep-sea habitats. Divers. Distrib. 2013, 19, 433–445. [Google Scholar] [CrossRef]
- Ross, L.K.; Ross, R.E.; Stewart, H.A.; Howell, K.L. The influence of data resolution on predicted distribution and estimates of extent of current protection of three ‘listed’ deep-sea habitats. PLoS ONE 2015, 10, e0140061. [Google Scholar] [CrossRef] [PubMed]
- Smale, D.A.; Moore, P.J.; Queirós, A.M.; Higgs, N.D.; Burrows, M.T. Appreciating interconnectivity between habitats is key to blue carbon management. Front. Ecol. Environ. 2018, 16, 71–73. [Google Scholar] [CrossRef]
- Lundsten, L.; Barry, J.; Cailliet, G.; Clague, D.; DeVogelaere, A.; Geller, J. Benthic invertebrate communities on three seamounts off southern and central California, USA. Mar. Ecol. Prog. Ser. 2019, 374, 23–32. [Google Scholar] [CrossRef]
- Levin, L.A.; Thomas, C.L. The influence of hydrodynamic regime on infaunal assemblages inhabiting carbonate sediments on central Pacific seamounts. Deep Sea Res. 1989, 36, 1897–1915. [Google Scholar] [CrossRef]
- O’Hara, T.D.; Tittensor, D.P. Environmental drivers of ophiuroid species richness on seamounts. Mar. Ecol. 2010, 31 (Suppl. S1), 26–38. [Google Scholar] [CrossRef]
- Morgan, N.B.; Goode, S.; Roark, E.B.; Baco, A.R. Fine scale assemblage structure of benthic invertebrate Megafauna on the north pacific seamount Mokumanamana. Front. Mar. Sci. 2019, 6, 715. [Google Scholar] [CrossRef]
- Queirós, A.M.; Hiddink, J.G.; Kaiser, M.J.; Hinz, H. Effects of chronic bottom trawling disturbance on benthic biomass, production and size spectra in different habitats. J. Exp. Mar. Biol. Ecol. 2006, 335, 91–103. [Google Scholar] [CrossRef]
- Watson, R.; Kitchingman, A.; Cheung, W. Catches from world seamount fisheries. In Seamounts: Ecology, Fisheries & Conservation; Pitcher, T.J., Morato, T., Hart, P.J.B., Clark, M.R., Haggan, N., Santos, R.S., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007. [Google Scholar]
- Clark, M.R.; Rowden, A.A.; Schlacher, T.; Williams, A.; Consalvey, M.; Stocks, K.I.; Rogers, A.D.; O’Hara, T.D.; White, M.; Shank, T.M.; et al. The ecology of seamounts: Structure, function, and human impacts. Ann. Rev. Mar. Sci. 2010, 2, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Van Oevelen, D.; Duineveld, G.; Lavaleye, M.; Mienis, F.; Soetaert, K.; Heip, C.H.R. The cold-water coral community as a hot spot for carbon cycling on continental margins: A food-web analysis from Rockall Bank (northeast Atlantic). Liminol. Oceanogr. 2009, 54, 1829–1844. [Google Scholar] [CrossRef]
- Sabatier, P.; Reyss, J.L.; Hall-Spencer, J.M.; Colin, C.; Frank, N. 210Pb-226Ra chronology reveals rapid growth rate of Madrepora oculata and Lophelia pertusa on world’s largest cold-water coral reef. Biogeosciences 2012, 9, 1253–1265. [Google Scholar] [CrossRef]
- Mallela, J. Calcification by reef-building sclerobionts. PLoS ONE 2013, 8, e60010. [Google Scholar] [CrossRef]
- Lunden, J.J.; McNicholl, C.G.; Sears, C.R.; Morrison, C.L.; Cordes, E.E. Acute survivorship of the deep-sea coral Lophelia pertusa from the Gulf of Mexico under acidification, warming, and deoxygenation. Front. Mar. Sci. 2014, 1, 78. [Google Scholar] [CrossRef]
- Koslow, J.; Gowlett-Holmes, K.; Lowry, J.; O’Hara, T.; Poore, G.; Williams, A. Seamount benthic macrofauna off southern Tasmania: Community structure and impacts of trawling. Mar. Ecol. Prog. Ser. 2001, 213, 111–125. [Google Scholar] [CrossRef]
- Nolan, E.; Barnes, D.; Brown, J.; Downes, K.; Enderlein, P.; Gowland, E.; Brickle, P. Biological and physical characterization of the seabed surrounding Ascension Island from 100–1000 m. J. Mar. Biol. Ass. UK 2017, 97, 647–659. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | SP | SC | SS | DC | DV | DS | GC | PS | PC | PM | PL | PA | FS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Islands | 13 | 16.3 | 0.3 | * | * | 0.8 | 2.2 | 17.4 | 70 | * | 2.2 | 1.5 | 1.1 |
| Yakhont | 4.6 | 45.1 | 0.2 | * | * | 4.7 | 9.7 | 83.9 | 21.6 | 0.6 | 1.1 | 1.1 | 18.7 |
| Crawford | 18.7 | 43.5 | * | * | * | 4.4 | 15.5 | 81 | 17.5 | 0.3 | 1.4 | 6.2 | 10.2 |
| Esk | 11.9 | 32.5 | 0.2 | * | * | 5 | 8.7 | 31 | 68.6 | 1.2 | 0.9 | 3.5 | 6.1 |
| McNish | 9.5 | 36.8 | * | * | * | 36.3 | 2.7 | 25 | 43.1 | 0.9 | 4 | 1.8 | 1.1 |
| RSA | 6.4 | 32.3 | 1 | * | * | 15.3 | 5 | 16.9 | 14.6 | * | 1.1 | 0.4 | 15.1 |
| Locality | Crawford | McNish | RSA | Esk | Gough | Tristan dC | Nightingale | Inaccessible |
|---|---|---|---|---|---|---|---|---|
| Yakhont | x | x | x | x | x | 0.72 ** | 0.83 ** | 0.68 * |
| Crawford | 0.63 * | 0.35 * | x | 0.72 * | 0.78 ** | 0.88 * | x | |
| McNish | x | x | x | 0.55 * | x | x | ||
| RSA | x | x | x | x | x | |||
| Esk | x | x | x | x | ||||
| Gough | x | x | x | |||||
| Tristan dC | x | x | ||||||
| Nightingale | x | |||||||
| Inaccessible |
| Locality | SP | SC | SS | DC | DV | DS | GC | PS | PC | PM | PL | PA | FS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (a) islands | 0.83 | 1.45 | 1.1 | 0.95 | 0.85 | 1.53 | 2.8 | 1.51 | 2.86 | 0.91 | 3.79 | 1.27 | 0.8 |
| seamount | 0.88 | 1.41 | 1.08 | 0.93 | 0.87 | 2.07 | 2.8 | 1.38 | 2.94 | 0.63 | 3.36 | 1.6 | 0.8 |
| overall | 0.86 | 1.43 | 1.09 | 0.95 | 0.85 | 1.83 | 2.8 | 1.43 | 2.88 | 0.78 | 3.58 | 1.44 | 0.8 |
| (b) Islands | 11 | 23 | 0.3 | * | * | 1.4 | 6.8 | 23 | 191 | * | 8 | 2.2 | 0.8 |
| Yakhont | 4.1 | 65 | 0.2 | * | * | 8.7 | 38 | 120 | 59 | 0.5 | 3.7 | 1.6 | 15 |
| Craw | 17 | 65 | * | * | * | 7.9 | 23 | 119 | 53 | 0.2 | 5.8 | 9 | 8.3 |
| Esk | 8.9 | 46 | 0.2 | * | * | 9.8 | 26 | 40 | 203 | 1.5 | 2 | 5.2 | 4 |
| McNish | 9 | 54 | * | * | * | 86 | 3.9 | 35 | 134 | * | 16 | 2.3 | 0.2 |
| RSA | 5.1 | 55 | 1.1 | * | * | 16 | 16 | 25 | 26 | * | 2.5 | 0.8 | 12 |
| Source | DF | Adj SS | Adj MS | F | p |
|---|---|---|---|---|---|
| Rugosity | 6 | 3,117,526 | 519,588 | 7.1 | 0.001 |
| Substrate | 8 | 3,773,735 | 471,717 | 6.4 | 0.001 |
| Richness | 9 | 3,993,682 | 443,742 | 6.1 | 0.001 |
| Locality | 9 | 2,886,060 | 320,673 | 4.4 | 0.001 |
| Site | 14 | 3,766,570 | 269,041 | 3.7 | 0.001 |
| Temperature | 1 | 235,935 | 235,935 | 3.2 | 0.073 |
| Oxygen | 1 | 230,455 | 230,455 | 3.2 | 0.076 |
| Chlorophyll | 1 | 22,701 | 22,701 | 0.3 | 0.578 |
| Salinity | 1 | 127 | 127 | 0.0 | 0.967 |
| Error | 936 | 68,533,741 | 73,220 | ||
| Total | 986 | 103,092,216 |
| Area Measure | Area km2 | Zb tC/km2 | Zb Million tC | Seq tC/km2 | Seq Million tC |
|---|---|---|---|---|---|
| Planar Area | |||||
| Islands | 659.5 | 200.6 | 0.13 | 28 | 0.019 |
| seamounts | 4859 | 274 | 1.33 | 25 | 0.124 |
| total | 5518 | 1.46 | 0.143 | ||
| Surface Area | |||||
| Islands | 1253 | 200.6 | 0.25 | 28 | 0.035 |
| seamounts | 7288 | 274 | 1.99 | 25 | 0.182 |
| total | 8541 | 2.25 | 0.217 | ||
| Locality | Area km2 | Density M × 1000 | Density L × 1000 | C Store Tonnes | Seq °C |
|---|---|---|---|---|---|
| Gough | 2.89 | 54 | 237 | 23.27 | 2.6 |
| Tristan dC | 0.95 | 51 | 543 | 15.11 | 1.6 |
| Inaccessible | 0.69 | 54 | 387 | 8.25 | 0.5 |
| Nightingale | 0.87 | 44 | 518 | 13.06 | 0.8 |
| Total | 5.4 | 5.5 |
| Seq Million tC | CO2 Equiv | £ Million | oES Million tC/year | oES Million tCO2 | oES £ Million | |
|---|---|---|---|---|---|---|
| TdC islands | 0.04 | 0.13 | 3.9 | 0.005 | 0.018 | 0.54 |
| seamounts | 0.18 | 0.68 | 20.5 | 0.026 | 0.095 | 2.87 |
| total | 0.22 | 0.80 | 24.4 | 0.031 | 0.114 | 3.42 |
| Ascension total | 0.01 | 0.03 | 1 | <0.001 | 0.002 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnes, D.K.A.; Bell, J.B.; Bridges, A.E.; Ireland, L.; Howell, K.L.; Martin, S.M.; Sands, C.J.; Mora Soto, A.; Souster, T.; Flint, G.; et al. Climate Mitigation through Biological Conservation: Extensive and Valuable Blue Carbon Natural Capital in Tristan da Cunha’s Giant Marine Protected Zone. Biology 2021, 10, 1339. https://doi.org/10.3390/biology10121339
Barnes DKA, Bell JB, Bridges AE, Ireland L, Howell KL, Martin SM, Sands CJ, Mora Soto A, Souster T, Flint G, et al. Climate Mitigation through Biological Conservation: Extensive and Valuable Blue Carbon Natural Capital in Tristan da Cunha’s Giant Marine Protected Zone. Biology. 2021; 10(12):1339. https://doi.org/10.3390/biology10121339
Chicago/Turabian StyleBarnes, David K. A., James B. Bell, Amelia E. Bridges, Louise Ireland, Kerry L. Howell, Stephanie M. Martin, Chester J. Sands, Alejandra Mora Soto, Terri Souster, Gareth Flint, and et al. 2021. "Climate Mitigation through Biological Conservation: Extensive and Valuable Blue Carbon Natural Capital in Tristan da Cunha’s Giant Marine Protected Zone" Biology 10, no. 12: 1339. https://doi.org/10.3390/biology10121339
APA StyleBarnes, D. K. A., Bell, J. B., Bridges, A. E., Ireland, L., Howell, K. L., Martin, S. M., Sands, C. J., Mora Soto, A., Souster, T., Flint, G., & Morley, S. A. (2021). Climate Mitigation through Biological Conservation: Extensive and Valuable Blue Carbon Natural Capital in Tristan da Cunha’s Giant Marine Protected Zone. Biology, 10(12), 1339. https://doi.org/10.3390/biology10121339