Abstract
The impact of steroid hormones, and particularly estradiol, on human microbiota could be recognized as a substantial part of human-microbiota interactions. However, an area that remains poorly investigated is that of the skin and vaginal microbial communities and biofilms, which contain non-pathogenic bacteria of phyla Firmicutes and Actinobacteria, especially probiotic bacteria of the genus Lactobacillus and the widespread, safe skin genus, Micrococcus. Experiments with Lactobacillus paracasei AK508 and Micrococcus luteus C01 biofilms on PTFE cubes showed dose-dependent effects of estradiol at concentrations of 0.22 nM and 22 nM. The hormone mostly inhibits L. paracasei growth and stimulates M. luteus. The presented studies of colony-forming unit (CFU) amountsand cell aggregation in biofilms on glass fiber filters showed the same general tendencies. Estradiol generally increased the aggregation of cells in monospecies communities and potentially changed the synthesis of antibacterial metabolites in L. paracasei. The balance between two bacteria in mixed-species biofilms depended on the initial adhesion stage, and when this stage was reduced, micrococci were more resistant to the antagonistic action of L. paracasei. Moreover, in mixed-species biofilms, the effect of estradiol on lactobacilli altered from inhibition to stimulation, potentially due to the presence of M. luteus. At the same time, ethanol as a solvent for estradiol at the concentration 0.6% acted mostly as an antagonist of the hormone and had an opposite effect on bacteria; nevertheless, the overlapping of ethanol and estradiol effects was shown to be minimal. The data obtained prove the complexity of microbial interactions and the regulatory effect of estradiol on commensal bacteria biofilms.
1. Introduction
In recent years, investigation of the effects of the human humoral system on the skin microbiota and its biofilms has become an important field of research. Relatively large quantities of data have been collected regarding catecholamines [1] and neuropeptides [2]. Less is known about steroid hormones and their action on microbial cells, despite the elucidation of some aspects concerning the microbial metabolism of steroids [3,4] and changes in microbiota structure caused by steroids [5,6,7]. Additionally, some bacterial species have been studied for their reaction to estradiol. For instance, it was found that estradiol and other steroids can be quorum-sensing inhibitors in Pseudomonas aeruginosa, and analogues of receptors were found in P. aeruginosa and Agrobacterium tumefaciens [8]. Biofilms and virulence in Prevotella aurantica, a pathogenic microorganism from the human mouth, are stimulated by estradiol via the dipeptidyl peptidase IV regulation pathway [9]. Hence, it is important to study how human steroid hormones can affect other representatives of the human microbiota and its biofilms—biofilm being the universal form of bacterial life in most ecological niches, especially in cutaneous or mucosal microorganisms [10].
Because of significant interconnections between vaginal microbiota, its balance with skin microbiota in areas around external genitals, sex hormonal status, and genital tract homeostasis in women, estradiol as one of the key sex hormones in women is of special interest. Little is known of how estradiol actually affects the important aspects of female microbiota–lactobacilli. In addition to the lower vaginal tract, these bacteria inhabit some areas of the skin, such as areas around the female external genitals and even the hands [11,12,13]. Lactobacilli usually inhabit zones that are affected by female steroid hormones, particularly β-estradiol, including the internal and external female genitals. Female sex hormones are now considered mediators of the vaginal microbial community, they participate in the maintenance of vaginal eubiosis [14].
Lactobacillus paracasei (aka Lacticaseibacillus paracasei) is an important member of the gut microbial community and also can be a component of the normal vaginal microbiota [15,16]. It was shown that L. paracasei, as well as other lactobacilli, can influence the processes in human skin, such as modulating the inflammatory mechanisms in skin and the maintenance of skin health [17,18]. Moreover, the interconnection between the gut microbiome and the skin seems to be obvious nowadays [19].
Micrococcus luteus is another component of the human skin microbiota that is highly abundant on the skin, as well as colonizing hair follicles and stratum corneum [20]. Micrococcus is not a dominant part of the vaginal microbiota; nevertheless, it can interact with lactobacilli on the outer parts of the female genital tract and become a part of the potential lactobacilli-containing community, for instance, in abdominal skin [21]. Furthermore, M. luteus is a robust model for the investigation of bacterial interactions in biofilms between non-pathogenic bacteria. This is important, as probiotics interact not only with pathogenic microbes, and the balance between all parts of a community may be altered by the hormone administration.
A concentration of the hormone that was as universal as possible for women was required for the study. Therefore, for maximal unification of the potential data, it was decided to base the estradiol concentration to be used on the lower limit in the blood plasma of midluteal-phase women (0.22 nM) [22].
Hence, the aim of the study was to find the potential effect of estradiol, in concentrations close to that of blood plasma, on monospecies and mixed-species biofilms of skin commensals L. paracasei AK508 and M. luteus C01, which may be components of skin microbiota in women’s genital tracts.
2. Materials and Methods
2.1. Strains and Growth Conditions
Lactobacillus paracasei AK508 was obtained from the UNIQEM collection of FRS “Fundamentals of biotechnology” RAS (UNIQEM NO UQM41480). It originated from a kvass wort and was characterized by API test (see Supplementary Materials Table S1, Figure S1) by 16S rRNA sequencing, and was submitted to the NCBI database (accession ok559730). Despite the different origin, it is possible to use this strain as a model object to study interactions between the human commensals affected by estradiol. The bacterium was stored in sealed glass vials filled with liquid reinforced clostridial medium (RCM). The composition of RCM has been previously described [23]. Vials were stored at 4 °C. For the experiments, 50 µL of the culture was plated onto a Petri dish with 1.5% agar RCM and cultivated at 33 °C for aerobic colony growth. Subsequently, the colony was inoculated into screw-capped tubes filled with RCM and incubated at 33 °C for 72 h. The resulting culture was adjusted to an appropriate OD540 for the experiments.
Micrococcus luteus C01 (UNIQEM NO UQM41481) strain was isolated from the skin of a healthy volunteer, as described previously [24]. The bacteria were stored and the cultures were prepared as described previously [25]. For inoculation, an appropriate OD540 was adjusted for the overnight M. luteus C01 cultures.
2.2. Hormone Storage and Testing
β-estradiol (Merck, Darmstadt, Germany) of molecular weight 272.38 and purity ≥98% was diluted in 96% ethanol and stored at −18 °C. For the experiments, a series of stock dilutions was prepared in ethanol: 3.7 µM (10 ng/mL), 3.7 µM (1 ng/mL), 0.37 µM (0.1 ng/mL) and 3.7 × 10−8 nM (0.01 ng/mL). These dilutions were used to represent different concentrations of the hormone in blood plasma, and hence, to simulate the variations in physiological hormonal status in women. To simulate a concentration close to serum, the concentration of 3.7 × 10−8 M was used, while higher concentrations simulated different physiological situations with increased estradiol levels (e.g., during hormonal therapy [26]). Ethanol solutions of estradiol were inoculated into the liquid or 1.5% agar RCM in an appropriate volume before cultures to reach the baseline concentration of 2.2 × 10−10 M in the medium. The baseline concentration was conditionally designated as “physiological”. To evaluate the impact of ethanol on bacteria, additional control samples were prepared, using pure ethanol in the same concentration as for the samples with estradiol.
2.3. Biofilms on Polytetrafluoroethylene Cubes
Monospecies biofilms were grown on the surface of polytetrafluoroethylene (PTFE, Polyplast, Moscow, Russia) cubes as described previously [24,27,28]. For lactobacilli, aerobic and anaerobic cultivation were used, while for M. luteus, as for a strict aerobe, only aerobic conditions were appropriate. Briefly, in tubes with 21 cubes of a standard size of 4 × 4 × 4 mm and 3 mL of the RCM for aerobic growth, 18 µL of appropriate estradiol solutions and ethanol (the final ethanol concentration was 5.96‰) were added. Then, 50 µL of cultures (OD540 = 0.5) were inoculated and the tubes were incubated at 33 °C and 150 rpm for 72 h. For L. paracasei, the colony-forming unit- (CFU) count in the inoculum was an average of 5.9 × 107 CFU/mL, while that for M. luteus was 6.1 × 107 CFU/mL. For anaerobic growth of L. paracasei, 21 mL of RCM was added into the same tubes, as for aerobic growth, to fill them. The filled tubes were closed with screw caps to avoid additional aeration. Then, 12 µL of an appropriate estradiol solution was administered per tube. Then, 350 µL of the culture (OD540 = 0.5) was inoculated into the tubes at 33 °C for 72 h without shaking. After incubation, the OD540 values of the planktonic cultures were measured. Biofilms were stained with crystal violet (CV, Merck, Darmstadt, Germany) and the OD590 of the CV extracts was measured for total biomass evaluation, as described previously [25]. Briefly, after the planktonic culture removal, cubes with biofilms were washed twice gently with tap water and fixed by 3 mL of ethanol for 20 min. Then the ethanol was removed and the cubes with biofilms were stained with 3 mL of 0.5% CV water solution for 20 min. The CV was removed and the cubes were washed six times with tap water. Subsequently, the cubes were placed into new clean tubes, and CV was extracted from the biofilms in 3 mL of 96% ethanol per tube for 24 h. To avoid the evaporation of ethanol, tubes were covered with parafilm.
2.4. Growth Kinetics of Biofilms and Planktonic Cultures
Kinetics of biofilm growth were studied in two model systems, as described previously [29], for growth comparisons. In the first model, the biofilms were obtained from planktonic cultures without a stage of preliminary adhesion, while in the second model, cells were first adhered to the well bottoms. All cultures were prepared as described previously. In the first model system, 200 µL of RCM medium with or without added estradiol and ethanol was added into wells, after 3.3 µL of culture was added to the medium. In the second model, cultures were washed twice with sterile PS, then the OD540 was adjusted as described previously in sterile PS. Monospecies and mixed-species cell suspensions in PS were administered into the wells (200 µL) and left at room temperature for 2 h to ensure cell adhesion. Then, the cell suspension was removed by a pipette, attached cells were washed once with sterile PS to remove residual suspension, and 200 µL of medium, with or without the addition of active compounds, was inoculated into each of the wells. The plates in each model were incubated at 33.5 °C; with shaking for 72 h in a spectrophotometer XMark (Biorad, Hercule, CA, USA). The interval for OD540 measurement was 15 min.
Growth curves were obtained using the Microsoft Excel 2010 software (14.0.7268.5000 64x). In each experiment repeat, the specific growth rate was calculated as µ = (ln(OD2/OD1))/(t2 − t1), where OD1 and OD2 (and the corresponding t meanings) are at the beginning and end of the linear portion of the semi-logarithmic plot of the growth curve, respectively. Semi-logarithmic plots were constructed, and the linear portions were calculated using the SLOPE function in Excel. SLOPE allowed us to find the coefficient “a” of linear regression y = ax + b; thus, we identified the portion as “linear” when (i) there was minimal standard deviation of coefficient a; and (ii) a was maximal; that is, the curve was closer to vertical. Furthermore, the generation time t was calculated using the formula t = ln(2)/µ.
2.5. Confocal Laser Scanning Microscopy
To analyze mono- and mixed-species biofilms, cultures were adjusted to OD540 = 1 and 2, respectively. To obtain mixed-species biofilms, cultures of M. luteus and L. paracasei with OD540 = 2 were mixed with each other at a ratio of 1:1, in accordance with previous experiments. Biofilms were obtained in 24-well flat-bottom glass plates (Eppendorf, Hamburg, Germany). In each well, 1 mL of RCM was added, after appropriate volumes of estradiol solutions and ethanol were inoculated. Then, 50 μL of mono- or mixed-species cultures were inoculated and plates were incubated in a wet chamber at 33 °C for 72 h.
After incubation, FISH (R6G) was conducted for labeling L. paracasei in monospecies and mixed-species biofilms. M. luteus C01 monospecies biofilms were stained with the universal fluorescent dye SYTO9 Green (ThermoFisher, USA), as described previously [24]. Furthermore, the L. paracasei and mixed-species biofilms were also stained with SYTO9 Green, for the estimation of biomass ratio in mixed-species biofilms and hybridization ratio in L. paracasei. After staining, biofilms were covered with Prolong Gold Antifade Mounting Media (ThermoFisher, Waltham, MA, USA) at 2 drops per well. The plates were covered with foil and stored at 4 °C before the CLSM analysis.
CLSM was performed, as previously described [29], on an LSM510 confocal microscope (Carl Zeiss, Jena, Germany) equipped with a C-Apochromat 63×/1.2 W corr objective. SYTO9-labeled samples were excited using a 458 nm wavelength Ar+ laser. Fluorescence was detected at wavelengths longer than 505 nm using an BP475-525 long-pass filter. R6G-labeled probes were excited with a 543 nm wavelength HeNe laser, and fluorescence was detected at wavelengths longer than 560 nm using an LP560 long-pass filter. In both cases, the diameter of the confocal diaphragm corresponded to one Airy disk. The lateral resolution of the images was approximately 250 nm, and the optical section thickness was <0.9 μm. The scanning step in the Z-direction was 1 µm for routine scanning and 0.5 µm for more detailed 3D images. To increase the reliability of the results, microscopy was conducted in a blind manner.
The visualization of images was conducted using the Zen Blue (2011) Edition software (Carl Zeiss, Jena, Germany), and the numeric processing of files was performed using the ImageJ software (NIH, Bethesda, MD, USA) with the Comstat2 plug-in. The average biomass density (µm3/µm2) and the average thickness (µm) were calculated for each image (where possible) using a threshold of 15, the ratio of biomass in dual-species biofilms was evaluated, and the effect of ANP in immature and mature biofilms was measured.
2.6. Fluorescent In Situ Hybridization
The FISH procedure was performed for L. paracasei AK508 in accordance with [30], modified by [29]. After incubation, the planktonic suspension was removed, and biofilms were washed with 1 mL of sterile PS. Then, the biofilms were fixed with 1 mL of 96% ethanol for 20 min at room temperature (RT). Then, ethanol was removed, and wells were dried at RT. Wells with M. luteus C01 monospecies biofilms were stained with SYTO9 Green, as described previously [31]. Monospecies L. paracasei and mixed-species biofilms were stained with the probe 5′-GTATTAGCAYCTGTTTCCA-3′ specific to Lactobacillaceae [30]. The probe was labeled with R6G rhodamine at the 5′-end. After fixation, 150 μL of lysis buffer (1 mg/mL lysozyme; 2 μg/mL lysostaphin; 10 mM Tris-HCl, pH 7.4) was added to the wells. All of the components were obtained from Merck (Darmstadt, Germany). Cell-wall lysis was performed at 37 °C for 15 min. Cell membranes were permeabilized by exposing them for 3 min to increasing concentrations of ethanol (50%, 80%, and 100%) in pH 7.4 phosphate buffer at room temperature. After treatment, the wells with biofilms were properly dried at room temperature. Then, 200 µL of the hybridization buffer (20% formamide; 0.9 M NaCl; 20 mM tris-HCl, pH 8.0; 0.02% SDS) containing 1 ng/µL of the probe was administered to each well with lactobacilli. All the components of the buffer were purchased from Merck (Darmstadt, Germany). Hybridization was conducted in the wet chamber at 48 °C for 1 h. Then, the hybridization buffer was removed, and 200 µL of the washing buffer (0.9 M NaCl, 20 mM tris-HCl) was administered to the wells. After 15 min of washing at 50 °C in the wet chamber, the washing buffer was removed, and the samples were stained with SYTO9 and stored as described previously.
2.7. Biofilms on Glass Microfiber Filters
Mono- and mixed-species biofilms were grown on glass microfiber filters (GMFFs, Maidstone, UK) in liquid RCM in glass tubes, or on the surface of 1.5% agar RCM in Petri dishes. Due to their standard size and form, GMFFs are good for CFU counting, for MTT staining, and for aggregation analysis, because they actually allow collecting of the whole biofilm, instead of scrubbing it from microplate wells or from other non-destructible carriers. Additionally, these filters are relevant models of the parenchymatous skin derma structure [32,33]. In the liquid RCM, biofilms were grown in the presence of and in balance with planktonic culture; while on the Petri dishes, no planktonic cultures developed. In tubes, 6 mL of the RCM was added, and two filters were placed opposite one another vertically. After filtering, the hormone and ethanol were added to the appropriate tubes, and 100 µL of a monospecies or mixed-species culture was inoculated into the tube. For monospecies biofilms, OD540 cultures were used. For mixed-species biofilms, cultures of M. luteus C01 and L. paracasei AK508 with OD540 = 1.0 were mixed in 1:1 proportion, and then, 100 µL of mixed-species culture per tube was inoculated. Tubes were incubated at 33 °C and 150 rpm for 72 h. After incubation, one filter was stained with MTT for metabolic activity estimation, while another was dispersed for CFU counting and aggregation analysis.
On Petri dishes, 20 mL of the RCM agar with estradiol or ethanol was added. Six filters per plate were placed onto the agar, and 25 µL of a cell suspension per filter was inoculated onto the center of the filter surface. Two filters were used for monospecies M. luteus, L. paracasei, and dual-species biofilms. Monospecies biofilms were grown in parallel with mixed-species biofilms, for better interpretation of the microbial interactions.
2.8. CFU Counting and Cell Aggregation Analysis
The GMFFs with biofilms were placed into 10 mL of sterile physiological saline (PS) containing 0.9% of NaCl. From the tubes, the GMFFs were first washed gently in 50 mL of sterile PS to remove planktonic cells. GMFFs were dispersed as described previously [27], and a series of dilutions were prepared and plated on Petri dishes with the RCM agar for the CFU count. Petri dishes were incubated at 33 °C for 3–5 days in aerobic conditions. In parallel, a 10 μL drop of non-diluted suspension was fixed and stained with the CV on a glass slide for the following analysis of bacterial aggregation. The analysis was conducted using a Carl Zeiss Jena light microscope, at magnitude 1000× and with immersion oil (Sigma, St. Louis, MO, USA). Samples were checked visually and as completely as possible, after five representative photos were taken using the microscope-associated camera (Toup Photonics, Hangzhou, China). The number of single cells and cells in aggregates were counted, and the number of cells in mono- and mixed-species aggregates was also assessed.
2.9. Test of L. paracasei Antibacterial Properties
A test of the antibacterial properties of L. paracasei AK508 was conducted using a classical agar diffusion test, as described previously [34]. Briefly, in a Petri dish, 30 mL of agar RCM with or without active compounds was added, and 100 µL of L. paracasei culture (OD540 = 2) was plated with a spatula for lawn growth. Plates were incubated aerobically or anaerobically (for comparison) for 72 h at 33.5 °C. In addition to M. luteus C01, four test strains were used for the antibacterial properties test: Staphylococcus aureus 209P, Staphylococcus epidermidis ATCC14990, Pseudomonas aeruginosa PAO1, and Escherichia coli K12. All strains are human commensals, which may potentially interact with L. paracasei in a mucosal community. A total of 100 µL of OD540 = 0.1 night cultures in LB medium were inoculated in 20 mL of cooled RCM agar, then mixed and plated on Petri plates for deep-plate growth. Then, 10 mm diameter agar blocks with 72 h L. paracasei lawn were extracted and placed onto the agar surface with the test bacteria. Plates were incubated for 24 h at 33.5 °C. Finally, the inhibition zone diameters were measured.
Additionally, to assess the potential changes in fermentation processes in the presence of estradiol and ethanol, liquid cultures of L. paracasei AK 508 were prepared both aerobically and anaerobically. For anaerobic growth, 150 µL of an OD540 = 2 culture was placed into a screw-capped 22 mL glass tube filled with RCM with or without active compounds. The tubes were then incubated at 33.5 °C for 72 h. For aerobic growth, 350 µL of an OD540 = 2 culture was added into 250 mL conical flasks with 50 mL of RCM with or without active compounds. Flasks were incubated at 150 rpm and 33.5 °C for 72 h. Then, the cultures were centrifuged using an LMC-4200R at 3000× g for 30 min at room temperature, and the pH of supernatants was measured.
2.10. Statistics
All experiments were conducted at least in triplicate. Statistical analysis of the data was performed using the non-parametric Mann-Whitney U-test. All microbiological data plots were designed using the Microsoft Excel 2010 Software. Where appropriate, average relative values (control without addition of estradiol and/or ethanol was designated as 100%) and absolute values are plotted on the graphs, and the standard error of the mean is depicted as error bars. The p-values are also marked on data plots.
3. Results
3.1. Biofilms on PTFE Cubes
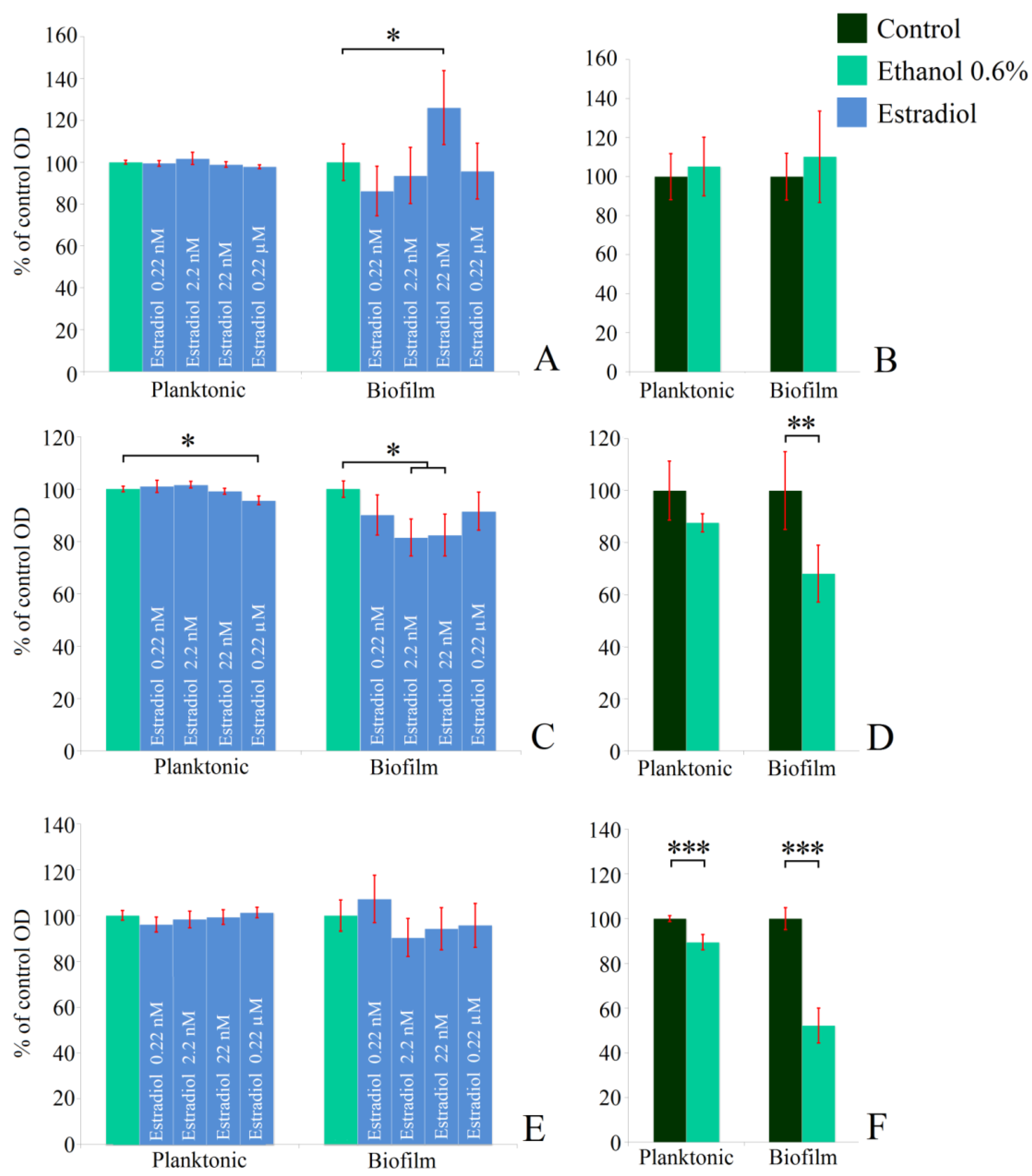
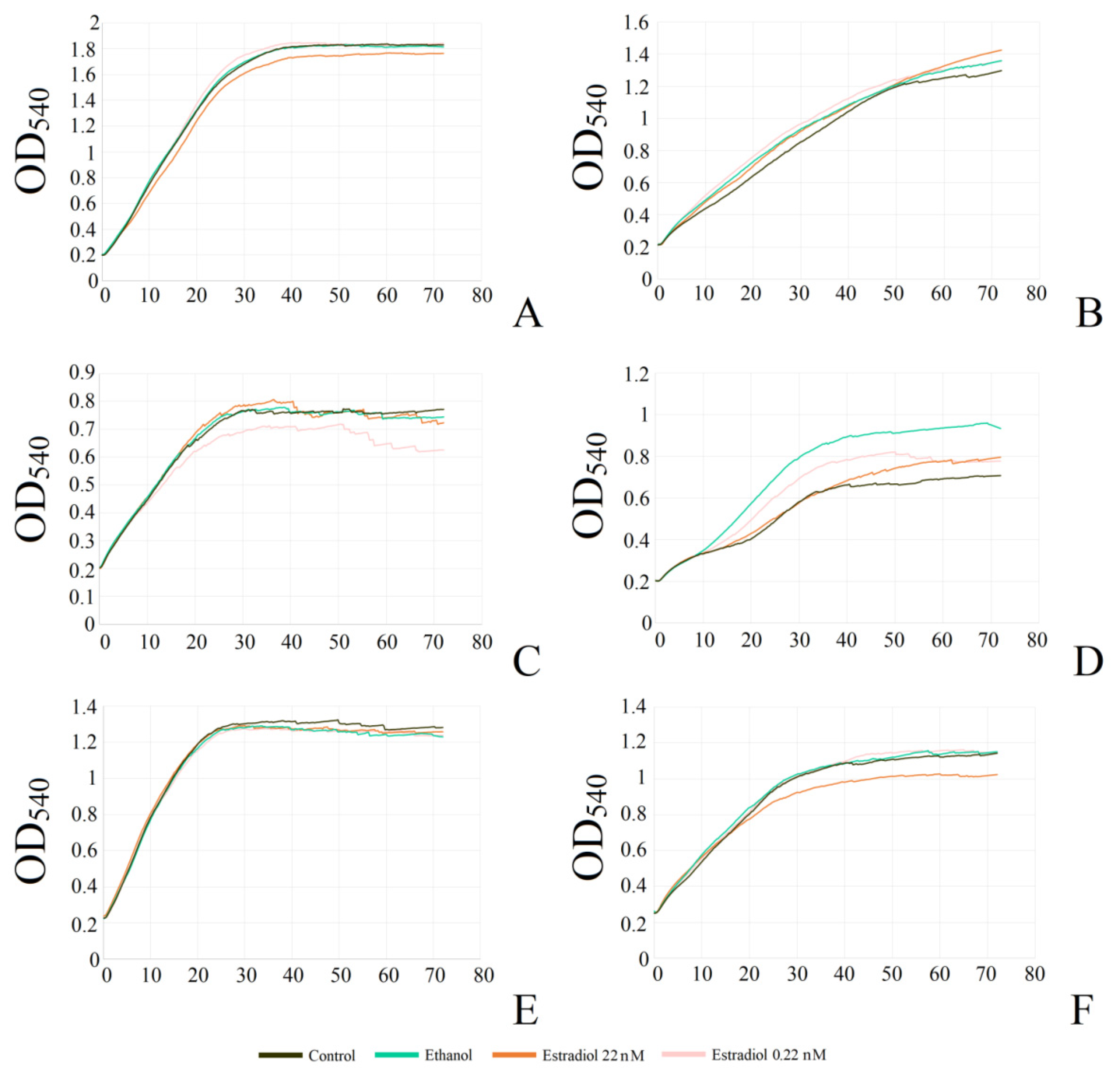
First, the monospecies planktonic culture growth in the presence of estradiol in comparison with vehicle controls (0.6% ethanol) was analyzed. Then ethanol samples were compared with controls without additions. M. luteus was incubated under aerobic conditions, while L. paracasei was tested both under aerobic and anerobic conditions. Neither 0.6% ethanol nor estradiol had effects on the planktonic cultures of M. luteus C01 (Figure 1A,B). The hormone reduced the OD of L. paracasei cultures by 5% at the highest concentration under aerobic cultivation, in comparison with 0.6% ethanol (Figure 1C). The effect was marginal, and it was difficult to determine any biological explanation; however, it was reproducible and statistically significant. Ethanol had also a marginal inhibitory effect (Figure 1D). Under anaerobic conditions, the effect of ethanol became statistically significant (Figure 1F), but no effect of the hormone was demonstrated.
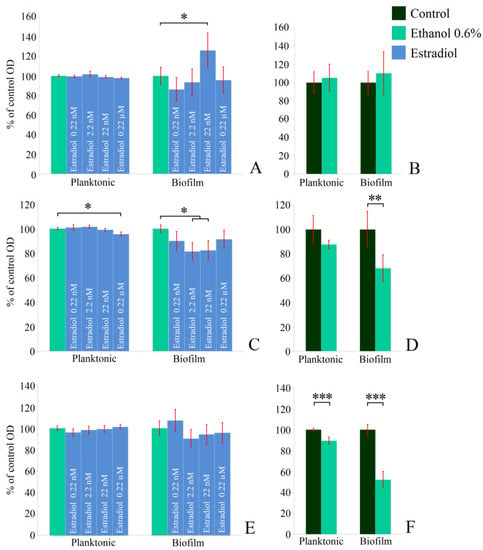
Figure 1.
Effect of β-estradiol on biofilms and planktonic cultures of M. luteus C01 (A,B) and L. paracasei AK508 (C,F) under aerobic (C,D) and anaerobic conditions (E,F). (A,C,E)—estradiol samples compared to 0.6% ethanol samples; (B,D,F)—ethanol 0.6% compared to controls without additions * denotes p ˂ 0.05; ** denotes p ˂ 0.01; *** denotes p ˂ 0.005.
Staining of monospecies biofilms with CV indicated the stimulatory effect of estradiol on M. luteus biofilms at a concentration of 22 nM (126 ± 17%), while the other concentrations did not affect the biofilms significantly (Figure 1A). Ethanol had no effect. Under aerobic conditions, L. paracasei biofilms were reduced marginally at concentrations of 0.22 nM and 0.22 µM; meanwhile, at 2.2 nM and 22 nM, the inhibition effect was statistically significant (81 ± 7% and 82 ± 8%, respectively). However, 0.6% of ethanol in the medium notably inhibited the biofilm growth of lactobacilli (68 ± 11%; Figure 1D). During anaerobic incubation, estradiol did not impact lactobacilli biofilms, however ethanol-caused biofilm inhibition became more pronounced (52 ± 8%, Figure 1F). Thus, the effect of estradiol seems to be condition- and concentration-dependent, while ethanol acted in the same way under both conditions. Meanwhile, under anaerobic cultivation, lactobacilli became even more sensible to the alcohol. Based on the results, 0.22 nM and 22 nM concentrations of estradiol were chosen for studying mixed-species biofilms of L. paracasei and M. luteus.
3.2. Growth Kinetics
In the study of the kinetics, the model systems were different in terms of the biofilm growth mode. The initial adhesion in the second model allowed us to study the biofilms of non-motile micro-organisms due to planktonic suspension formation by the adhered cells and biofilms, while in the first system, cells started to grow in suspension and form the biofilm from the planktonic phase. Hence, generally, in the first system, the studied planktonic cultures were presented, while in the second system, there were mostly biofilms. It was shown that monospecies and dual-species planktonic cultures in the system without preliminary adhesion were not significantly affected by estradiol and ethanol (Figure 2A, Table 1). In the second system, M. luteus monospecies biofilms were also not affected significantly by either ethanol or estradiol (Figure 2B, Table 1).
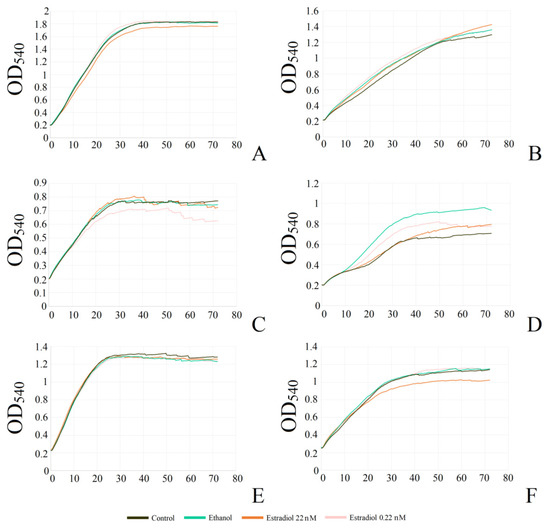
Figure 2.
Growth curves of mono- and mixed-species planktonic cultures and biofilms of M. luteus and L. paracasei. (A,C,E): model without preliminary adhesion; (B,D,F): model with preliminary adhesion; (A,B): monospecies M. luteus; (C,D): monospecies L. paracasei; (E,F): mixed-species culture.

Table 1.
Kinetic parameters of M. luteus and L. paracasei mono- and mixed-species planktonic cultures and biofilms.
In L. paracasei, the addition of ethanol did not strongly affect growth in the first system (Figure 2C); however, there was a tendency to reduction in growth rate and the maximal OD, especially in samples with 0.22 nM estradiol, which corresponds the results in the PTFE experiments. The addition of a higher concentration of estradiol did not decrease the growth of lactobacilli as well as the lower concentration. Biofilms, instead of planktonic cultures, were affected more strongly by both estradiol and ethanol. This effect seemed to appear after the logarithmic phase (Figure 2D). Here, estradiol did not affect the biofilm growth in the first few hours as significantly as later: after 8–9 h, there was observed an increase in biofilm growth in the presence of ethanol, and a decrease in the presence of estradiol, which resulted in lower OD540 as on the PTFE cubes (Table 1). Hence, it seems that estradiol facilitates the biofilm growth rate in initial stages, and reduces the final biomass and growth rate in later stages. However, ethanol acted as a growth stimulator in this system. Potentially, this could be explained by the altered surface for cell adhesion, or by a possible ethanol-caused shift between biofilm and planktonic suspension, which should be tested in future research.
The results of the mixed-species cultures and biofilms in our experiments appeared to fall between the results of M. luteus and L. paracasei monospecies cultures and biofilms (Figure 2E,F, Table 1). There was no effect from the tested compounds on the planktonic cultures; however, biofilm growth was altered in the presence of 22 nM estradiol. In the beginning, the growth rate was higher than in the presence of ethanol; later, the biofilm yield OD540 decreased. Thus, it could be suggested M. luteus has an advantage in the first stage of growth, with a subsequent balance shift to L. paracasei.
3.3. Confocal Microscopy of Mono- and Mixed-Species Biofilms
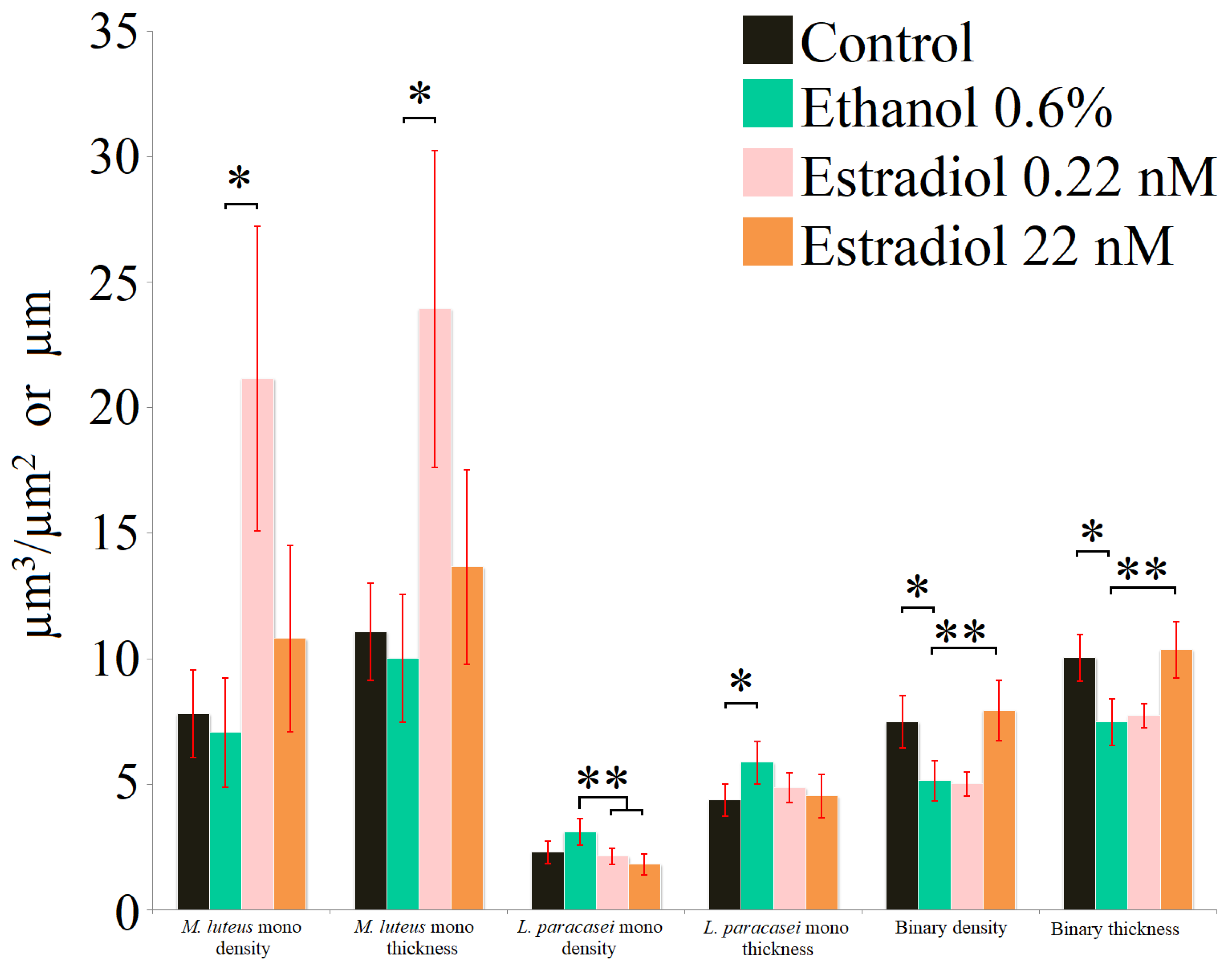
Analysis of the biofilms stained with SYTO9 (Figure 3, Table 2) showed the different results in comparison to the PTFE cube data. On the PTFE cubes and the 96-well plates, stimulation of M. luteus was observed at higher estradiol concentrations: in the 24-well plates with glass bottoms, the maximal stimulation was observed at 0.22 nM, while ethanol had no effect (similarly to the CV-staining results). The difference between the surface properties of hydrophilic glass and hydrophobic PTFE or polystyrene seem to be key for explaining these shifts. According to the results obtained using SYTO9, L. paracasei monospecies biofilms were reduced with estradiol (biofilm density), while ethanol increased the average thickness of biofilms, which seem to be caused by the hydrophilic glass as a carrier for the biofilms instead of the hydrophobic PTFE. Interestingly, FISH showed the low hybridization ratio of L. paracasei (Table 2), which may be explained by the biofilm matrix—an additional barrier for the FISH probe.
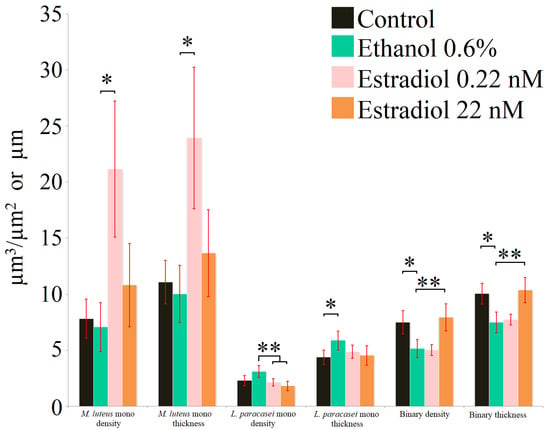
Figure 3.
Parameters of monospecies and binary biofilms stained with SYTO9 after 72 h of incubation. * denotes p ˂ 0.05; ** denotes p ˂ 0.01.
Table 2.
Parameters of mono- and mixed-species biofilms of M. luteus and L. paracasei in the presence of ethanol and estradiol after 72 h of incubation.
The mixed-species biofilms after 72 h mostly contained L. paracasei and very marginal M. luteus; thus, the contribution of Micrococcus biomass in binary biofilms after 72 h can be overlooked. Nevertheless, it is interesting that in this system, the biomass density and thickness of mixed-species biofilms increased in the presence of a higher concentration of estradiol in comparison with the ethanol control, while ethanol reduced those parameters significantly. It can be proposed that M. luteus has a positive impact on L. paracasei in binary systems; hence, the effects of estradiol may change significantly. Visualization of the biofilms is presented in the Supplementary Materials, Figure S2.
3.4. Biofilms on the Glass Microfiber Filters
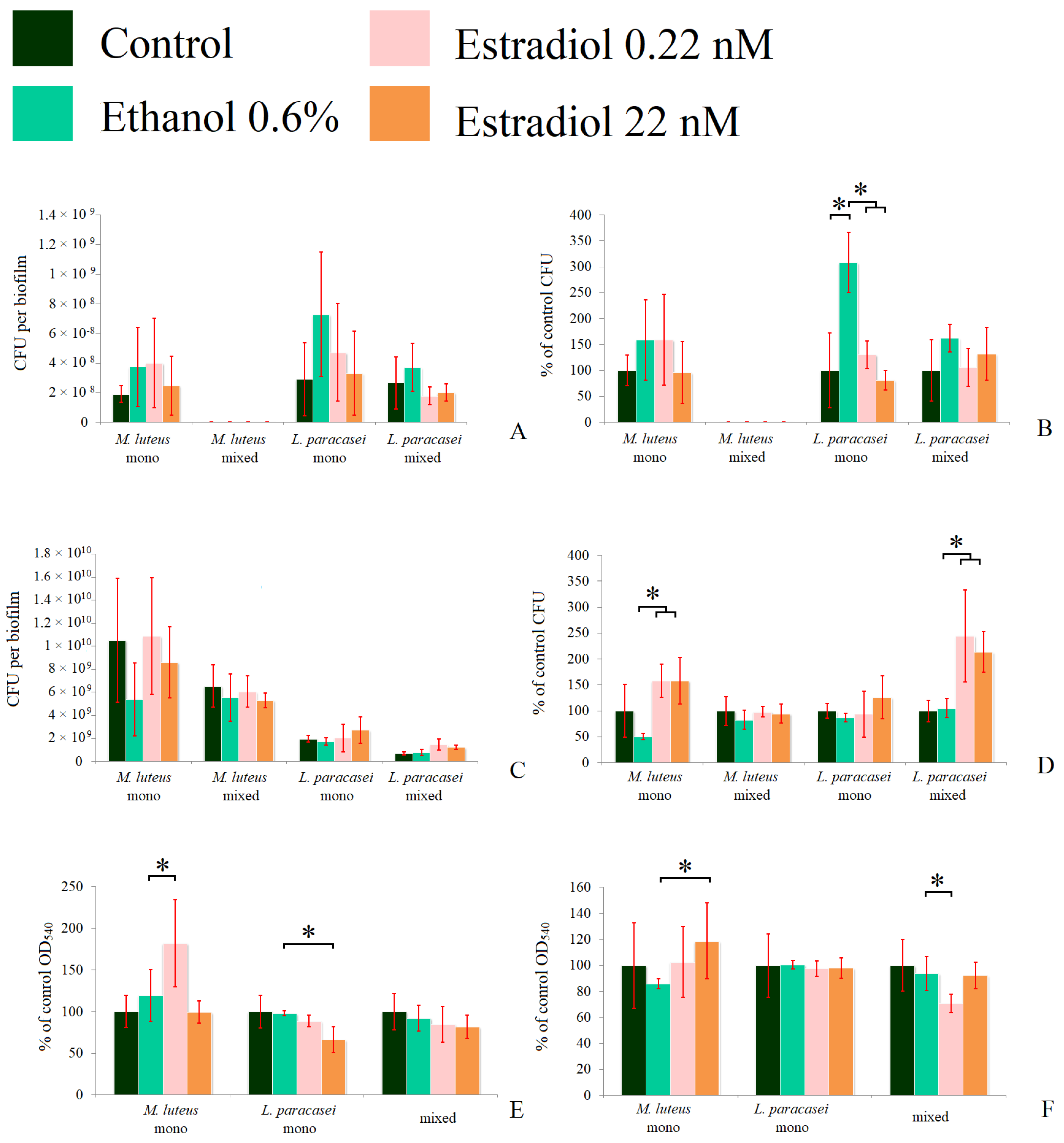
We analyzed the number of CFUs in 72 h planktonic cultures and biofilms of the tested bacteria, and found large statistical dispersions when counting the absolute values of CFUs. The dispersion was due to differences between experiments resulting in inappropriate general average values. Thus, we also calculated the relative values of the CFU counts. We used two model systems similar to those described in the kinetics measurements. In the first system, biofilms were grown in the presence of planktonic cultures; in each experiment, no M. luteus CFUs were found in the mixed communities (Figure 4A,B). A significant increase in CFUs in the presence of ethanol (308 ± 50%) was observed, in comparison to the control without additions (Figure 4B) in L. paracasei, which proves the hypothesis of the importance of surface properties. Estradiol eradicated this stimulatory effect of ethanol, in accordance with the results obtained in the CLSM section. A similar picture was observed for L. paracasei in the mixed-species biofilms, but the changes were not statistically significant. This can be potentially explained by the different surface structure, and thus, the different biofilm growth mode on the fibrous GMFFs and on the flat glass bottoms. Despite this, it was established that no visible M. luteus was observed in the mixed biofilms on the glass surfaces in the liquid medium. The metabolic activity of the biofilms was altered in the same way as the CFU count.
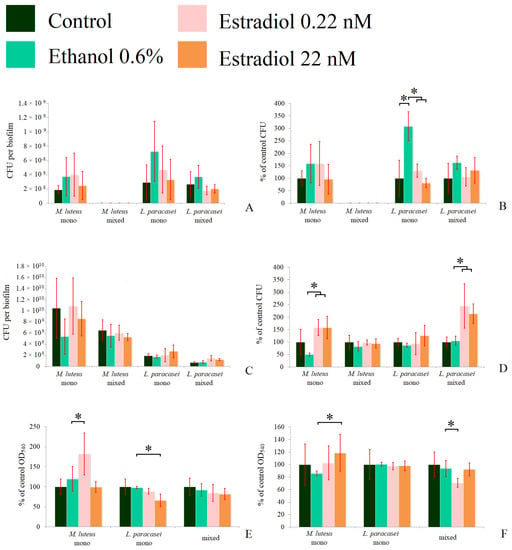
Figure 4.
The CFU counts in the absolute values (A,C), the relative values (B,D), and the formazan solutions OD540 (E,F) of mono- and mixed-species biofilms of M. luteus and L. paracasei after 72 h of incubation. (A,B,E): biofilms in the system in the tubes; (C,D,F): biofilms on the GMFFs on the surface of Petri plates. * denotes p ˂ 0.05.
3.5. Biofilms on the Glass Microfiber Filters
We analyzed the amount of CFUs in 72 h planktonic cultures and biofilms of the tested bacteria and found large statistical dispersions when counting the absolute values of CFUs. The dispersion was due to differences between experiments, resulting in inappropriate general average values. Thus, we also calculated the relative values of the CFU counts. We used two model systems similar to those described in the kinetics measurements. In the first system, biofilms were grown in the presence of planktonic cultures; in each experiment, no M. luteus CFUs were found in the mixed communities (Figure 4A,B). A significant increase in CFUs in the presence of ethanol (308 ± 50%) was observed, in comparison to the control without additions (Figure 4B) in L. paracasei, which approves the hypothesis of surface properties importance. Estradiol eradicated this stimulatory effect of ethanol, in accordance with the results obtained in the CLSM section. A similar picture was observed for L. paracasei in the mixed-species biofilms, but the changes were not statistically significant. This can be potentially explained by the different surface structure and, thus, the different biofilm growth mode on the fibrous GMFFs and on the flat glass bottoms. Despite this, it was established that no visible M. luteus part was in the mixed biofilms on the glass surfaces in the liquid medium. The metabolic activity of biofilms was altered in the same way as the CFUs count.
On the solid medium, the behavior of M. luteus was altered in comparison to the liquid medium system. Here, M. luteus was dominant over L. paracasei. The reason for this may be that without the liquid phase, M. luteus obtained the biofilm phenotype and started to form biofilms faster, while L. paracasei could not affect it by antibacterial activity, as in the liquid planktonic phase. In this system, estradiol significantly stimulated the growth of M. luteus, with CFU counts of up to 158 ± 31% and 158 ± 45% at the lower and higher concentrations, respectively (Figure 4D), which was also found in absolute values (Figure 4C). However, there was no stimulation of M. luteus CFUs in mixed-species biofilms. L. paracasei CFUs counts were increased significantly (more than 200% of control) in multi-species biofilms on Petri plates (Figure 4D) at both concentrations, but the absolute amounts were lower than the M. luteus counts. Thus, we can conclude that estradiol also has an effect in the initial adhesion stage. In the case of mixed-species biofilms, there is coherence with the CLSM results. Thus, we can propose that M. luteus in mixed-species biofilms potentially modulates the growth of L. paracasei, changing the effect of estradiol on this bacterium.
The metabolic activity of M. luteus monospecies biofilms was increased in the presence of a lower concentration of estradiol (Figure 4E) in the tube system. In L. paracasei biofilms, a significant decrease in metabolic activity (66 ± 15%) was observed at a concentration of 22 nM of estradiol. In mixed-species biofilms, the metabolic activity was decreased in the presence of 0.22 nM estradiol. This can be explained, on one hand, by the absence of M. luteus stimulation and, on the other hand, by changes in its metabolism in the presence of lactobacilli.
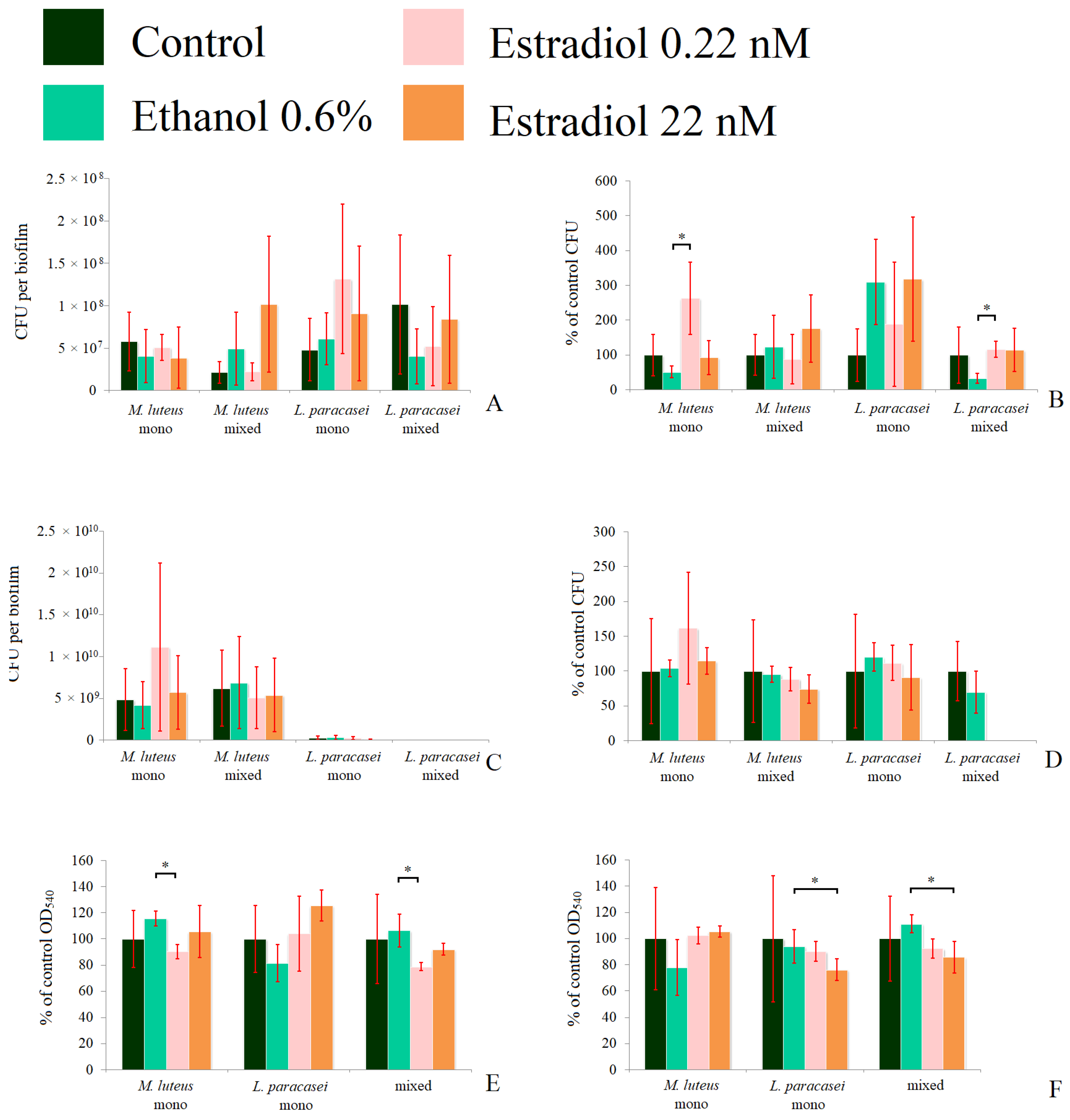
To clarify what happens in immature biofilms, we decided to test 24 h biofilms in the same experiments (Figure 5). In M. luteus monospecies biofilms, 0.22 nM estradiol increased the CFU count by 168 ± 37%, while ethanol decreased it by 60 ± 20% in the model system in tubes (Figure 5A,B). In mixed-species biofilms, M. luteus was stimulated by estradiol at both concentrations, especially at the higher concentration (334 ± 109%). In L. paracasei monospecies biofilms, ethanol stimulated the CFU amounts insignificantly, and estradiol slightly enhanced this effect, but no significant differences between estradiol and ethanol samples were found. Thus, most of the stimulation was based on the administration of ethanol to the medium. In the mixed-species biofilm, the tendency of biofilm inhibition was observed in the presence of ethanol and 0.22 nM of estradiol, with no effect of higher estradiol concentration, according closely with the CLSM results. In the second model system on the Petri dishes, M. luteus was stimulated by estradiol at both concentrations (224 ± 67% and 148 ± 22% at the lower and the higher concentrations, respectively; see Figure 5D). No effect was observed on M. luteus in the mixed-species biofilms. L. paracasei was not found in mixed-species biofilms; this can be explained by the competitive advantage of M. luteus inside the biofilm (importantly, not in the planktonic phase). The metabolic activity test showed no alteration in the presence of active compounds. Thus, we can propose that neither estradiol nor ethanol significantly affect the activity of at least NDH-dehydrogenases in both bacteria.
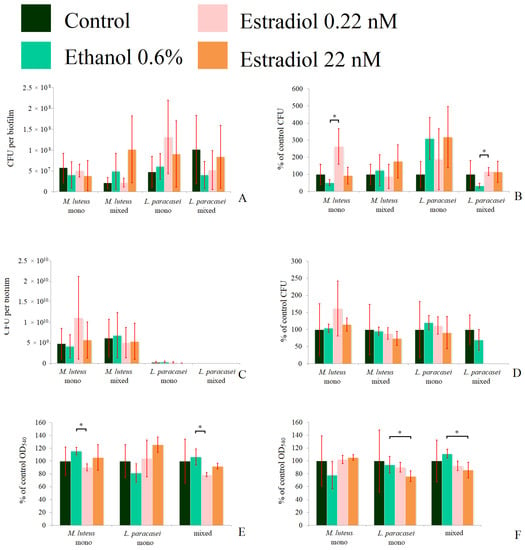
Figure 5.
The CFU counts in the absolute values (A,C), the relative values (B,D), and the formazan solutions OD540 (E,F) of mono- and mixed-species biofilms of M. luteus and L. paracasei after 24 h of incubation. (A,B,E): biofilms in the system in tubes; (C,D,F): biofilms on the GMFFs on the surface of Petri plates. * denotes p ˂ 0.05.
Hence, we can conclude that in the case of L. paracasei AK508, the hydrophobicity of a surface seems to play a key role in estradiol and ethanol action on biofilm growth; however, micrococci were more consistent and did not depend on the surface type. Hence, the differences in cell surfaces seem to be important factors.
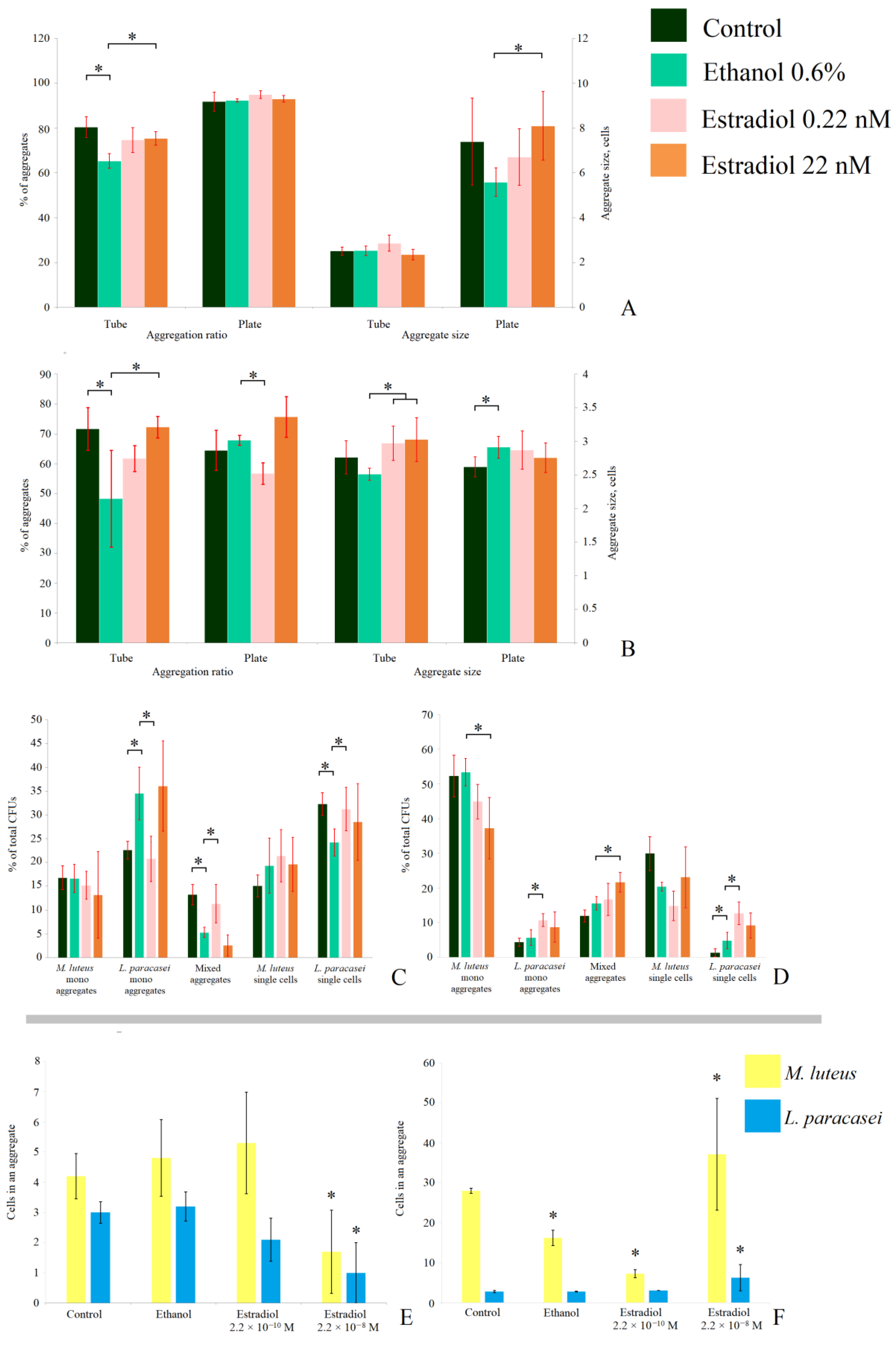
3.6. Analysis of Cell Aggregation in Biofilm
To study how the CFU counts were relevant to the actual cell biomass in biofilms in depth, we additionally analyzed the aggregates of M. luteus and L. paracasei in the 24 h biofilms. The visualization of the samples is depicted in the Supplementary Data, Figure S3. In M. luteus, biofilms in the liquid medium in tubes, 80 ± 5% (Figure 6A) of all analyzed cells in the samples were in aggregates, while the average aggregate size was 2.5. This means that most cells were actually in pairs. The addition of active compounds did not lead to a change in the aggregate size, but ethanol decreased the number of aggregated cells to 65 ± 3%. Estradiol at both lower and higher concentrations increased the aggregation ratio in comparison with ethanol samples, but the effect was statistically significant only at higher concentrations (75 ± 3%). On the filters on the solid medium, the aggregation was higher than in the liquid medium, which can be explained by more intensive biofilm growth (about 90% in each sample), and the aggregate size was 7.3 ± 2 cells per aggregate. Ethanol insignificantly decreased the size, but an addition of 22 nM of estradiol significantly reduced the effect of ethanol, resulting in larger cell aggregates (5.6 ± 0.6 and 8.1 ± 1.5 cells per aggregate, respectively). Thus, ethanol in both model systems reduced the aggregation, while estradiol increased it. In combination with the results of CFU counting, it can be carefully suggested that the hormone increased the growth of cell biomass in M. luteus, while ethanol suppressed cell proliferation in monospecies biofilms.
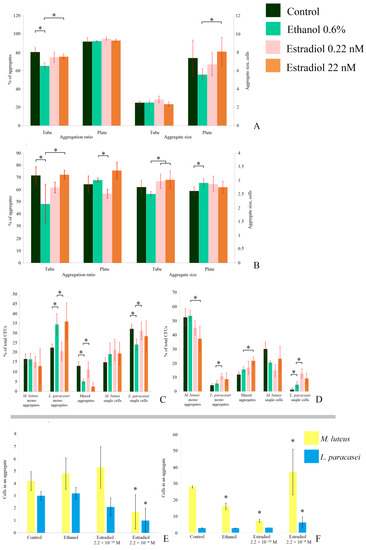
Figure 6.
Aggregation analysis of 24 h monospecies and mixed-species biofilms of M. luteus and L. paracasei on the GMFFs in different model systems: (A). M. luteus monospecies biofilms; (B). L. paracasei monospecies biofilms; (C). aggregation ratio in mixed-species biofilms in the liquid medium; (D). aggregation ratio in mixed-species biofilms on the solid medium; (E). aggregate size in mixed-species biofilms in the liquid medium; and (F). aggregate size in mixed-species biofilms on the solid medium. * denotes p ˂ 0.05.
In L. paracasei monospecies biofilms (Figure 6B), the aggregation ratio in the liquid RCM was close to that of M. luteus. Furthermore, the effects of active compounds in the liquid medium were the same as for M. luteus: ethanol decreased the aggregation ratio (73 ± 13% and 46 ± 16% in the control and ethanol samples, respectively). Estradiol increased the aggregation ratio under both concentrations, with a statistically significant difference in comparison with ethanol at the higher concentration (73 ± 4%). Moreover, the aggregate size was larger in the presence of estradiol, in comparison to the ethanol samples: 2.9 ± 0.3, 3 ± 0.4, and 2.5 ± 0.1 cells per aggregate in the samples with 0.22 nM, 22 nM, and in the presence of ethanol, respectively. Thus, interestingly, estradiol also increased the proliferation of L. paracasei in 24 h biofilms, while ethanol reduced this process. This may also be explained by the potential changes in cell surface properties: in the presence of active compounds, cell adhesion could be altered. Thus, the amount of cell aggregates differed in different samples. Additionally, increased coaggregation could be a reason for the marginal OD540 decrease in L. paracasei shown in Figure 1.
In mixed-species biofilms, the effect of the active compounds was more complex (Figure 6C–F). We attempted to analyze the origins of the CFUs on the Petri plates after biofilm dispersion. We found that, despite there being no colonies of M. luteus on Petri dishes after plating of the biomass of biofilms from the liquid medium, 12%–16% of M. luteus monospecies cell aggregates and 14%–22% of M. luteus single cells on the CV-stained glass slides could be potential CFUs. The percentage depended on the presence of active compounds (Figure 6C), and was not altered significantly. Ethanol increased the number of monospecies aggregates of L. paracasei (from 22 ± 3% to 35 ± 5%) and decreased the number of single cells (from 32 ± 2 to 24 ± 3%), while 0.22 nM had an opposite effect: decreased aggregation to 21 ± 5%, and increased single cell amount to 31 ± 5%. The higher concentration of estradiol had no effect in comparison to ethanol samples (Figure 6C). The counts of mixed-species aggregates and their composition (Figure 6E) were also altered in the presence of estradiol and ethanol. Estradiol at the concentration 0.22 nM increased the number of mixed-species aggregates (from 5 ± 1% in ethanol samples to 11 ± 4%), but did not affect the aggregate size. The concentration of 22 nM reduced both the ratio and size of the aggregates (Figure 6E). Here, the average composition of an aggregate was 1.7 cells of M. luteus and 1 cell of L. paracasei; while in ethanol samples, it was 4.8 M. luteus cells and 3.2 L. paracasei cells. Thus, the action of estradiol in the case of cell aggregation also depends on concentration.
In the solid medium, the effects of active compounds on aggregation in mixed-species biofilms varied (Figure 6D,F). First, we found that L. paracasei was not totally eradicated—about 8–20% of all potential CFUs were L. paracasei single cells or aggregates. The absence of L. paracasei colonies should be a consequence of the potential dormancy of lactobacilli, due to unfavorable conditions in a mixed population. Moreover, the numbers of monospecies aggregates of L. paracasei and mixed-species aggregates were increased in the presence of the hormone, while monospecies M. luteus aggregates were reduced in number. Aggregates were several times larger than in the tubes (Figure 6F), and at different concentrations, estradiol had a controversial effect: at the concentration of 0.22 nM, it decreased the M. luteus portion in mixed-species aggregates (16.2 ± 2.8 M. luteus cells in the ethanol samples and 7.3 ± 1 in the presence of the hormone). At the concentration of 22 nM, the hormone increased the number of cells—37 ± 14 cells of M. luteus and 6.2 ± 3.3 cells of L. paracasei—while in the ethanol samples there were 2.8 ± 0.1 lactobacilli cells per aggregate. Thus, it can be suggested that the balance of M. luteus colonies in mixed-species biofilms (no effect of estradiol; Figure 5) was achieved by shifting between single cells, monospecies, and mixed-species aggregates, which were caused by active compounds. Furthermore, some of the M. luteus colonies seem to have originated from mixed-species aggregates. Thus, the hormone acts as a mediator for cell-to-cell aggregation, and its action depends on the concentration.
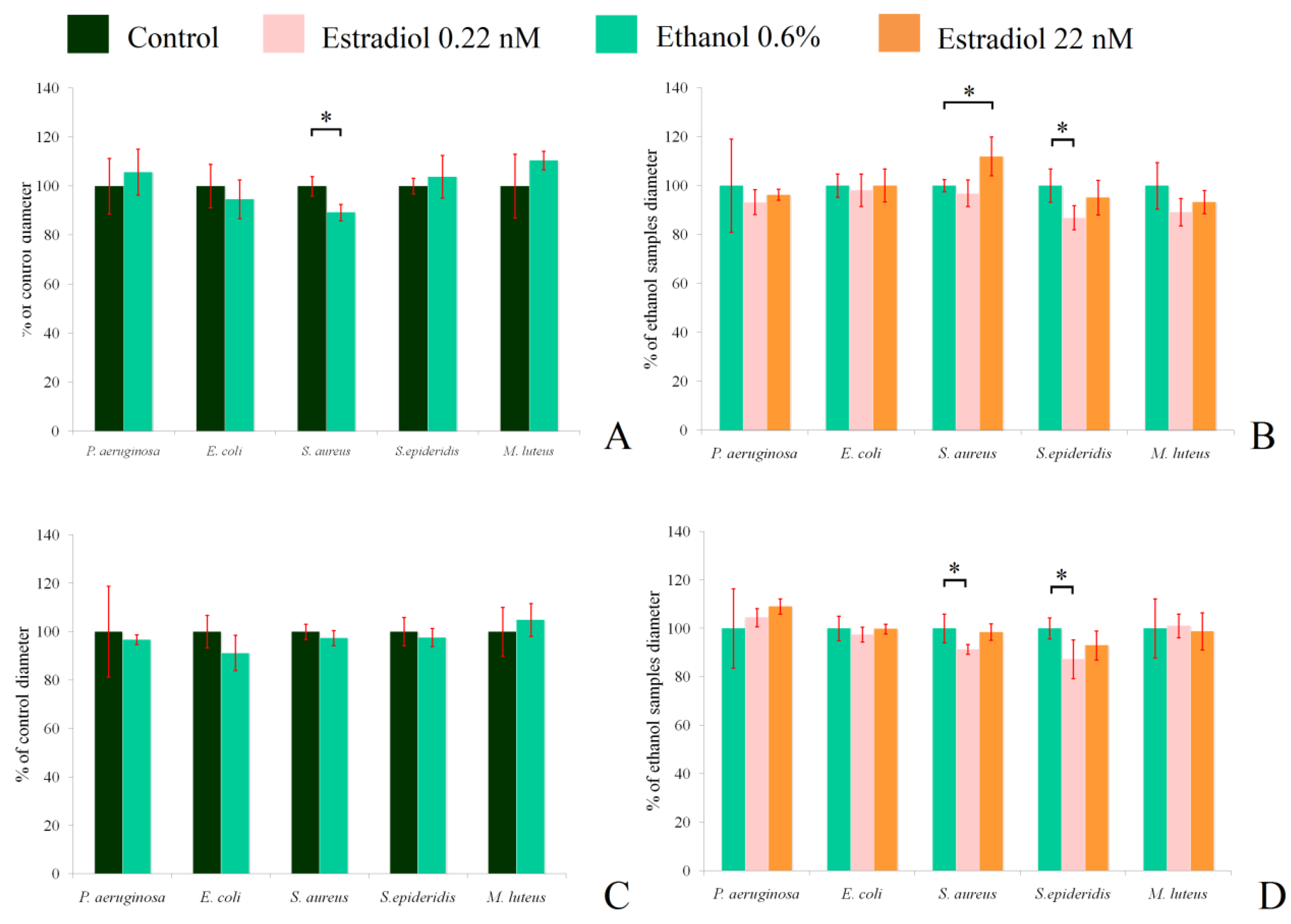
3.7. Antibacterial Properties Test
Inhibition zone measurements showed that the hormone insignificantly decreased the antibacterial activity of L. paracasei against M. luteus when grown under aerobic conditions (Figure 7B), and did not have any effect under anaerobic conditions (Figure 7D). Ethanol had a statistically significant inhibitory effect on the activity against S. aureus 209P under aerobic conditions (89.2 ± 3.4% of the control; Figure 7A), while the higher concentration of estradiol increased the activity (112 ± 8%, in comparison with ethanol samples). The antimicrobial action of lactobacilli against S. epidermidis under aerobic conditions was lower in the presence of 0.22 nM estradiol (86.9 ± 4.9%; Figure 7B). Under anaerobic conditions, ethanol did not impact the antimicrobial properties of lactobacilli, but the effect of estradiol was altered in the case of S. aureus. Here, the lower concentration of estradiol decreased the activity to 91 ± 2%, in comparison to ethanol, while the hormone did not impact lactobacilli at the higher concentration. Against S. epidermidis, the effect was similar under both aerobic and anaerobic conditions. Measurement of the medium pH showed no effect of estradiol or ethanol on the final results (the pH varied from 3.8 to 4.2, depending on the repeat); thus, estradiol potentially shifts the synthesis of special antimicrobial compounds in L. paracasei, depending on the growth conditions. The changes in mixed-species biofilms with M. luteus seem not to be related to the alterations of antibacterial properties in the presence of the hormone, despite the statistically insignificant decrease in antibacterial activity of L. paracasei on M. luteus. The photos of the inhibition zones are in the Supplementary Materials, Figure S4.
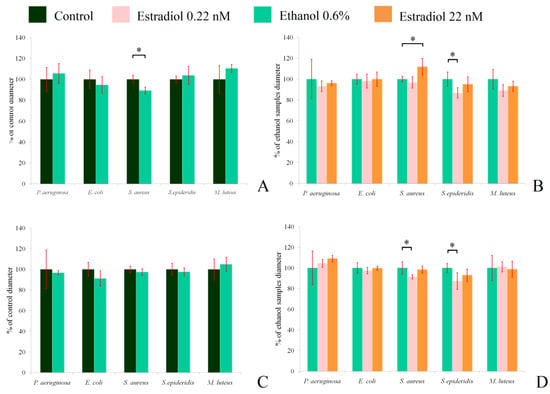
Figure 7.
Antibacterial activity of L. paracasei against test strains under aerobic (A,B) and anaerobic (C,D) conditions. (A,C): the impact of ethanol in comparison with the control without additions; (C,D): impact of the hormone in comparison with ethanol. * denotes p ˂ 0.05.
4. Discussion
It was found earlier that the action of estradiol is dose-dependent and strain-specific. The hormone alters the surface properties of cells, biosurfactant production, cell adhesion, and has a small effect on biofilm formation in L. crispatus [35,36]. In the present study, dual- species biofilms of M. luteus and L. paracasei were investigated for the first time. As shown in previous works of Clabaut et al., estradiol acted as an effector on lactobacilli at a concentration of 0.1 nM [35,36]. In our case, we also found that estradiol acted in dose-dependent manner, and the so-called “physiological” blood-plasma concentration had an effect on both bacteria in different systems. However, the concentration of hormones in tissues is not a trivial question, and in the case of human skin microbiota studies it becomes especially important, due to the need for correct experimental design. In the case of estradiol, the easiest way was to use the average concentration of estradiol in serum. According to the review of Carmina and Lobo [22], the estradiol concentration in blood plasma in midluteal-phase women is 0.22–0.73 nM. However, the correlation between estradiol concentration in tissues and serum seems to be marginal.
In the present study, it was shown that estradiol is able to regulate the dual-species community of skin and mucosal bacteria comprising M. luteus and L. paracasei. The action of the hormone depends on the cultivation conditions, the carrier surface type, and the concentration, which may be key for its regulatory function. Estradiol has a complex effect on both bacteria, as has been previously shown for other hormones [1,27,29]. Estradiol may alter some deep mechanisms in cells, with effects that should be investigated in the future: (i) regulation of cell growth and duplication in M. luteus and L. paracasei; (ii) cell surface changes and consequent adherence shifts depending on the carrier type; (iii) changes in extracellular matrix. Potentially, the matrix of L. paracasei becomes less dense, thus on flat surfaces, such as PTFE cubes and microtiter plates with glass bottoms, monospecies biofilms of lactobacilli become less stable, which results in decreased biomass, while on rough and fibrous filter surfaces, cells can better attach and maintain biomass.
M. luteus C01 is stimulated by estradiol. Its behavior and presence in dual-species biofilms shifts the estradiol effect on L. paracasei, as shown in the CLSM experiments. Despite lactobacilli dominating over the micrococci in that system, the presence of micrococci seemed to change the structure of biofilms, resulting in a shift from biofilm reduction to stimulation in the presence of estradiol at higher concentrations, while ethanol decreased both biofilm density and thickness. Interestingly, in dual-species communities, L. paracasei, CFU numbers and aggregate size were stimulated as well as in monospecies biofilms. M. luteus growth in dual-species communities’ cell growth became suppressed, despite the stimulation of CFUs and aggregates in monospecies biofilms. To keep in mind the effects of different bacteria on each other within a community is crucially important for fundamental studies, but also for new antibacterial drug development, due to the fact that a target bacterium can be protected by its counterparts [37].
Additionally, micrococci were shown to be promotors of other bacteria growth in some cases. Gene rpf encodes growth factors stimulating other bacteria [38,39]. This can be an explanation of the M. luteus-caused stimulation of lactobacilli (this must be verified in future). Despite the fact that the hormone generally increased aggregation in monospecies biofilms, M. luteus in the community lost most of its own aggregation ability and growth in stimulating the lactobacillus. On the other hand, as a well-known producer of bacteriocins, L. paracasei inhibits other bacteria. Thus, it is important that the dramatically increased resistance of M. luteus to antibacterial compounds produced by L. paracasei is caused by the rapid switch to the biofilm growth mode in systems where cells adhere faster to surfaces. In this case, estradiol shifts the balance between M. luteus C01 and L. paracasei AK508 to weaken the lactobacilli.
Interestingly, estradiol shifted the antibacterial activity of L. paracasei cells against different model microorganisms, especially against staphylococci. This effect was also dependent on the concentration and cultivation conditions. According to the NCBI database, L. paracasei is able to synthesize at least two bacteriocins of higher (57 aa) and lower (39 aa) mass, and it could be suggested that estradiol causes alterations in the synthesis of the different bacteriocins, due to its controversial effect on staphylococci. It could also be suggested that the alterations are connected with bacteriocinsdue to the lack of alterations in medium pH, and as a result, in the fermentation and medium acidification processes in the presence of the hormone.
Another interesting observation is connected with ethanol. At a concentration of 0.6% vol., it is a strong regulator of the microbial community, and ethanol acts—in most cases—as an antagonist for estradiol. Ethanol is an appropriate solvent for substances with low water solubility, and the use of vehicle control with the same ethanol (or other solvents) concentration is a common practice [35,36,40]. Vidaillac et al. [40] found interesting effects of steroids (including estradiol) in P. aeruginosa membranes, and the final concentration of ethanol in TSB was 1%. In the work of Plotkin & Konakieva [41] the highest vehicle control concentration was 0.5%. However, a vehicle control does not appear to be a magic pill for clarifying and distinguishing the effects of vehicles themselves, because some studies have revealed the influence of very low concentrations (0.1% v/v) of ethanol [42] on the behavior of bacteria. However, we can speculate that hypothetically, in some microniches in the genital tract, there may occur microenvironments where heterofermentative lactobacilli or other ethanol producers might concentrate and locally increase the ethanol concentration. For instance, L. fermentum., L. reuteri, and L. crispatus are potential producers of ethanol [43,44,45,46]. Hypothetically, Candida albicans as a fermentative microorganism [47] in the case of candidiasis may increase the ethanol concentration locally.
In our case, the effect of estradiol is minimally overlapped by ethanol, because of the similarity of ethanol effects in different systems. In the case of M. luteus, there was no effect of ethanol observed in CLSM and PTFE. Both bacteria altered their aggregation in the presence of active compounds on the GMFFs, and this proves the hypothesis regarding the surface-property changes.
5. Conclusions
We was established that estradiol, at concentrations of 0.22nM or 22 nM, has a regulatory effect on mono- and mixed-species biofilms of M. luteus C01 and L. paracasei AK508. The hormone shifts aggregation processes and biofilm biomass density in both bacteria. The effect of ethanol as a vehicle depended on surface hydrophobicity, while estradiol had generally the same effects on monospecies biofilms in different systems. This suggests, on the one hand, that the distinguishing of ethanol and estradiol effects was successful, and on the other hand, the potentially significant effect of ethanol and estradiol on cell surface properties. The presence of micrococci in the mixed community stimulated lactobacilli and switched the hormone-caused inhibition to stimulation. The data obtained, together with the previous studies, show the complexity of interactions between human commensal microbiota and the humoral systems. We can suggest that estradiol acts as a regulator for human microbiota communities, but the deep mechanisms of such an action should be investigated in future studies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/coatings12040436/s1, Figure S1: The API test of L. paracasei AK508. The list of compounds is presented in the Table S1; Figure S2: The CLSM images of monospecies (A–L) and mixed-species (M–T) biofilms of M. luteus and L. paracasei. A–D—monospecies biofilms of M. luteus stained with SYTO9, E–L—monospecies biofilms of L. paracasei stained with SYTO9 (E–H) and the FISH probe with R6G (I–L). M–T—mixed-species biofilms stained with SYTO9 (M–P) and the FISH probe with R6G (Q–T). A,E,I,M,Q—control samples, B,F,J,N,R—ethanol samples, C,G,K,O,S—estradiol 2.2 × 10−10 M; D,H,L,P,T—2.2 × 10−8 M; Figure S3: Representative images of the samples of monospecies M. luteus biofilms (A–H), monospecies L. paracasei biofilms (I–P) and mixed-species biofilms (Q–X) for aggregation assessment in the system with tubes (A–D, I–L, Q–T) and on the solid medium in Petri dishes (E–H, M–P, U–X). A,E,I,M,Q,U—control samples, B,F,J,N,R,V—ethanol samples, C,G,K,O,S,W—estradiol 2.2 × 10−10 M, D,H,L,P,T,X—estradiol 2.2 × 10−8 M; Figure S4: The images of inhibition zones around the agar blocks with L. paracasei grown areobiaclly (A,C,E,G,I) and anaepobically (B,D,F,H,J). A,B—M. luteus, C,D—P. aeruginosa PAO1, E,F—Escherichia coli K12, G,H—Staphylococcus aureus 209P, I,J—Staphylococcus epidermidis ATCC14990. 1—control, 2—ethanol, 3—estradiol 2.2 × 10−8 M, 4—estradiol 2.2 × 10−10 M; Table S1: API test of L. paracasei AK508. Red label is for differences between strain AK508 and type strain R094.
Author Contributions
Conceptualization, A.V.G. and V.K.P.; methodology, A.A.K., T.V.S. and A.V.G.; validation, A.A.K., T.V.S., M.A.O., S.V.M., O.V.G., N.D.D., A.V.F., E.A.B., V.K.P. and A.V.G.; formal analysis, A.A.K., T.V.S., S.V.M. and A.V.G.; investigation, A.A.K., T.V.S., M.A.O., O.V.G., S.V.M., N.D.D., T.A.C., M.V.Z. and A.V.G.; resources, A.V.G.; data curation, A.A.K., T.V.S., M.A.O. and A.V.G.; writing—original draft preparation, A.A.K., T.V.S. and A.V.G.; writing—review and editing, S.V.M., O.V.G., N.D.D., A.V.F., E.A.B., T.A.C., M.V.Z. and V.K.P.; visualization, A.A.K.; supervision, V.K.P. and A.V.G.; project administration, A.V.G.; funding acquisition, A.V.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by RSCF project №19-74-10071. The work of V.K.P. was supported by the Ministry of Science and Higher education of Russian Federation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to colleagues of the Laboratory of Viability of Microorganisms, and especially to Yu. A. Nikolayev, for valuable advice and help in the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gannesen, A.V.; Schelkunov, M.I.; Geras’kina, O.V.; Makarova, N.E.; Sukhacheva, M.V.; Danilova, N.D.; Ovcharova, M.A.; Mart’yanov, S.V.; Pankratov, T.A.; Muzychenko, D.S.; et al. Epinephrine affects gene expression levels and has a complex effect on biofilm formation in Micrococcus luteus strain C01 isolated from human skin. Biofilm, 2021; in press. [Google Scholar]
- Racine, P.J.; Janvier, X.; Clabaut, M.; Catovic, C.; Souak, D.; Boukerb, A.M.; Groboillot, A.; Konto-Ghiorghi, Y.; Duclairoir-Poc, C.; Lesouhaitier, O.; et al. Dialog between skin and its microbiota: Emergence of “Cutaneous Bacterial Endocrinology”. Exp. Dermatol. 2020, 29, 790–800. [Google Scholar] [PubMed]
- Donova, M.V.; Egorova, O. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef] [PubMed]
- Insenser, M.; Murri, M.; Del Campo, R.; Martinez-Garcia, M.A.; Fernandez-Duran, E.; Escobar-Morreale, H.F. Gut mi-crobiota and the polycystic ovary syndrome: Influence of sex, sex hormones, and obesity. J. Clin. Endocrinol. Metab. 2018, 103, 2552–2562. [Google Scholar]
- Pigrau, M.; Rodiño-Janeiro, B.K.; Bedmar, M.T.C.; Lobo, B.; Vicario, M.; Santos, J.; Cotoner, C.A. The joint power of sex and stress to modulate brain-gut-microbiota axis and intestinal barrier homeostasis: Implications for irritable bowel syndrome. Neurogastroenterol. Motil. 2015, 28, 463–486. [Google Scholar] [CrossRef]
- Haro, C.; Rangel-Zuñiga, O.A.; Alcala-Diaz, J.F.; Delgado, F.G.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortes, J.; Tena-Sempere, M.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE 2016, 11, e0154090. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sun, Z.; Jiang, S.; Bai, X.; Ma, C.; Peng, Q.; Zhang, H. Probiotic Bifidobacterium lactis V9 regulates the secre-tion of sex hormones in polycystic ovary syndrome patients through the gut-brain axis. Msystems 2019, 4, e00017-19. [Google Scholar]
- Beury-Cirou, A.; Tannières, M.; Minard, C.; Soulère, L.; Rasamiravaka, T.; Dodd, R.H.; Queneau, Y.; Dessaux, Y.; Guillou, C.; Vandeputte, O.M.; et al. At a Supra-Physiological Concentration, Human Sexual Hormones Act as Quorum-Sensing Inhibitors. PLoS ONE 2013, 8, e83564. [Google Scholar] [CrossRef]
- Fteita, D.; Musrati, A.A.; Könönen, E.; Ma, X.; Gürsoy, M.; Peurla, M.; Söderling, E.; Sintim, H.O.; Gürsoy, U.K. Dipeptidyl peptidase IV and quorum sensing signaling in biofilm-related virulence of Prevotella aurantiaca. Anaerobe 2017, 48, 152–159. [Google Scholar]
- Zommiti, M.; Feuilloley, M.G.J.; Connil, N. Update of Probiotics in Human World: A Nonstop Source of Benefactions till the End of Time. Microorganisms 2020, 8, 1907. [Google Scholar] [CrossRef]
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb. Ecol. 2018, 76, 113–120. [Google Scholar] [PubMed]
- Egert, M.; Simmering, R. The Microbiota of the Human Skin. In Microbiota of the Human Body. Advances in Experimental Medicine and Biology; Schwiertz, A., Ed.; Springer: Cham, Switzerland, 2016; Volume 902, pp. 61–81. [Google Scholar]
- Cho, H.-W.; Eom, Y.-B. Forensic Analysis of Human Microbiome in Skin and Body Fluids Based on Geographic Location. Front. Cell. Infect. Microbiol. 2021, 11, 695191. [Google Scholar] [CrossRef] [PubMed]
- Bastianelli, C.; Farris, M.; Bianchi, P.; Benagiano, G. The effect of different contraceptive methods on the vaginal microbiome. Expert Rev. Clin. Pharmacol. 2021, 14, 1–16. [Google Scholar]
- Zárate, G.; Santos, V.; Nader-Macias, M.E. Protective effect of vaginal Lactobacillus paracasei CRL 1289 against urogeni-tal infection produced by Staphylococcus aureus in a mouse animal model. Infect. Dis. Obstet. Gynecol. 2007, 2007, 048358. [Google Scholar]
- Balzaretti, S.; Taverniti, V.; Rondini, G.; Marcolegio, G.; Minuzzo, M.; Remagni, M.C.; Fiore, W.; Arioli, S.; Guglielmetti, S. The vaginal isolate Lactobacillus paracasei LPC-S01 (DSM 26760) is suitable for oral administration. Front. Microbiol. 2015, 6, 952. [Google Scholar] [CrossRef]
- Gueniche, A.; Benyacoub, J.; Philippe, D.; Bastien, P.; Kusy, N.; Breton, L.; Blum, S.; Castiel-Higounenc, I. Lactobacillus paracasei CNCM I-2116 (ST11) inhibits substance P-induced skin inflammation and accelerates skin barrier function recovery in vitro. Eur. J. Dermatol. 2010, 20, 731–737. [Google Scholar]
- Jeong, J.H.; Lee, C.Y.; Chung, D.K. Probiotic lactic acid bacteria and skin health. Crit. Rev. Food Sci. Nutr. 2016, 56, 2331–2337. [Google Scholar]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [Green Version]
- Lange-Asschenfeldt, B.; Marenbach, D.; Lang, C.; Patzelt, A.; Ulrich, M.; Maltusch, A.; Terhorst, D.; Stockfleth, E.; Sterry, W.; Lademann, J. Distribution of Bacteria in the Epidermal Layers and Hair Follicles of the Human Skin. Ski. Pharmacol. Physiol. 2011, 24, 305–311. [Google Scholar] [CrossRef]
- Vongsa, R.; Hoffman, D.; Shepard, K.; Koenig, D. Comparative study of vulva and abdominal skin microbiota of healthy females with high and average BMI. BMC Microbiol. 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Carmina, E.; Lobo, R.A. Evaluation of hormonal status. In Yen and Jaffe’s Reproductive Endocrinology: Physiology, Pathophysiology and Clinical Management, 6th ed.; Strauss, J.F., III, Barbieri, R.L., Eds.; Elsevier: Saunders, PA, USA, 2009; pp. 801–823. [Google Scholar]
- Santen, R.J.; Mirkin, S.; Bernick, B.; Constantine, G.D. Systemic estradiol levels with low-dose vaginal estrogens. Menopause 2019, 27, 361–370. [Google Scholar] [CrossRef]
- Zhurina, M.V.; Gannesen, A.V.; Mart’Yanov, S.V.; Teteneva, N.A.; Shtratnikova, V.Y.; Plakunov, V.K. Niclosamide as a promising antibiofilm agent. Microbiology 2017, 86, 455–462. [Google Scholar] [CrossRef]
- Danilova, N.D.; Solovyeva, T.V.; Mart’Yanov, S.V.; Zhurina, M.V.; Gannesen, A.V. Stimulatory Effect of Epinephrine on Biofilms of Micrococcus luteus C01. Microbiology 2020, 89, 493–497. [Google Scholar] [CrossRef]
- Lynch, C. Vaginal Estrogen Therapy for the Treatment of Atrophic Vaginitis. J. Women’s Health 2009, 18, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Mart’Yanov, S.; Botchkova, E.; Plakunov, V.; Gannesen, A. The Impact of Norepinephrine on Mono-Species and Dual-Species Staphylococcal Biofilms. Microorganisms 2021, 9, 820. [Google Scholar] [CrossRef]
- Plakunov, V.K.; Mart’yanov, S.V.; Teteneva, N.A.; Zhurina, M.V. A universal method for quantitative characteriza-tion of growth and metabolic activity of microbial biofilms in static models. Microbiology 2016, 85, 509–513. [Google Scholar]
- Ovcharova, M.; Geraskina, O.; Danilova, N.; Botchkova, E.; Martyanov, S.; Feofanov, A.; Plakunov, V.; Gannesen, A. Atrial Natriuretic Peptide Affects Skin Commensal Staphylococcus epidermidis and Cutibacterium acnes Dual-Species Biofilms. Microorganisms 2021, 9, 552. [Google Scholar] [CrossRef]
- Lebeer, S.; Verhoeven, T.L.A.; Claes, I.J.J.; De Hertogh, G.; Vermeire, S.; Buyse, J.; Van Immerseel, F.; Vanderleyden, J.; De Keersmaecker, S.C.J. FISH analysis of Lactobacillus biofilms in the gastrointestinal tract of different hosts. Lett. Appl. Microbiol. 2011, 52, 220–226. [Google Scholar] [CrossRef]
- Gannesen, A.V.; Borrel, V.; Lefeuvre, L.; Netrusov, A.I.; Plakunov, V.K.; Feuilloley, M.G.J. Effect of two cosmetic compounds on the growth, biofilm formation activity, and surface properties of acneic strains of Cutibacterium acnes and Staphylococcus aureus. MicrobiologyOpen 2019, 8, e00659. [Google Scholar] [CrossRef]
- Dąbrowska, A.K.; Rotaru, G.-M.; Derler, S.; Spano, F.; Camenzind, M.; Annaheim, S.; Stämpfli, R.; Schmid, M.; Rossi, R.M. Materials used to simulate physical properties of human skin. Ski. Res. Technol. 2016, 22, 3–14. [Google Scholar] [CrossRef]
- Brown, T.M.; Krishnamurthy, K. Histology, Dermis. StatPearls Publishing, Treasure Island (FL). 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK535346/ (accessed on 11 February 2022).
- Elleuch, L.; Shaaban, M.; Smaoui, S.; Mellouli, L.; Karray-Rebai, I.; Fourati-Ben Fguira, L.; Shaaban, K.A.; Laatsch, H. Bioac-tive secondary metabolites from a new terrestrial Streptomyces sp. TN262. Appl. Biochem. Biotechnol. 2010, 162, 579–593. [Google Scholar]
- Clabaut, M.; Suet, A.; Racine, P.J.; Tahrioui, A.; Verdon, J.; Barreau, M.; Maillot, O.; Le Tirant, A.; Karsybayeva, M.; Kremser, C.; et al. Effect of 17β-estradiol on a hu-man vaginal Lactobacillus crispatus strain. Sci. Rep. 2021, 11, 1–16. [Google Scholar]
- Clabaut, M.; Boukerb, A.M.; Ben Mlouka, A.; Suet, A.; Tahrioui, A.; Verdon, J.; Barreau, M.; Maillot, O.; Le Tirant, A.; Karsybayeva, M.; et al. Variability of the response of human vaginal Lactobacillus crispatus to 17β-estradiol. Sci. Rep. 2021, 11, 11533. [Google Scholar] [CrossRef] [PubMed]
- Plakunov, V.K.; Nikolaev, Y.A.; Gannesen, A.V.; Chemaeva, D.S.; Zhurina, M.V. A New Approach to Detection of the Protective Effect of Escherichia coli on Gram-Positive Bacteria in Binary Biofilms in the Presence of Antibiotics. Microbiology 2019, 88, 275–281. [Google Scholar] [CrossRef]
- Mukamolova, G.V.; Murzin, A.G.; Salina, E.G.; Demina, G.R.; Kell, D.B.; Kaprelyants, A.S.; Young, M. Muralytic activity of Micrococcus luteus Rpf and its relationship to physiological activity in promoting bacterial growth and resuscitation. Mol. Microbiol. 2006, 59, 84–98. [Google Scholar]
- Mukamolova, G.V.; Turapov, O.A.; Kazarian, K.; Telkov, M.; Kaprelyants, A.S.; Kell, D.B.; Young, M. The rpf gene of Micrococcus luteus encodes an essential secreted growth factor. Mol. Microbiol. 2002, 46, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Vidaillac, C.; Yong, V.F.L.; Aschtgen, M.S.; Qu, J.; Yang, S.; Xu, G.; Seng, Z.J.; Brown, A.C.; Ali, M.K.; Jaggi, T.K.; et al. Sex Steroids induce membrane stress responses and virulence properties in Pseudomonas aeruginosa. mBio 2020, 11, e01774-20, Erratum in mBio, 3 November 2020. [Google Scholar]
- Plotkin, B.J.; Konakieva, M.I. Attenuation of antimicrobial activity by the human steroid hormones. Steroids 2017, 128, 120–127. [Google Scholar] [CrossRef]
- Chatterjee, I.; Somerville, G.A.; Heilmann, C.; Sahl, H.-G.; Maurer, H.H.; Herrmann, M. Very Low Ethanol Concentrations Affect the Viability and Growth Recovery in Post-Stationary-Phase Staphylococcus aureus Populations. Appl. Environ. Microbiol. 2006, 72, 2627–2636. [Google Scholar] [CrossRef] [Green Version]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshaghabee, F.; Ebockelmann, W.; Emeske, D.; Vrese, M.E.; Ewalte, H.-G.; Eschrezenmeir, J.; Heller, K.J. Ethanol Production by Selected Intestinal Microorganisms and Lactic Acid Bacteria Growing under Different Nutritional Conditions. Front. Microbiol. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Ainala, S.K.; Park, S. Metabolic engineering of Lactobacillus reuteri DSM 20,016 for improved 1,3-propanediol production from glycerol. Bioresour. Technol. 2021, 338, 125590. [Google Scholar] [CrossRef] [PubMed]
- Burgain, A.; Tebbji, F.; Khemiri, I.; Sellam, A. Metabolic Reprogramming in the Opportunistic Yeast Candida albicans in Response to Hypoxia. mSphere 2020, 5, e00913-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).