Increased Intraspecies Diversity in Escherichia coli Biofilms Promotes Cellular Growth at the Expense of Matrix Production

 ,
,  , and
, and 
Abstract

1. Introduction
2. Results and Discussion
2.1. Single- and Multi-Strain Biofilm Growth
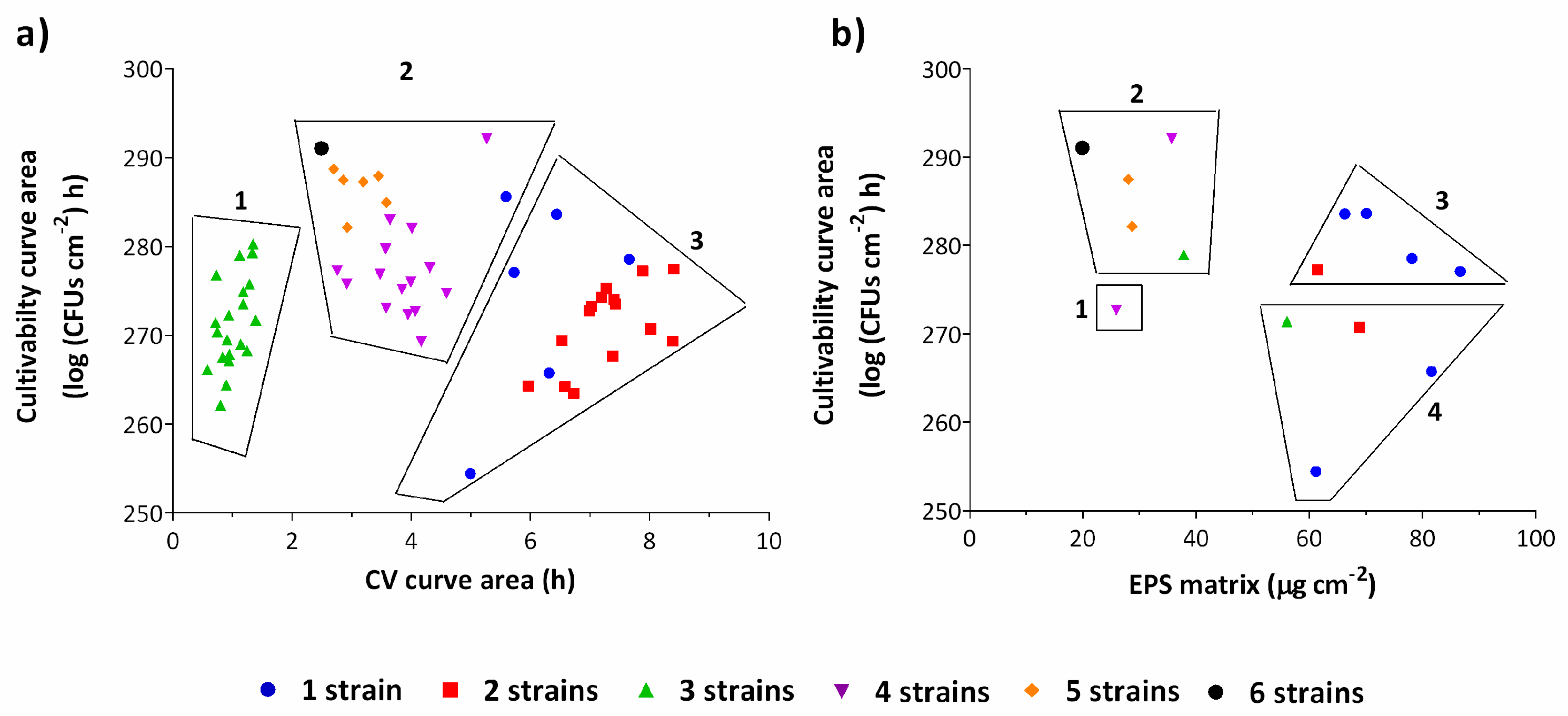
2.2. Biomass and Matrix Production as a Function of the Number of Strains in a Consortium
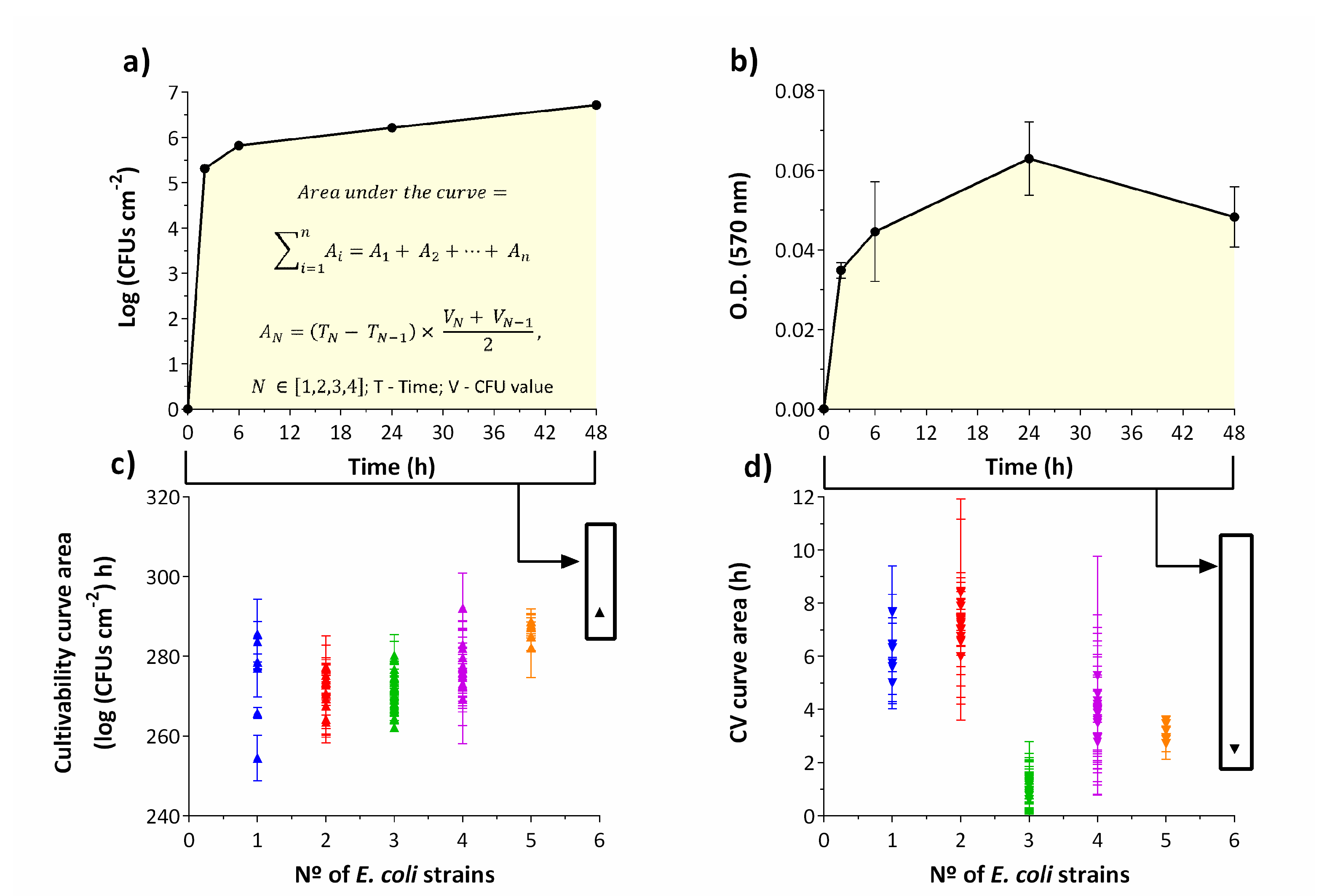
2.3. How Long Does It Take for Microorganisms to Adapt Their Metabolism?
2.4. Impact of Phylogenetic Closeness in Biomass Production for Two-Strain Biofilms
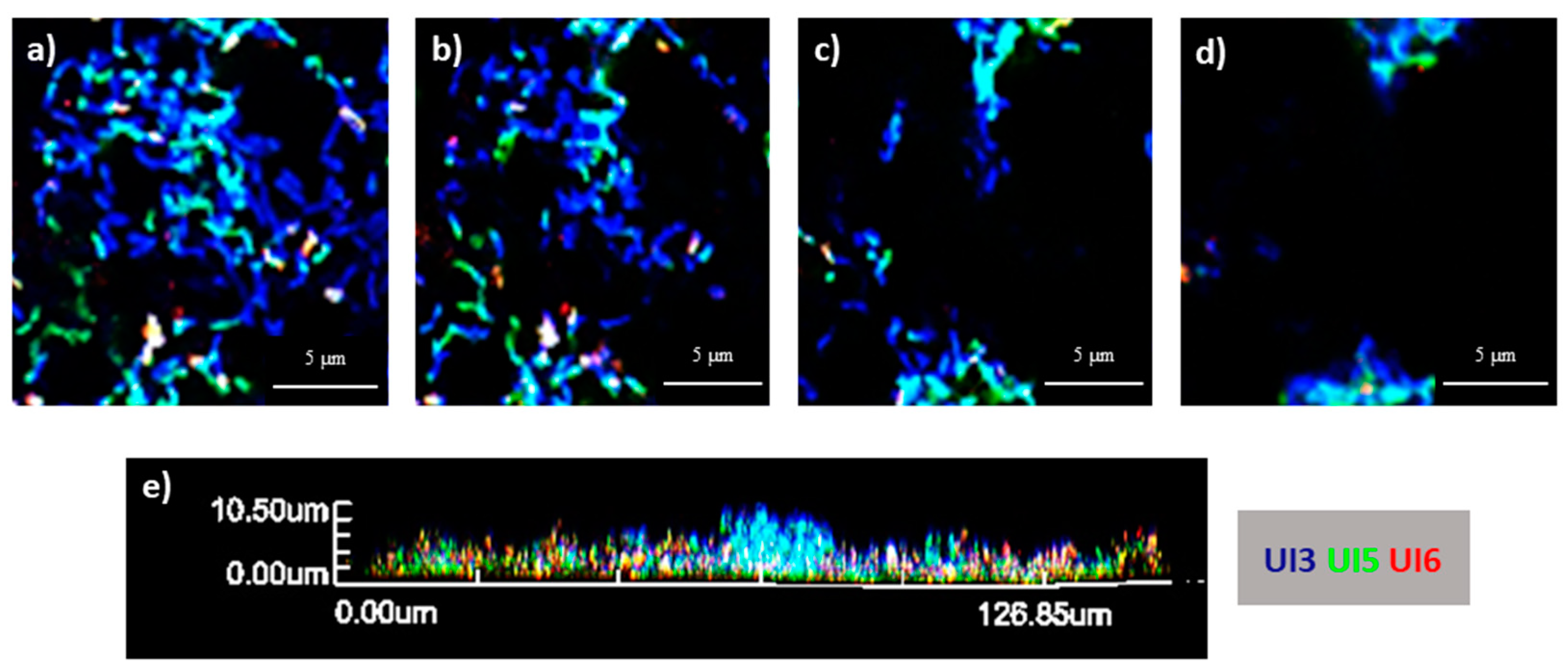
2.5. Spatial Organization of Biofilms Using PNA-FISH Combined with CLSM
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
3.2. Single- and Multi-Strain Biofilms Assays
3.3. Biomass Quantification by Crystal Violet Staining
3.4. Cultivability Assessment
3.5. EPS Matrix Quantification by Dry Weight
3.6. Multilocus Sequence Typing for Phylogenetic Analysis
3.7. Impact of the Phylogenetic Closeness in Biomass Production
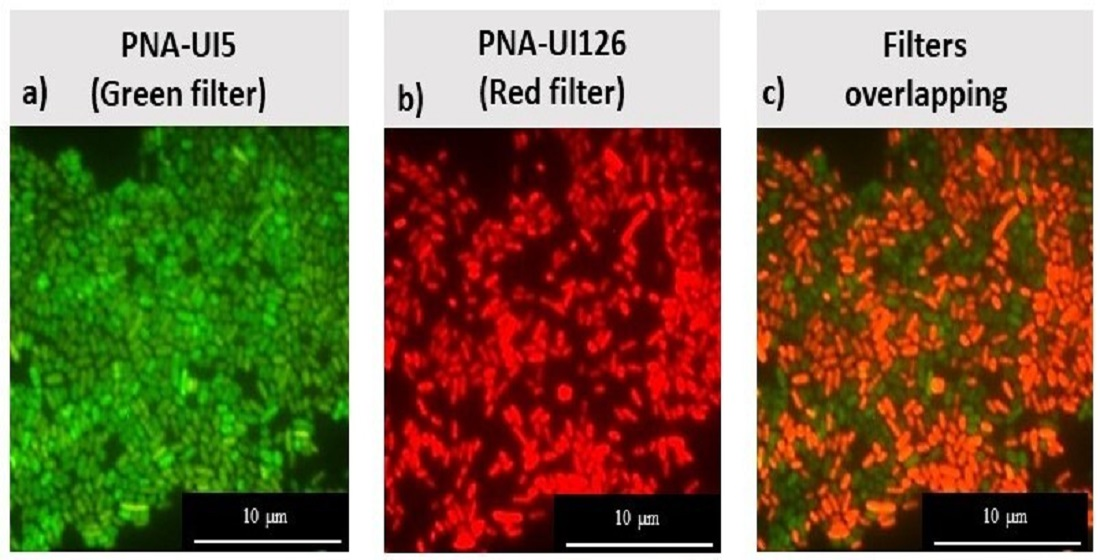
3.8. Optimization of the PNA-FISH Protocol
3.9. Study of the Spatial Organization in E. coli Multi-Strain Biofilms Using PNA-FISH Combined with Confocal Laser Scanning Microscopy Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Costerton, J.W.; Cheng, K.-J.; Geesey, G.G.; Ladd, T.I.; Curtis, N.J.; Mrinal, D.; Marrie, T.J. Bacterial Biofilms in Nature and Disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef]
- Donlan, R. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial Biofilms: From the Natural Environment to Infectious Diseases. Nat. Rev. Genet. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Czárán, T.; Hoekstra, R.F. Microbial Communication, Cooperation and Cheating: Quorum Sensing Drives the Evolution of Cooperation in Bacteria. PLoS ONE 2009, 4, e6655. [Google Scholar] [CrossRef]
- Azevedo, A.S.; Almeida, C.; Melo, L.F.; Azevedo, N.F. Impact of Polymicrobial Biofilms in Catheter-Associated Urinary Tract Infections. Crit. Rev. Microbiol. 2016, 43, 423–439. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial Competition: Surviving and Thriving in the Microbial Jungle. Nat. Rev. Genet. 2010, 8, 15–25. [Google Scholar] [CrossRef]
- Rendueles, O.; Ghigo, J.-M. Mechanisms of Competition in Biofilm Communities. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef]
- Gao, C.; Zhang, M.; Wu, Y.; Huang, Q.; Cai, P. Divergent Influence to a Pathogen Invader by Resident Bacteria with Different Social Interactions. Microbiol. Ecol. 2018, 77, 76–86. [Google Scholar] [CrossRef]
- Nadell, C.D.; Drescher, C.D.N.K.; Foster, K.R. Spatial Structure, Cooperation and Competition in Biofilms. Nat. Rev. Genet. 2016, 14, 589–600. [Google Scholar] [CrossRef]
- López, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-Specific Antibiotic Tolerance and Resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- Lee, K.W.K.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; A Rice, S. Biofilm Development and Enhanced Stress Resistance of a Model, Mixed-Species Community Biofilm. ISME J. 2014, 8, 894–907. [Google Scholar] [CrossRef]
- Kjaergaard, K.; Schembri, M.A.; Ramos, C.; Molin, S.; Klemm, P. Antigen 43 Facilitates Formation of Multispecies Biofilms. Environ. Microbiol. 2000, 2, 695–702. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, H.-C.F.J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S.A.R.S. Biofilms: An Emergent form of Bacterial Life. Nat. Rev. Genet. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Sztajer, H.; Szafranski, S.P.; Tomasch, J.; Reck, M.; Nimtz, M.; Rohde, M.; Wagner-Döbler, I. Cross-Feeding and Interkingdom Communication in Dual-Species Biofilms of Streptococcus Mutans and Candida Albicans. ISME J. 2014, 8, 2256–2271. [Google Scholar] [CrossRef]
- West, S.A.; Griffin, A.S.; Gardner, A.; Diggle, S.P. Social Evolution Theory for Microorganisms. Nat. Rev. Genet. 2006, 4, 597–607. [Google Scholar] [CrossRef]
- Xavier, J.B. Social Interaction in Synthetic and Natural Microbial Communities. Mol. Syst. Biol. 2011, 7, 483. [Google Scholar] [CrossRef]
- West, S.A.; Diggle, S.P.; Buckling, A.; Gardner, A.; Griffins, A.S. The Social Lives of Microbes. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 53–77. [Google Scholar] [CrossRef]
- Jiricny, N.; Diggle, S.P.; A West, S.; Evans, B.A.; Ballantyne, G.; Ross-Gillespie, A.; Griffin, A.S. Fitness Correlates with the Extent of Cheating in a Bacterium. J. Evol. Biol. 2010, 23, 738–747. [Google Scholar] [CrossRef]
- Nadell, C.D.; Xavier, J.B.; Foster, K.R. The Sociobiology of Biofilms. FEMS Microbiol. Rev. 2009, 33, 206–224. [Google Scholar] [CrossRef]
- Stefanic, P.; Decorosi, F.; Viti, C.; Petito, J.; Cohan, F.M.; Mandic-Mulec, I. The Quorum Sensing Diversity within and between Ecotypes of Bacillus Subtilis. Environ. Microbiol. 2012, 14, 1378–1389. [Google Scholar] [CrossRef]
- Lee, K.W.K.; Yam, J.K.H.; Mukherjee, M.; Periasamy, S.; Steinberg, P.D.; Kjelleberg, S.; Rice, S.A. Interspecific Diversity Reduces and Functionally Substitutes for Intraspecific Variation in Biofilm Communities. ISME J. 2016, 10, 846–857. [Google Scholar] [CrossRef]
- Oluyombo, O.; Penfold, C.N.; Diggle, S.P. Competition in Biofilms between Cystic Fibrosis Isolates of Pseudomonas aeruginosa Is Shaped by R-Pyocins. mBio 2019, 10, e01828-18. [Google Scholar] [CrossRef]
- CDCP, Centers for Disease Control and Prevention. Catheter-Associated Urinary Tract Infections (CAUTI). 2019. Available online: https://www.cdc.gov/hai/ca_uti/uti.html (accessed on 12 November 2019).
- Azevedo, N.F.; Almeida, C.; Fernandes, I.; Cerqueira, L.; Dias, S.; Keevil, C.W.; Vieira, M.J. Survival of Gastric and Enterohepatic Helicobacter spp. in Water: Implications for Transmission. Appl. Environ. Microbiol. 2008, 74, 1805–1811. [Google Scholar] [CrossRef]
- Almeida, C.; Azevedo, N.F.; Santos, S.R.B.; Keevil, C.W.; Vieira, M.J. Discriminating Multi-Species Populations in Biofilms with Peptide Nucleic Acid Fluorescence in Situ Hybridization (PNA FISH). PLoS ONE 2011, 6, e14786. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Survival of Escherichia Coli in the Environment: Fundamental and Public Health Aspects. ISME J. 2011, 5, 367. [Google Scholar] [CrossRef]
- López-Maury, L.; Marguerat, S.; Bähler, J. Tuning Gene Expression to Changing Environments: From Rapid Responses to Evolutionary Adaptation. Nat. Rev. Genet. 2008, 9, 583–593. [Google Scholar] [CrossRef]
- Drazic, A.; Kutzner, E.; Winter, J.; Eisenreich, W. Metabolic Response of Escherichia Coli Upon Treatment with Hypochlorite at Sub-Lethal Concentrations. PLoS ONE 2015, 10, e0125823. [Google Scholar] [CrossRef]
- Lukjancenko, O.; Wassenaar, T.M.; Ussery, D.W. Comparison of 61 Sequenced Escherichia Coli Genomes. Microbiol. Ecol. 2010, 60, 708–720. [Google Scholar] [CrossRef]
- Liu, W.; Røder, H.L.; Madsen, J.S.; Bjarnsholt, T.; Sørensen, S.J.; Burmølle, M. Interspecific Bacterial Interactions Are Reflected in Multispecies Biofilm Spatial Organization. Front. Microbiol. 2016, 7, 1366. [Google Scholar] [CrossRef]
- Azevedo, A.S.; Almeida, C.; Pereira, B.; Madureira, P.; Wengel, J.; Azevedo, N.F. Detection and Discrimination of Biofilm Populations Using Locked Nucleic Acid/2′-O-Methyl-Rna Fluorescence in Situ Hybridization (LNA/2′OMe-FISH). Biochem. Eng. J. 2015, 104, 64–73. [Google Scholar] [CrossRef]
- Azevedo, A.S.; Almeida, C.; Pereira, B.; Melo, L.F.; Azevedo, N.F. Impact ofDelftia Tsuruhatensis and Achromobacter Xylosoxidans on Escherichia Colidual-Species Biofilms Treated with Antibiotic Agents. Biofouling 2016, 32, 227–241. [Google Scholar] [CrossRef]
- Ledder, R.G.; Timperley, A.S.; Friswell, M.K.; Macfarlane, S.; McBain, A.J. Coaggregation between and among Human Intestinal and Oral Bacteria. FEMS Microbiol. Ecol. 2008, 66, 630–636. [Google Scholar] [CrossRef]
- Brooks, T.; Keevil, C. A Simple Artificial Urine for the Growth of Urinary Pathogens. Lett. Appl. Microbiol. 1997, 24, 203–206. [Google Scholar] [CrossRef]
- Azevedo, A.S.; Almeida, C.; Melo, L.F.; Azevedo, N.F. Interaction between Atypical Microorganisms and E. Coli in Catheter-Associated Urinary Tract Biofilms. Biofouling 2014, 30, 893–902. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A Modified Microtiter-Plate Test for Quantification of Staphylococcal Biofilm Formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Azeredo, J.; Lazarova, V.; Oliveira, R. Methods to Extract the Exopolymeric Matrix from Biofilms: A Comparative Study. Water Sci. Technol. 1999, 39, 243–250. [Google Scholar] [CrossRef]
- Azeredo, J.; Henriques, M.; Sillankorva, S.; Oliveira, R. Extraction of Exopolymers from Biofilms: The Protective Effect of Glutaraldehyde. Water Sci. Technol. 2003, 47, 175–179. [Google Scholar] [CrossRef]
- Liu, X.; Thungrat, K.; Boothe, D.M. Multilocus Sequence Typing and Virulence Profiles in Uropathogenic Escherichia Coli Isolated from Cats in the United States. PLoS ONE 2015, 10, e0143335. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Baptista, J.; Simões, M.; Borges, A. Effect of Plant-Based Catecholic Molecules on the Prevention and Eradication of Pre-Established Escherichia Coli Biofilms: A Structure Activity Relationship Study. Int. Biodeterior. Biodegrad. 2018, 141, 101–113. [Google Scholar] [CrossRef]
- Almeida, C.; Azevedo, N.F.; Fernandes, R.M.; Keevil, C.W.; Vieira, M.J. Fluorescence In Situ Hybridization Method Using a Peptide Nucleic Acid Probe for Identification of Salmonella spp. in a Broad Spectrum of Samples. Appl. Environ. Microbiol. 2010, 76, 4476–4485. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, N.F.; Pacheco, A.; Keevil, C.W.; Vieira, M.J. Adhesion of Water Stressed Helicobacter Pylori to Abiotic Surfaces. J. Appl. Microbiol. 2006, 101, 718–724. [Google Scholar] [CrossRef]
- McLachlan, G. 9 The Classification and Mixture Maximum Likelihood Approaches to Cluster Analysis. Handb. Stat. 1982, 2, 199–208. [Google Scholar] [CrossRef]
- Baudry, J.-P.; Raftery, A.E.; Celeux, G.; Lo, K.; Gottardo, R. Combining Mixture Components for Clustering. J. Comput. Graph. Stat. 2010, 19, 332–353. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How Antibiotics Kill Bacteria: From Targets to Networks. Nat. Rev. Genet. 2010, 8, 423–435. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PNA-Probes | Strains | Sequence | Position in 16S rRNA | bp | % GC | ΔG (kcal/mol) | Tm (°C) |
|---|---|---|---|---|---|---|---|
| PNA-UI126 | UI1, 2 and 6 | 5′-GTGAGCCTTTACCC-3′ | 144 to 157 | 14 | 0.57 | −16.43 | 73.30 |
| PNA-UI5 | UI5 | 5′-TCCATCGGGCAGT-3′ | 18 to 30 | 13 | 0.62 | −15.88 | 75.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azevedo, A.S.; Gerola, G.P.; Baptista, J.; Almeida, C.; Peres, J.; Mergulhão, F.J.; Azevedo, N.F. Increased Intraspecies Diversity in Escherichia coli Biofilms Promotes Cellular Growth at the Expense of Matrix Production. Antibiotics 2020, 9, 818. https://doi.org/10.3390/antibiotics9110818
Azevedo AS, Gerola GP, Baptista J, Almeida C, Peres J, Mergulhão FJ, Azevedo NF. Increased Intraspecies Diversity in Escherichia coli Biofilms Promotes Cellular Growth at the Expense of Matrix Production. Antibiotics. 2020; 9(11):818. https://doi.org/10.3390/antibiotics9110818
Chicago/Turabian StyleAzevedo, Andreia S., Gislaine P. Gerola, João Baptista, Carina Almeida, Joana Peres, Filipe J. Mergulhão, and Nuno F. Azevedo. 2020. "Increased Intraspecies Diversity in Escherichia coli Biofilms Promotes Cellular Growth at the Expense of Matrix Production" Antibiotics 9, no. 11: 818. https://doi.org/10.3390/antibiotics9110818
APA StyleAzevedo, A. S., Gerola, G. P., Baptista, J., Almeida, C., Peres, J., Mergulhão, F. J., & Azevedo, N. F. (2020). Increased Intraspecies Diversity in Escherichia coli Biofilms Promotes Cellular Growth at the Expense of Matrix Production. Antibiotics, 9(11), 818. https://doi.org/10.3390/antibiotics9110818