Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi


Abstract
1. Introduction
2. Materials and Methods
2.1. Organism and Culture Conditions
2.2. Essential Oils and Drugs
2.3. Microscopy
2.4. Evaluation of Essential Oils for Their Activities Against B. Burgdorferi Stationary Phase Cultures
2.5. Essential Oil and Drug Susceptibility Testing
2.6. Subculture Studies to Assess Viability of Essential Oil-Treated B. Burgdorferi Organisms
3. Results
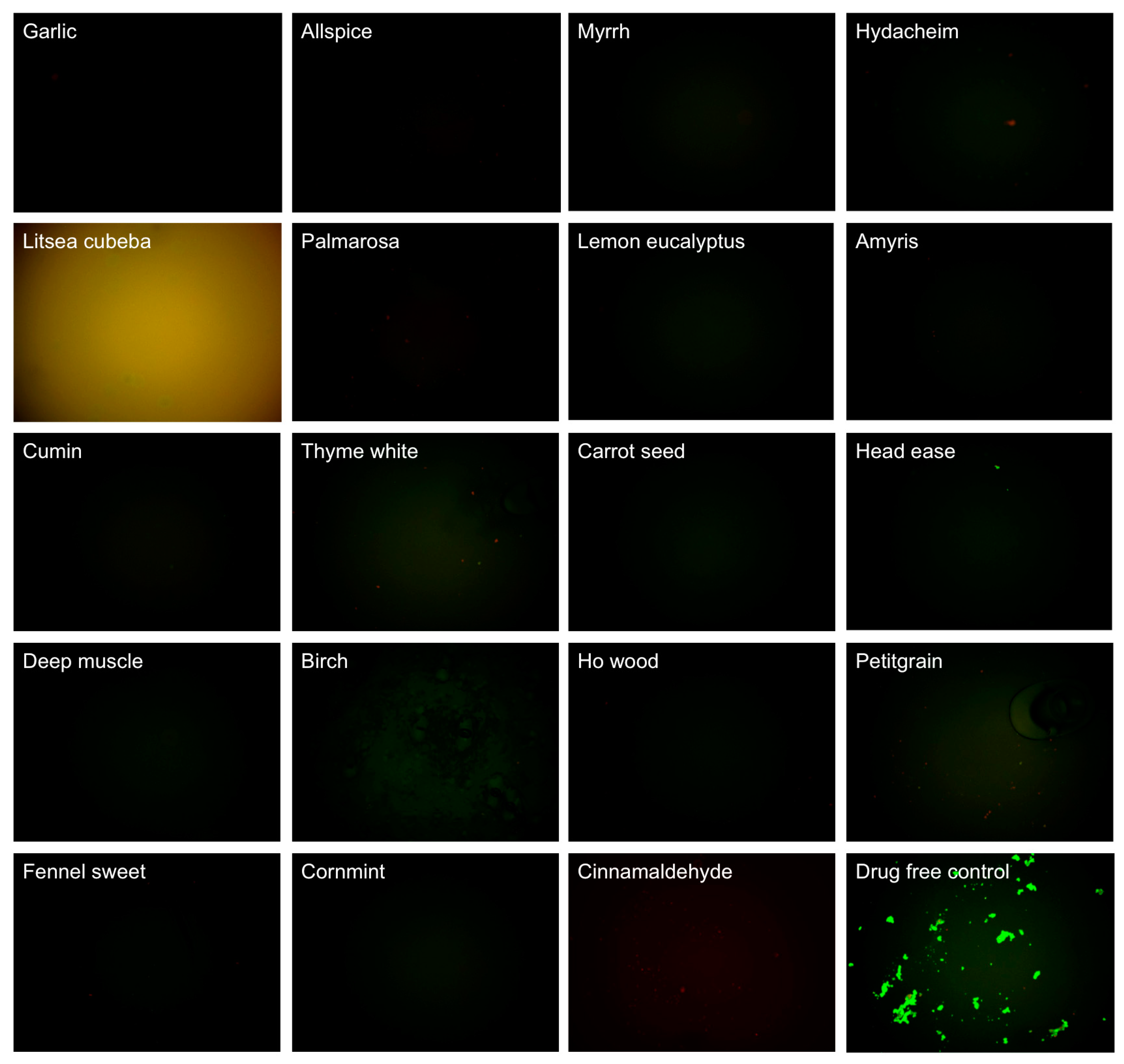
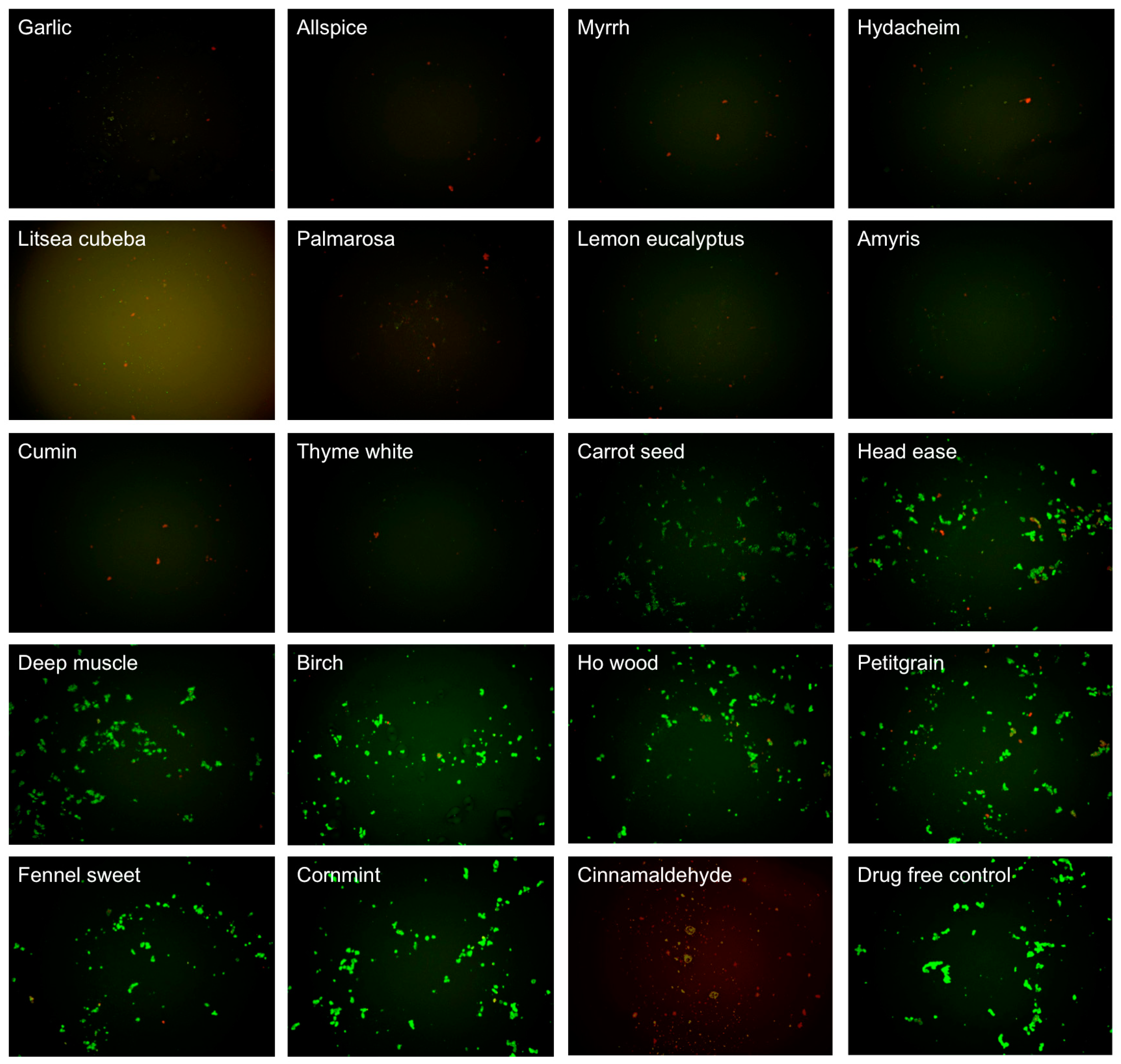
3.1. Evaluating Activity of Essential Oils Against Stationary Phase B. Burgdorferi
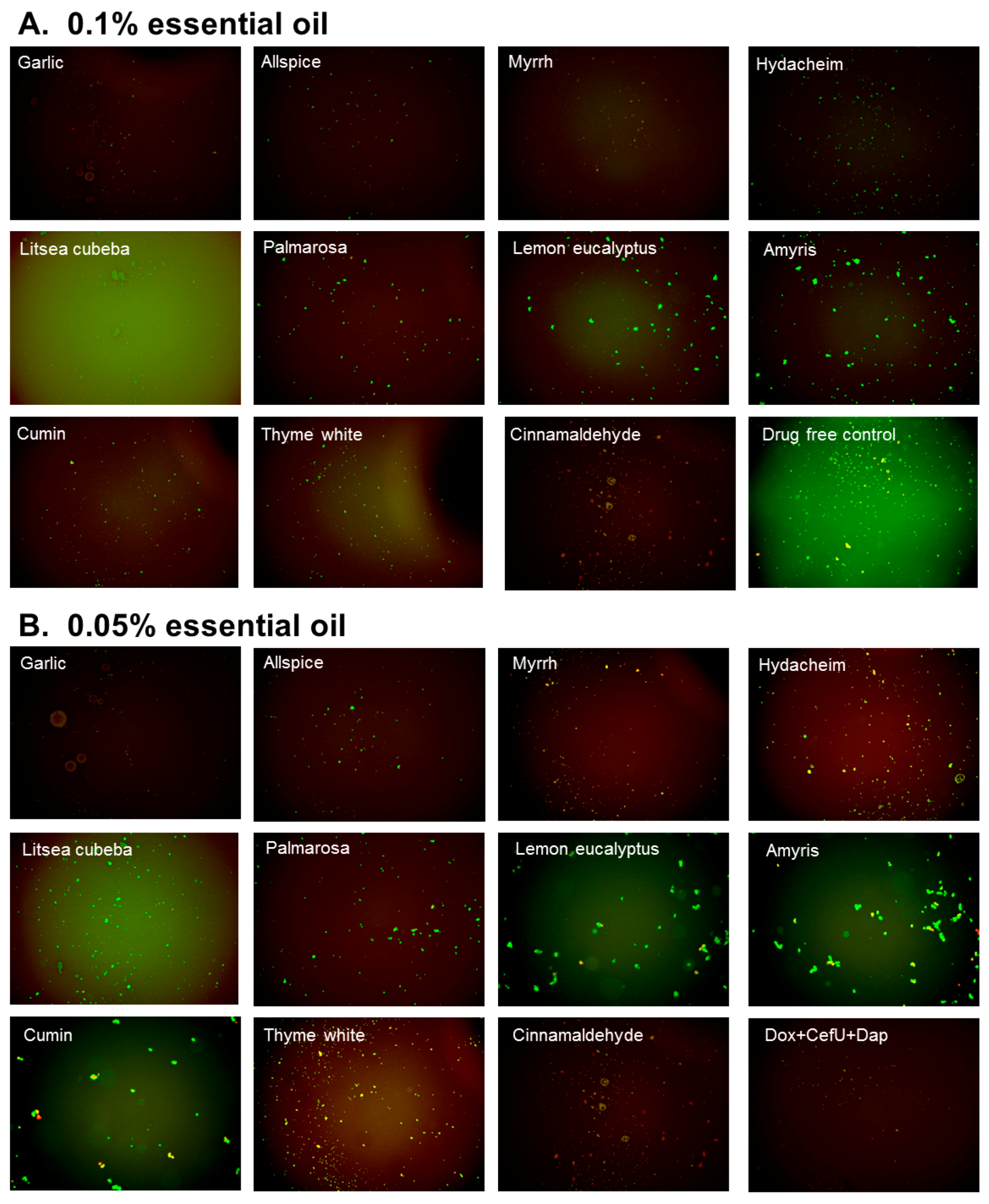
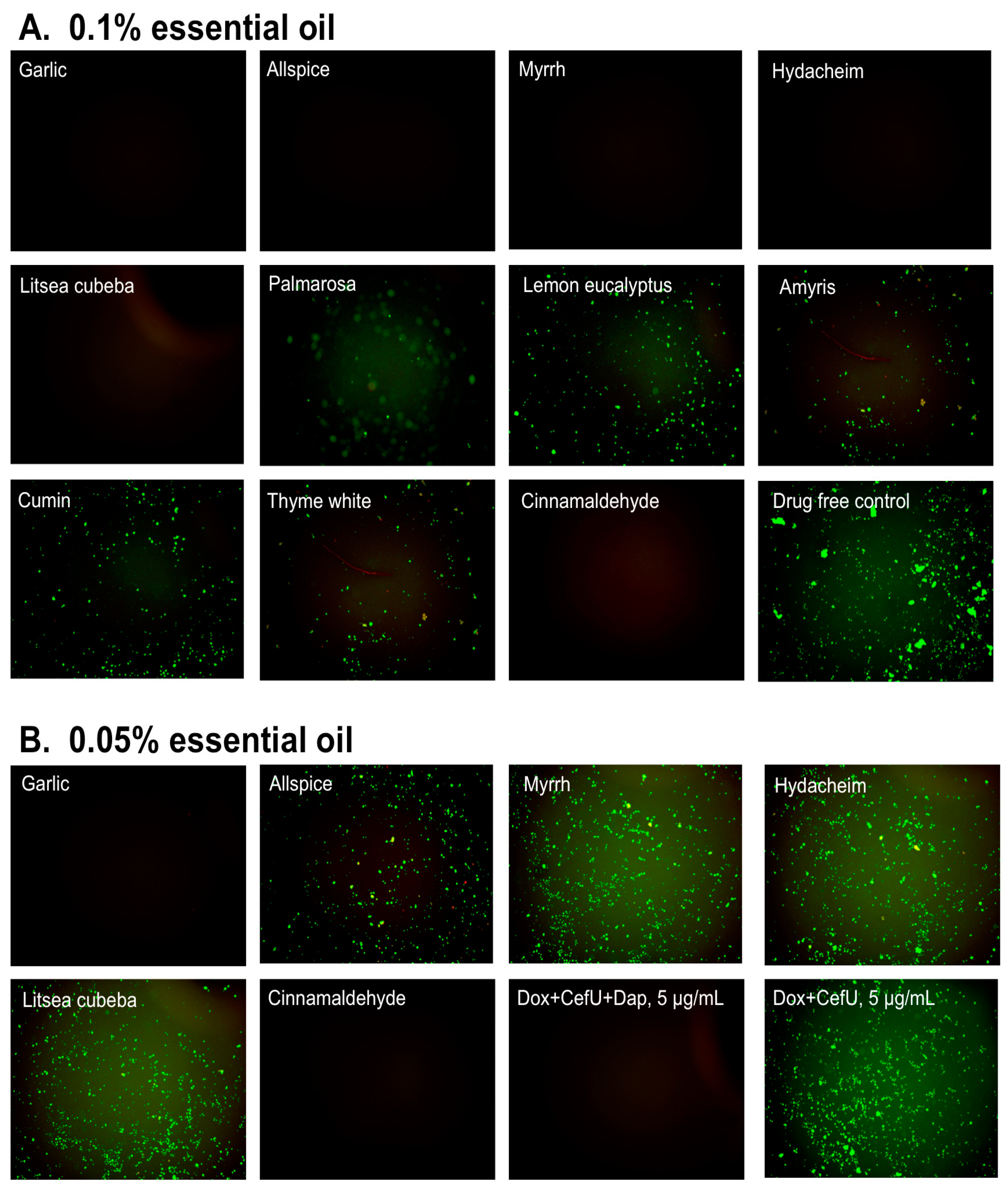
3.2. Subculture Studies to Evaluate the Activity of Essential Oils Against Stationary Phase B. burgdorferi
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CDC. Lyme Disease. Available online: http://www.cdc.gov/lyme/ (accessed on 7 March 2018).
- Wormser, G.P.; Dattwyler, R.J.; Shapiro, E.D.; Halperin, J.J.; Steere, A.C.; Klempner, M.S.; Krause, P.J.; Bakken, J.S.; Strle, F.; Stanek, G.; et al. The clinical assessment, treatment, and prevention of lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the infectious diseases society of America. Clin. Infect. Dis. 2006, 43, 1089–1134. [Google Scholar] [CrossRef] [PubMed]
- Aucott, J.N.; Rebman, A.W.; Crowder, L.A.; Kortte, K.B. Post-treatment Lyme disease syndrome symptomatology and the impact on life functioning: Is there something there? Qual. Life Res. 2013, 22, 75. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Gross, D.; Meyer, A.L.; Huber, B.T. Autoimmune mechanisms in antibiotic treatment-resistant Lyme arthritis. J. Autoimmun. 2001, 16, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Bockenstedt, L.K.; Gonzalez, D.G.; Haberman, A.M.; Belperron, A.A. Spirochete antigens persist near cartilage after murine Lyme borreliosis therapy. J. Clin. Investig. 2012, 122, 2652–2660. [Google Scholar] [CrossRef] [PubMed]
- Hodzic, E.; Imai, D.; Feng, S.; Barthold, S.W. Resurgence of Persisting Non-Cultivable Borrelia burgdorferi following Antibiotic Treatment in Mice. PLoS ONE 2014, 9, e86907. [Google Scholar] [CrossRef] [PubMed]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia burgdorferi in rhesus macaques following antibiotic treatment of disseminated infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Hodzic, E.; Feng, S.; Holden, K.; Freet, K.J.; Barthold, S.W. Persistence of Borrelia burgdorferi following antibiotic treatment in mice. Antimicrob. Agents Chemother. 2008, 52, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Telford, S.R., 3rd; Turk, S.P.; Chung, E.; Williams, C.; Dardick, K.; Krause, P.J.; Brandeburg, C.; Crowder, C.D.; Carolan, H.E.; et al. Xenodiagnosis to detect Borrelia burgdorferi infection: A first-in-human study. Clin. Infect. Dis. 2014, 58, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Straubinger, R.K.; Summers, B.A.; Chang, Y.F.; Appel, M.J. Persistence of Borrelia burgdorferi in experimentally infected dogs after antibiotic treatment. J. Clin. Microbiol. 1997, 35, 111–116. [Google Scholar] [PubMed]
- Zhang, Y. Persistent and dormant tubercle bacilli and latent tuberculosis. Front. Biosci. 2004, 9, 1136–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yew, W.W.; Barer, M.R. Targeting persisters for tuberculosis control. Antimicrob. Agents Chemother. 2012, 56, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Brown, A.V.; Matluck, N.E.; Hu, L.T.; Lewis, K. Borrelia burgdorferi, the causative agent of Lyme disease, forms drug-tolerant persister cells. Antimicrob. Agents Chemother. 2015, 59, 4616–4624. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Auwaerter, P.G.; Zhang, Y. Drug Combinations against Borrelia burgdorferi persisters in vitro: Eradication achieved by using daptomycin, cefoperazone and doxycycline. PLoS ONE 2015, 10, e0117207. [Google Scholar] [CrossRef] [PubMed]
- Caskey, J.R.; Embers, M.E. Persister development by Borrelia burgdorferi populations in vitro. Antimicrob. Agents Chemother. 2015, 59, 6288–6295. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, T.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. Identification of novel activity against Borrelia burgdorferi persisters using an FDA approved drug library. Emerg. Microbes Infect. 2014, 3, e49. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, S.; Shi, W.; Zhang, Y. Ceftriaxone pulse dosing fails to eradicate biofilm-like microcolony B. burgdorferi persisters which are sterilized by daptomycin/doxycycline/cefuroxime without pulse dosing. Front. Microbiol. 2016, 7, 1744. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, S.; Shi, W.; Zubcevik, N.; Miklossy, J.; Zhang, Y. Selective Essential oils from spice or culinary herbs have high activity against stationary phase and biofilm Borrelia burgdorferi. Front. Med. 2017, 4, 169. [Google Scholar] [CrossRef] [PubMed]
- GC/MS TESTING. Available online: https://www.theplantguru.com/gc-ms-testing (accessed on 13 September 2018).
- Wikler, M.A.; National Committee for Clinical Laboratory. Performance Standards for Antimicrobial Susceptibility Testing: Fifteenth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2005.
- The United States Pharmacopeial Convention. The United States Pharmacopeia, 24th ed.; The United States Pharmacopeial Convention: Philadelphia, PA, USA, 2000.
- Feng, J.; Wang, T.; Zhang, S.; Shi, W.; Zhang, Y. An Optimized SYBR Green I/PI Assay for Rapid Viability Assessment and Antibiotic Susceptibility Testing for Borrelia burgdorferi. PLoS ONE 2014, 9, e111809. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Shi, W.; Zhang, S.; Zhang, Y. Identification of new compounds with high activity against stationary phase Borrelia burgdorferi from the NCI compound collection. Emerg. Microbes Infect. 2015, 4, e31. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.D. Garlic: A Review of Its Medicinal Effects and Indicated Active Compounds. In Phytomedicines of Europe; American Chemical Society: Washington, DC, USA, 1998; pp. 176–209. [Google Scholar]
- Petrovska, B.B.; Cekovska, S. Extracts from the history and medical properties of garlic. Pharmacogn. Rev. 2010, 4, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Shelef, L.A.; Naglik, O.A.; Bogen, D.W. Sensitivity of some common food-borne bacteria to the spices sage, rosemary, and allspice. J. Food Sci. 1980, 45, 1042–1044. [Google Scholar] [CrossRef]
- Lisa, E.L.; Carac, G.; Barbu, V.; Robu, S. The synergistic antioxidant effect and antimicrobial efficacity of propolis, myrrh and chlorhexidine as beneficial toothpaste components. Rev. Chim.-Buchar. 2017, 68, 2060–2065. [Google Scholar]
- Dolara, P.; Corte, B.; Ghelardini, C.; Pugliese, A.M.; Cerbai, E.; Menichetti, S.; Lo Nostro, A. Local anaesthetic, antibacterial and antifungal properties of sesquiterpenes from myrrh. Planta Med. 2000, 66, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Ritu, A.; Avijit, M. Phytochemical screening and antimicrobial activity of rhizomes of hedychium spicatum. Phcog. J. 2017, 9. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y. Chemical composition and antibacterial activity of essential oils from different parts of Litsea cubeba. Chem. Biodivers. 2010, 7, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Chemistry, antimicrobial mechanisms, and antibiotic activities of cinnamaldehyde against pathogenic bacteria in animal feeds and human foods. J. Agric. Food Chem. 2017, 65, 10406–10423. [Google Scholar] [CrossRef] [PubMed]
- Subash Babu, P.; Prabuseenivasan, S.; Ignacimuthu, S. Cinnamaldehyde—A potential antidiabetic agent. Phytomedicine 2007, 14, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Feketa, V.V.; Marrelli, S.P. Systemic Administration of the TRPV3 Ion Channel Agonist Carvacrol Induces Hypothermia in Conscious Rodents. PLoS ONE 2015, 10, e0141994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, W.; Zhang, W.; Mitchison, D. Mechanisms of Pyrazinamide Action and Resistance. Microbiol. Spectr. 2013, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Permar, S.; Sun, Z. Conditions that may affect the results of susceptibility testing of Mycobacterium tuberculosis to pyrazinamide. J. Med. Microbiol. 2002, 51, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Niu, H.; Shi, W.; Zhang, S.; Zhang, H.; Margolick, J.; Zhang, W.; Zhang, Y. Disruption of membrane by colistin kills uropathogenic Escherichia coli persisters and enhances killing of other antibiotics. Antimicrob. Agents Chemother. 2016, 60, 6867–6871. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oils and Control Drugs | Plant and Extracted Part | Residual Viability (%) after 0.2% EO or 40 μM Antibiotic Treatment | Residual Viability (%) after 0.1% EO Treatment | ||||
|---|---|---|---|---|---|---|---|
| Plate Reader b | Microscope c | p-Value e | Plate Reader b | Microscope c | p-Value e | ||
| Doxycycline | -- | 73 ± 4 | 66 ± 2 | 1.000 | -- | -- | -- |
| Cefuroxime | -- | 58 ± 3 | 55 ± 3 | 0.0016 | -- | -- | -- |
| Daptomycin | -- | 28 ± 5 | 21 ± 1 | 0.0004 | -- | -- | -- |
| Cinnamaldehyde | -- | 26 ± 5 | 0 | 0.0002 | 54 ± 2 | 28 | 0.0016 |
| Garlic | Allium sativum L. (syn. Porrum sativum (L.) Rchb.), bulbs | 25 ± 4 | 0 | 0.0001 | 24 ± 6 | 18 ± 2 | 0.0002 |
| Allspice | Pimenta officinalis Lindl., berries | 21 ± 4 | 0 | 0.0001 | 30 ± 6 | 25 ± 3 | 0.0005 |
| Myrrh | Commiphora myrrha (T. Nees) Engl. (syn. Balsamea myrrha (T. Nees) Oken), resin | 32 ± 3 | 0 | 0.0001 | 35 ± 6 | 25 ± 2 | 0.0009 |
| Hydacheim | Hedychium spicatum Thymus (syn. Gandasulium spicatum (Buch.-Ham. ex Sm.) Kuntze), flowers | 34 ± 4 | 23 ± 2 | 0.0002 | 38 ± 7 | 26 ± 1 | 0.0017 |
| Litsea cubeba | Litsea cubeba (Lour.) Pers. (syn. Benzoin cubeba (Lour.) Hatus., Persea cubeba (Lour.) Spreng.), fruits | 98 ± 4 | ND d | -- | 77 ± 4 | 27 ± 3 | (0.00004) |
| Palmarosa | Cymbopogon martini var. motia Bruno, grass | 26 ± 5 | 0 | 0.0002 | 35 ± 5 | 29 ± 2 | 0.0004 |
| Lemon eucalyptus | Eucalyptus citriodora Hook. (syn. Corymbia citriodora (Hook.) K.D. Hill & L.A.S. Johnson), leaves | 35 ± 6 | 0 | 0.0006 | 39 ± 7 | 29 ± 4 | 0.0015 |
| Amyris | Amyris balsamifera L. (syn. Elemifera balsamifera (L.) Kuntze), wood | 32 ± 3 | 4 ± 2 | 0.0001 | 38 ± 5 | 29 ± 3 | 0.0006 |
| Cumin | Cuminum cyminum L., seeds | 31 ± 3 | 0 | 0.0001 | 31 ± 6 | 30 ± 1 | 0.0005 |
| Thyme white | Thymus vulgaris L. (syn. Origanum thymus (L.) Kuntze), leaves | 37 ± 2 | 26 ± 2 | 0.0001 | 36 ± 1 | 30 ± 2 | 0.0001 |
| Carrot seed | Daucus carota L., seeds | 38 ± 4 | 5 ± 3 | 0.0004 | 40 ± 3 | 60 ± 2 | 0.0003 (0.0705) |
| Head ease | Synergy blend | 41 ± 3 | 25 ± 3 | 0.0003 | 74 ± 4 | 65 ± 1 | 0.8008 |
| Deep muscle | Synergy blend | 42 ± 4 | 3 ± 2 | 0.0004 | 56 ± 4 | 68 ± 4 | 0.0060 (0.3911) |
| Birch | Betula lenta L., bark | 86 ± 5 | 22 ± 2 | (0.00001) | 91 ± 4 | 69 ± 2 | -- |
| Ho wood | Cinnamomum camphora (L.) J. Presl (syn. Cinnamomum camphora (L.) Nees & Eberm., Cinnamomum camphora (L.) Siebold), twigs and bark | 36 ± 4 | 3 ± 2 | 0.0004 | 69 ± 5 | 70 ± 3 | 0.3078 |
| Petitgrain | Citrus aurantium L, trees and leaves | 38 ± 3 | 19 ± 2 | 0.0002 | 71 ± 4 | 70 ± 3 | 0.4743 |
| Fennel sweet | Foeniculum vulgare Mill., seeds | 40 ± 5 | 2 ± 1 | 0.0006 | 72 ± 3 | 75 ± 4 | 0.6235 |
| Cornmint | Mentha arvensis, leaf | 35 ± 5 | 0 | 0.0004 | 68 ± 4 | 85 ± 1 | 0.1359 |
| Citrus blast | Synergy blend | 51 ± 5 | >70 | 0.0039 | 71 ± 5 | >70 | 0.5865 |
| Nutmeg | Myristica fragrans Houtt., seeds | 43 ± 4 | >70 | 0.0008 | 71 ± 4 | >70 | 0.6533 |
| Alive | Synergy blend | 40 ± 4 | >70 | 0.0004 | 71 ± 3 | >70 | 0.5228 |
| New beginning | Synergy blend | 48 ± 4 | >70 | 0.0013 | 75 ± 4 | >70 | 0.5107 |
| Happy | Synergy blend | 47 ± 4 | >70 | 0.0009 | 78 ± 2 | >70 | -- |
| Meditation | Synergy blend | 55 ± 4 | >70 | 0.0041 | 79 ± 4 | >70 | -- |
| Deep forest | Synergy blend | 61 ± 1 | >70 | 0.0039 | 79 ± 3 | >70 | -- |
| Copaiba | Copaifera officinalis (Jacq.) L., balsm | 51 ± 2 | >70 | 0.0007 | 79 ± 2 | >70 | -- |
| Balsam fir | Abies balsamea (L.) Mill. (syn. Peuce balsamea (L.) Rich.), needles | 57 ± 5 | >70 | 0.0124 | 80 ± 1 | >70 | -- |
| Juniper Berry | Juniperus communis L., berries | 56 ± 5 | >70 | 0.0086 | 82 ± 3 | >70 | -- |
| Camphor | Cinnamomum camphora (L.) J. Presl (syn. Cinnamomum camphora (L.) Nees & Eberm., Cinnamomum camphora (L.) Siebold), wood | 58 ± 3 | >70 | 0.0047 | 82 ± 3 | >70 | -- |
| Vetiver | Vetiveria zizanioides (L.) Nash (syn. Phalaris zizanioides L.), root | 41 ± 3 | >70 | 0.0003 | 82 ± 5 | >70 | -- |
| Fir needle | Abies sibirica Ledeb. (syn. Pinus sibirica (Ledeb.) Turcz.), needles | 60 ± 3 | >70 | 0.0109 | 83 ± 5 | >70 | -- |
| Sleep tight | Synergy blend | 57 ± 5 | >70 | 0.0130 | 85 ± 6 | >70 | -- |
| Turmeric | Curcuma longa L. (syn. Kua domestica (L.) Medik.), root | 50 ± 2 | >70 | 0.0007 | 93 ± 3 | >70 | -- |
| Elemi | Canarium luzonicum (Blume) A. Gray (syn. Pimela luzonica Blume), resin | 58 ± 3 | >70 | 0.0059 | 95 ± 2 | >70 | -- |
| Parsley seed | Petroselinum sativum Hoffm., seeds | 64 ± 5 | >70 | 0.0645 | 97 ± 3 | >70 | -- |
| Essential Oil Treatment | Residual Viability after 0.1% Essential Oil Treatment | Residual Viability after 0.05% Essential Oil Treatment | ||
|---|---|---|---|---|
| Treatment b | Subculture c | Treatment b | Subculture c | |
| Drug free control | 93% | + | 93% | + |
| Daptomycin+Doxycycline+Cefuroxime d | 18%d | − d | N/A | N/A |
| Garlic | 30% | − | 33% | − |
| Allspice | 34% | − | 48% | + |
| Myrrh | 42% | − | 41% | + |
| Hydacheim | 44% | − | 61% | + |
| Litsea cubeba | 68% | − | 69% | + |
| Palmarosa | 39% | + | 67% | + |
| Lemon eucalyptus | 46% | + | 79% | + |
| Amyris | 48% | + | 71% | + |
| Cumin | 42% | + | 60% | + |
| Thyme white | 40% | + | 76% | + |
| Cinnamaldehyde | 34% | − | 56% | − |
| Essential Oil | Components | Content a |
|---|---|---|
| Garlic Allium sativum bulbs | Diallyl disulfide | 19% |
| (E)-1-Allyl-2-(prop-1-en-1-yl) disulfane | 15% | |
| Disulfide, methyl 2-propenyl | 6% | |
| 2-Vinyl-4H-1,3-dithiine | 6% | |
| (E)-methyl 1-propenyl sulfide | 4% | |
| Allspice Pimenta officinalis berries | Eugenol | 82% |
| β-Caryophyllene | 6% | |
| Methyleugenol | 5% | |
| α-Humulene | 1% | |
| α-Selinene | 0.47% | |
| Myrrh Commiphora myrrha resin | Curzerene | 38% |
| Furanoeudesma-1,3-diene | 24% | |
| β-Elemene | 7% | |
| Lindestrene | 7% | |
| E-Elemene | 3% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Shi, W.; Miklossy, J.; Tauxe, G.M.; McMeniman, C.J.; Zhang, Y. Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi. Antibiotics 2018, 7, 89. https://doi.org/10.3390/antibiotics7040089
Feng J, Shi W, Miklossy J, Tauxe GM, McMeniman CJ, Zhang Y. Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi. Antibiotics. 2018; 7(4):89. https://doi.org/10.3390/antibiotics7040089
Chicago/Turabian StyleFeng, Jie, Wanliang Shi, Judith Miklossy, Genevieve M. Tauxe, Conor J. McMeniman, and Ying Zhang. 2018. "Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi" Antibiotics 7, no. 4: 89. https://doi.org/10.3390/antibiotics7040089
APA StyleFeng, J., Shi, W., Miklossy, J., Tauxe, G. M., McMeniman, C. J., & Zhang, Y. (2018). Identification of Essential Oils with Strong Activity against Stationary Phase Borrelia burgdorferi. Antibiotics, 7(4), 89. https://doi.org/10.3390/antibiotics7040089