
Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review


Abstract
:1. Introduction
2. Results
2.1. AMU in Farms
2.2. Antimicrobial Residues in Meat/Fish
2.3. Phenotypic AMR among E. coli and Non-Typhoidal Salmonella (NTS)
2.4. Phenotypic AMR among Campylobacter spp.
2.5. Phenotypic AMR in Bacteria from Aquaculture
2.6. AMR in Bacteria from Terrestrial Animals Other Than E. coli, NTS, and Campylobacter spp.
2.7. Genotypic Studies
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Antimicrobials in Agriculture and the Environment: Reducing Unnecessary Use and Waste. Available online: https://amr-review.org/sites/default/files/Antimicrobials%20in%20agriculture%20and%20the%20environment%20-%20Reducing%20unnecessary%20use%20and%20waste.pdf (accessed on 6 September 2016).
- Da Costa, P.M.; Loureiro, L.; Matos, A.J.F. Transfer of multidrug-resistant bacteria between intermingled ecological niches: The interface between humans, animals and the environment. Int. J. Environ. Res. Public Health 2013, 10, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Joint FAO/WHO/OIE Expert Meeting on Critically Important Antimicrobials; FAO, WHO and OIE: Rome, Italy, 2007. [Google Scholar]
- Critically Important Antimicrobials for Human Medicine 2011. Available online: http://apps.who.int/iris/bitstream/10665/77376/1/9789241504485_eng.pdf (accessed on 30 August 2016).
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.A.; Boete, C.; Binot, A.; By, Y.; Cappelle, J.; Carrique-Mas, J.; Chou, M.; Furey, N.; Kim, S.; Lajaunie, C.; et al. Biodiversity and health: Lessons and recommendations from an interdisciplinary conference to advise Southeast Asian research, society and policy. Infect. Genet. Evol. 2016, 40, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.H.; Custer, B.; Steele, J.A.; Wilcox, B.A.; Xu, J. Intensified food production and correlated risks to human health in the greater mekong subregion: A systematic review. Environ. Health 2015. [Google Scholar] [CrossRef] [PubMed]
- Coker, R.J.; Hunter, B.M.; Rudge, J.W.; Liverani, M.; Hanvoravongchai, P. Emerging infectious diseases in Southeast Asia: Regional challenges to control. Lancet 2011, 377, 599–609. [Google Scholar] [CrossRef]
- von Wintersdorff, C.J.; Penders, J.; Stobberingh, E.E.; Oude Lashof, A.M.; Hoebe, C.J.; Savelkoul, P.H.; Wolffs, P.F. High rates of antimicrobial drug resistance gene acquisition after international travel, the Netherlands. Emerg. Infect. Dis. 2014, 20, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance: Global Report on Surveillance 2014. Available online: http://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 23 August 2016).
- Analysis of Aquaculture Development in Southeast Asia: A Policy Perspective 2009. Available online: http://www.fao.org/docrep/012/i0950e/i0950e00.pdf (accessed on 31 August 2016).
- Livestock Industrialization Project: Phase II—Policy, Technical, and Environmental Determinants and Implications of the Scaling-up of Swine, Broiler, Layer and Milk Production in Thailand. Available online: http://www.fao.org/wairdocs/lead/x6170e/x6170e39.htm (accessed on 23 August 2016).
- Dang, S.T.; Petersen, A.; van Truong, D.; Chu, H.T.; Dalsgaard, A. Impact of medicated feed on the development of antimicrobial resistance in bacteria at integrated pig-fish farms in Vietnam. Appl. Environ. Microbiol. 2011, 77, 4494–4498. [Google Scholar] [CrossRef] [PubMed]
- Carrique-Mas, J.; Trung, N.V.; Hoa, N.T.; Mai, H.H.; Thanh, T.T.; Campbell, J.; Wagenaar, J.; Hardon, A.; Hieu, T.Q.; Schultsz, C. Antimicrobial usage in chicken production in the Mekong Delta of Vietnam. Zoonoses Public Health 2014, 61, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pham, D.K.; Chu, J.; Do, N.T.; Brose, F.; Degand, G.; Delahaut, P.; De Pauw, E.; Douny, C.; Nguyen, K.V.; Vu, T.D.; et al. Monitoring antibiotic use and residue in freshwater aquaculture for domestic use in Vietnam. EcoHealth 2015, 12, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Nguyen, H.M.; Nguyen, C.V.; Nguyen, T.V.; Nguyen, M.T.; Thai, H.Q.; Ho, M.H.; Thwaites, G.; Ngo, H.T.; Baker, S.; et al. Use of colistin and other critical antimicrobials on pig and chicken farms in southern Vietnam and its association with resistance in commensal Escherichia coli bacteria. Appl. Environ. Microbiol. 2016, 82, 3727–3735. [Google Scholar] [CrossRef] [PubMed]
- Cuong, N.; Nhung, N.T.; Nghia, N.H.; Hoa, N.T.M.; Trung, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial consumption in medicated feeds in vietnamese pig and poultry production. EcoHealth 2016. [Google Scholar] [CrossRef]
- Dang, P.K.; Saegerman, C.; Douny, C.; Ton, V.D.; Bo, H.X.; Binh, D.V.; Ngan, P.H.; Scippo, M.L. First survey on the use of antibiotics in pig and poultry production in the red river delta region of Vietnam. Food Public Health 2013, 3, 247–256. [Google Scholar]
- Petersen, A.; Dalsgaard, A. Antimicrobial resistance of intestinal Aeromonas spp. and Enterococcus spp. in fish cultured in integrated broiler-fish farms in Thailand. Aquaculture 2003, 219, 71–82. [Google Scholar] [CrossRef]
- Rico, A.; Phu, T.; Satapornvanit, K.; Min, J.; Shahabuddin, A.; Henriksson, P.; Murray, F.; Little, D.; Dalsgaard, A.; van den Brink, P. Use of veterinary medicines, feed additives and probiotics in four major internationally traded aquaculture species farmed in Asia. Aquaculture 2013, 412, 231–243. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Carrique-Mas, J.J.; Ngo, T.H.; Ho, H.M.; Ha, T.T.; Campbell, J.I.; Nguyen, T.N.; Hoang, N.N.; Pham, V.M.; Wagenaar, J.A.; et al. Prevalence and risk factors for carriage of antimicrobial-resistant Escherichia coli on household and small-scale chicken farms in the Mekong Delta of Vietnam. J. Antimicrob. Chemother. 2015, 70, 2144–2152. [Google Scholar] [PubMed]
- Persoons, D.; Dewulf, J.; Smet, A.; Herman, L.; Heyndrickx, M.; Martel, A.; Catry, B.; Butaye, P.; Haesebrouck, F. Antimicrobial use in belgian broiler production. Preventive Vet. Med. 2012, 105, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Okihashi, M.; Harada, K.; Konishi, Y.; Uchida, K.; Do, M.H.; Bui, H.D.; Nguyen, T.D.; Nguyen, P.D.; Chau, V.V.; et al. Antibiotic residue monitoring results for pork, chicken, and beef samples in Vietnam in 2012–2013. J. Agric. Food Chem. 2015, 63, 5141–5145. [Google Scholar] [CrossRef] [PubMed]
- Van Nhiem, D.; Paulsen, P.; Witaya Suriyasathaporn, W.; Smulders, F.; Kyule, M.; Baumann, M.; Zessin, K.; Ngan, P.H. Preliminary analysis of tetracycline residues in marketed pork in Hanoi, Vietnam. Ann. N. Y. Acad. Sci. 2006, 1081, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Angkititrakul, S.; Chomvarin, C.; Chaita, T.; Kanistanon, K.; Waethewutajarn, S. Epidemiology of antimicrobial resistance in Salmonella isolated from pork, chicken meat and humans in Thailand. Southeast. Asian J. Trop. Med. Public Health 2005, 36, 1510–1515. [Google Scholar] [PubMed]
- Padungtod, P.; Kaneene, J.B. Salmonella in food animals and humans in northern Thailand. Int. J. Food Microbiol. 2006, 108, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Phongpaichit, S.; Liamthong, S.; Mathew, A.G.; Chethanond, U. Prevalence of class 1 integrons in commensal Escherichia coli from pigs and pig farmers in Thailand. J. Food Prot. 2007, 70, 292–299. [Google Scholar] [PubMed]
- Chuanchuen, R.; Pathanasophon, P.; Khemtong, S.; Wannaprasat, W.; Padungtod, P. Susceptibilities to antimicrobials and disinfectants in Salmonella isolates obtained from poultry and swine in Thailand. J. Vet. Med. Sci. 2008, 70, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Bangtrakulnonth, A.; Pulsrikarn, C.; Pornreongwong, S.; Hasman, H.; Song, S.W.; Aarestrup, F.M. Antimicrobial resistance and molecular epidemiology of salmonella rissen from animals, food products, and patients in Thailand and Denmark. Foodborne Pathog. Dis. 2008, 5, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Chuanchuen, R.; Padungtod, P. Antimicrobial resistance genes in Salmonella enterica isolates from poultry and swine in Thailand. J. Vet. Med. Sci. 2009, 71, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Prapasarakul, N.; Tummaruk, P.; Niyomtum, W.; Tripipat, T.; Serichantalergs, O. Virulence genes and antimicrobial susceptibilities of hemolytic and nonhemolytic Escherichia coli isolated from post-weaning piglets in central Thailand. J. Vet. Med. Sci. 2010, 72, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Sanpong, P.; Theeragool, G.; Wajjwalku, W.; Amavisit, P. Characterization of multiple-antimicrobial resistant Salmonella isolated from pig farms in Thailand. Kasetsart J. 2010, 44, 643–651. [Google Scholar]
- Chaisatit, C.; Tribuddharat, C.; Pulsrikarn, C.; Dejsirilert, S. Molecular characterization of antibiotic-resistant bacteria in contaminated chicken meat sold at supermarkets in Bangkok, Thailand. Jpn. J. Infect. Dis. 2012, 65, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Lay, K.K.; Koowattananukul, C.; Chansong, N.; Chuanchuen, R. Antimicrobial resistance, virulence, and phylogenetic characteristics of Escherichia coli isolates from clinically healthy swine. Foodborne Pathog. Dis. 2012, 9, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Chotinun, S.; Rojanasthien, S.; Unger, F.; Tadee, P.; Patchanee, P. Prevalence and antimicrobial resistance of salmonella isolated from carcasses, processing facilities and the environment surrounding small scale poultry slaughterhouses in Thailand. Southeast. Asian J.Trop. Med. Public Health 2014, 45, 1392–1400. [Google Scholar] [PubMed]
- Boonkhot, P.; Tadee, P.; Yamsakul, P.; Pocharoen, C.; Chokesajjawatee, N.; Patchanee, P. Class 1 integrons characterization and multilocus sequence typing of Salmonella spp. from swine production chains in Chiang Mai and Lamphun Provinces, Thailand. Jpn. J. Vet. Res. 2015, 63, 83–94. [Google Scholar] [PubMed]
- Changkaew, K.; Intarapuk, A.; Utrarachkij, F.; Nakajima, C.; Suthienkul, O.; Suzuki, Y. Antimicrobial resistance, extended-spectrum β-lactamase productivity, and class 1 integrons in Escherichia coli from healthy swine. J. Food Prot. 2015, 78, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Love, D.C.; Tharavichitkul, P.; Arjkumpa, O.; Imanishi, M.; Hinjoy, S.; Nelson, K.; Nachman, K.E. Antimicrobial use and multidrug-resistant Salmonella spp., Escherichia coli, and Enterococcus faecalis in swine from northern Thailand. Thai J. Vet. Med. 2015, 45, 43–53. [Google Scholar]
- Saengthongpinit, C.; Kongsoi, S.; Viriyarampa, S.; Songserm, T. Prevalence and antimicrobial resistance of Salmonella and Campylobacter species isolated from laying duck flocks in confinement and free-grazing systems. Thai J. Vet. Med. 2015, 45, 341–350. [Google Scholar]
- Tadee, P.; Boonkhot, P.; Pornruangwong, S.; Patchanee, P. Comparative phenotypic and genotypic characterization of Salmonella spp. in pig farms and slaughterhouses in two provinces in northern Thailand. PLoS ONE 2015, 10, e0116581. [Google Scholar] [CrossRef] [PubMed]
- Changkaew, K.; Utrarachkij, F.; Siripanichgon, K.; Nakajima, C.; Suthienkul, O.; Suzuki, Y. Characterization of antibiotic resistance in Escherichia coli isolated from shrimps and their environment. J. Food Prot. 2014, 77, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Pornsukarom, S.; Patchanee, P.; Erdman, M.; Cray, P.F.; Wittum, T.; Lee, J.; Gebreyes, W.A. Comparative phenotypic and genotypic analyses of Salmonella rissen that originated from food animals in Thailand and United States. Zoonoses Public Health 2015, 62, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, N.; Tran, T.P.; Ly, T.L.; Nguyen, T.T.; Iwata, T.; Okatani, A.T.; Watanabe, M.; Taniguchi, T.; Hirota, Y.; Hayashidani, H. Antimicrobial susceptibilities of Salmonella from domestic animals, food and human in the Mekong Delta, Vietnam. J. Vet. Med. Sci. 2008, 70, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.; Chin, J.; Chapman, T.; Tran, L.T.; Coloe, P.J. Safety of raw meat and shellfish in vietnam: An analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int. J. Food Microbiol. 2008, 124, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Ellerbroek, L.; Narapati, D.; Phu Tai, N.; Poosaran, N.; Pinthong, R.; Sirimalaisuwan, A.; Tshering, P.; Fries, R.; Zessin, K.H.; Baumann, M.; et al. Antibiotic resistance in Salmonella isolates from imported chicken carcasses in bhutan and from pig carcasses in Vietnam. J. Food Prot. 2010, 73, 376–379. [Google Scholar] [PubMed]
- Vo, A.T.T.; van Duijkeren, E.; Gaastra, W.; Fluit, A.C. Antimicrobial resistance, class 1 integrons, and genomic island 1 in Salmonella isolates from Vietnam. PLoS ONE 2010, 5, e9440. [Google Scholar] [CrossRef] [PubMed]
- Thai, T.H.; Hirai, T.; Lan, N.T.; Yamaguchi, R. Antibiotic resistance profiles of Salmonella serovars isolated from retail pork and chicken meat in north Vietnam. Int. J. Microbiol. 2012, 156, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Ta, Y.T.; Nguyen, T.T.; To, P.B.; Pham da, X.; Le, H.T.; Thi, G.N.; Alali, W.Q.; Walls, I.; Doyle, M.P. Quantification, serovars, and antibiotic resistance of Salmonella isolated from retail raw chicken meat in Vietnam. J. Food Prot. 2014, 77, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Nhung, N.T.; Cuong, N.V.; Campbell, J.; Hoa, N.T.; Bryant, J.E.; Truc, V.N.; Kiet, B.T.; Jombart, T.; Trung, N.V.; Hien, V.B.; et al. High levels of antimicrobial resistance among Escherichia coli isolates from livestock farms and synanthropic rats and shrews in the Mekong Delta of Vietnam. Appl. Environ. Microbiol. 2015, 81, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Noor Uddin, G.M.; Larsen, M.H.; Christensen, H.; Aarestrup, F.M.; Phu, T.M.; Dalsgaard, A. Identification and antimicrobial resistance of bacteria isolated from probiotic products used in shrimp culture. PLoS ONE 2015, 10, e0132338. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.T.P.; Hoang, N.V.M.; Cuong, N.V.; Campbell, J.; Bryant, J.E.; Hoa, N.T.; Kiet, B.T.; Thompson, C.; Duy, D.T.; Phat, V.V.; et al. High levels of contamination and antimicrobial resistant non-typhoidal Salmonella serovars on pig and poultry farms in the Mekong Delta of Vietnam. Epidemiol. Infect. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lettini, A.A.; Vo Than, T.; Marafin, E.; Longo, A.; Antonello, K.; Zavagnin, P.; Barco, L.; Mancin, M.; Cibin, V.; Morini, M.; et al. Distribution of Salmonella serovars and antimicrobial susceptibility from poultry and swine farms in central Vietnam. Zoonoses Public Health 2016. [Google Scholar] [CrossRef] [PubMed]
- Trung, N.V.; Carrique-Mas, J.J.; Nghia, N.H.; Tu, L.T.P.; Mai, H.H.; Tuyen, H.T.; Campbell, J.; Nhung, N.T.; Nhung, H.N.; Minh, P.V.; et al. Non-typhoidal salmonella colonization in chickens and humans in the Mekong Delta of Vietnam. Zoonoses Public Health 2016. [Google Scholar] [CrossRef] [PubMed]
- Thung, T.Y.; Mahyudin, N.A.; Basri, D.F.; Wan Mohamed Radzi, C.W.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella enteritidis and Salmonella typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Usui, M.; Ozawa, S.; Onozato, H.; Kuge, R.; Obata, Y.; Uemae, T.; Ngoc, P.T.; Heriyanto, A.; Chalemchaikit, T.; Makita, K.; et al. Antimicrobial susceptibility of indicator bacteria isolated from chickens in Southeast Asian countries (Vietnam, Indonesia and Thailand). J. Vet. Med. Sci. 2014, 76, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Lay, K.S.; Vuthy, Y.; Song, P.; Phol, K.; Sarthou, J.L. Prevalence, numbers and antimicrobial susceptibilities of Salmonella serovars and Campylobacter spp. in retail poultry in Phnom Penh, Cambodia. J. Vet. Med. Sci. 2011, 73, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Boonmar, S.; Markvichitr, K.; Chaunchom, S.; Chanda, C.; Bangtrakulnonth, A.; Pornrunangwong, S.; Yamamoto, S.; Suzuki, D.; Kozawa, K.; Kimura, H.; et al. Salmonella prevalence in slaughtered buffaloes and pigs and antimicrobial susceptibility of isolates in Vientiane, Lao People’s Democratic Republic. J. Vet. Med. Sci. 2008, 70, 1345–1348. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Lertworapreecha, M.; Evans, M.C.; Bangtrakulnonth, A.; Chalermchaikit, T.; Hendriksen, R.S.; Wegener, H.C. Antimicrobial susceptibility and occurrence of resistance genes among Salmonella enterica serovar weltevreden from different countries. J. Antimicrob. Chemother. 2003, 52, 715–718. [Google Scholar] [CrossRef] [PubMed]
- The Prevalence and Antimicrobial Resistance of Campylobacter Species Isolated from Intestines of Chicken in Retail Markets of Thailand. Available online: http://www.vet.chula.ac.th/vet2014/Ebook/ebook/Proceedings/Symposium%2011th/CD_Proceedings/P110-P111%20p116.pdf (accessed on 26 August 2016).
- Boonmar, S.; Sangsuk, L.; Suthivarakom, K.; Padungtod, P.; Morita, Y. Serotypes and antimicrobial resistance of Campylobacter jejuni isolated from humans and animals in Thailand. Southeast. Asian J. Trop. Med. Public Health 2005, 36, 130–134. [Google Scholar] [PubMed]
- Sukhapesna, J.; Amavisit, P.; Wajjwalku, W.; Thamchaipenet, A.; Sukpuaram, T. Antimicrobial resistance of Campylobacter jejuni isolated from chicken in Nakhon Pathom Province, Thailand. Kasetsart J. 2005, 39, 240–246. [Google Scholar]
- Padungtod, P.; Kaneene, J.B.; Hanson, R.; Morita, Y.; Boonmar, S. Antimicrobial resistance in Campylobacter isolated from food animals and humans in northern Thailand. FEMS Immunol. Med. Microbiol. 2006, 47, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Noppon, B.; Sthitmatee, N.; Asai, T.; Kataoka, Y.; Sawada, T. Isolation and antimicrobial resistance of Campylobacter spp. From chicken faecal samples in Khon Kaen and nearby province of Thailand. Chiang Mai Vet. J. 2009, 7, 115–123. [Google Scholar]
- Garin, B.; Gouali, M.; Wouafo, M.; Perchec, A.M.; Pham, M.T.; Ravaonindrina, N.; Urbes, F.; Gay, M.; Diawara, A.; Leclercq, A.; et al. Prevalence, quantification and antimicrobial resistance of Campylobacter spp. on chicken neck-skins at points of slaughter in 5 major cities located on 4 continents. Int. J. Food Microbiol. 2012, 157, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Chokboonmongkol, C.; Patchanee, P.; Golz, G.; Zessin, K.H.; Alter, T. Prevalence, quantitative load, and antimicrobial resistance of Campylobacter spp. from broiler ceca and broiler skin samples in Thailand. Poult. Sci. 2013, 92, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Carrique-Mas, J.J.; Bryant, J.E.; Cuong, N.V.; Hoang, N.V.; Campbell, J.; Hoang, N.V.; Dung, T.T.; Duy, D.T.; Hoa, N.T.; Thompson, C.; et al. An epidemiological investigation of Campylobacter in pig and poultry farms in the Mekong Delta of Vietnam. Epidemiol. Infect. 2014, 142, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Sison, F.B.; Chaisowwong, W.; Alter, T.; Tiwananthagorn, S.; Pichpol, D.; Lampang, K.N.; Baumann, M.P.; Golz, G. Loads and antimicrobial resistance of Campylobacter spp. on fresh chicken meat in Nueva Ecija, Philippines. Poul. Sci. 2014, 93, 1270–1273. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.M.; Hotzel, H.; El-Adawy, H.; Tran, H.T.; Le, M.T.H.; Tomaso, H.; Neubauer, H.; Hafez, H.M. Genotyping and antibiotic resistance of thermophilic Campylobacter isolated from chicken and pig meat in Vietnam. Gut Pathog. 2016. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchaikul, N.; Garelick, H.; Jones, H.; Jacobs, J.; Ball, D.; Choudhury, M. Resistance to three antimicrobial agents of Campylobacter isolated from organically- and intensively-reared chickens purchased from retail outlets. Int. J. Antimicrob. Agents 2006, 27, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Mansouri-najand, L.; Saleha, A.A.; Wai, S.S. Prevalence of multidrug resistance Campylobacter jejuni and Campylobacter coli in chickens slaughtered in selected markets, Malaysia. Trop. Biomed. 2012, 29, 231–238. [Google Scholar] [PubMed]
- Schwan, P. Prevalence and Antibiotic Resistance of Campylobacter spp. in Poultry and Raw Meat in the Can tho Province, Vietnam. Available online: http://stud.epsilon.slu.se/1066/1/Schwan_P_20100419.pdf (accessed on 15 August 2016).
- Saleha, A.A. Isolation and characterization of Campylobacter jejuni from broiler chickens in Malaysia. Int. J. Poult. Sci. 2002, 1, 94–97. [Google Scholar]
- Ekkapobyotin, C.; Padungtod, P.; Chuanchuen, R. Antimicrobial resistance of Campylobacter coli isolates from swine. Int. J. Food Microbiol. 2008, 128, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchaikul, N.; Garelick, H. Antimicrobial resistance of Campylobacter species isolated from edible bivalve molluscs purchased from Bangkok markets, Thailand. Foodborne Pathog. Dis. 2009, 6, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.D.; Haesebrouck, F.; Nguyen, A.T.; Sorgeloos, P.; Baele, M.; Decostere, A. Antimicrobial susceptibility pattern of Edwardsiella ictaluri isolates from natural outbreaks of bacillary necrosis of pangasianodon hypophthalmus in Vietnam. Microb. Drug Resist. 2008, 14, 311–316. [Google Scholar] [PubMed]
- Abdelsalam, M.; Chen, S.C.; Yoshida, T. Phenotypic and genetic characterizations of Streptococcus dysgalactiae strains isolated from fish collected in Japan and other Asian countries. FEMS Microbiol. Lett. 2010, 302, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Khan, S.A.; Tran, Q.; Sung, K.; Khan, A.A.; Adamu, I.; Steele, R.S. Isolation and characterization of multidrug-resistant Klebsiella spp. isolated from shrimp imported from Thailand. Inte. J. Food Microbiol. 2012, 155, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Rogge, M.L.; Dubytska, L.; Jung, T.S.; Wiles, J.; Elkamel, A.A.; Rennhoff, A.; Oanh, D.T.; Thune, R.L. Comparison of Vietnamese and us isolates of Edwardsiella ictaluri. Dis. Aquat. Org. 2013, 106, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.N.; Van, T.T.; Nguyen, H.T.; Smooker, P.M.; Shimeta, J.; Coloe, P.J. Molecular characterization of antibiotic resistance in Pseudomonas and Aeromonas isolates from catfish of the Mekong Delta, Vietnam. Vet. Microbiol. 2014, 171, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.; Wang, R.; Wiles, J.; Baumgartner, W.; Green, C.; Plumb, J.; Hawke, J. Characterization of isolates of Streptococcus agalactiae from diseased farmed and wild marine fish from the U.S. Gulf Coast, Latin America, and Thailand. J. Aquat. Anim. Health 2015, 27, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Dangwetngam, M.; Suanyuk, N.; Kong, F.; Phromkunthong, W. Serotype distribution and antimicrobial susceptibilities of Streptococcus agalactiae isolated from infected cultured tilapia (Oreochromis niloticus) in Thailand: Nine-year perspective. J. Med. Microbiol. 2016, 65, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Getachew, Y.M.; Hassan, L.; Zakaria, Z.; Saleha, A.A.; Kamaruddin, M.I.; Che Zalina, M.Z. Characterization of vancomycin-resistant Enterococcus isolates from broilers in Selangor, Malaysia. Trop. Biomed. 2009, 26, 280–288. [Google Scholar] [PubMed]
- Shah-Majid, M.; Maria, A.R.; Shahidayani, S.; Salwani, A.M.; Khairani, S. Occurrence of vancomycin-resistant Enterococci in chickens in Malaysia. Vet. Rec. 2007, 160, 702–703. [Google Scholar] [CrossRef] [PubMed]
- Fifadara, N.; Radu, S.; Hassan, Z.; Beuchat, L.R.; Rusul, G. Hemolytic and nonhemolytic vancomycin-resistant Enterococcus faecalis isolated from beef imported to Malaysia. J. Food Prot. 2003, 66, 1845–1850. [Google Scholar] [PubMed]
- Toosa, H.; Radu, S.; Rusul, G.; Latif, A.R.A.; Rahim, R.A.; Ahmad, N.; Ling, O.W. Detection of vancomycin-resistant Enterococcus spp. (VRE) from poultry. Malaysian J. Med. Sci. 2001, 8, 53–58. [Google Scholar]
- Pruksakorn, C.; Pimarn, C.; Boonsoongnern, A.; Narongsak, W. Detection and phenotypic characterization of vancomycin-resistant Enterococci in pigs in Thailand. Agric. Nat. Resour. 2016, 50, 199–203. [Google Scholar] [CrossRef]
- Larsen, J.; Imanishi, M.; Hinjoy, S.; Tharavichitkul, P.; Duangsong, K.; Davis, M.F.; Nelson, K.E.; Larsen, A.R.; Skov, R.L. Methicillin-resistant Staphylococcus aureus ST9 in pigs in Thailand. PLoS ONE 2012, 7, e31245. [Google Scholar] [CrossRef] [PubMed]
- Anukool, U.; O′Neill, C.E.; Butr-Indr, B.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M. Meticillin-resistant Staphylococcus aureus in pigs from Thailand. Int. J. Antimicrob. Agents 2011, 38, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Cavaco, L.; Sirichote, P.; Unahalekhakac, A.; Dangsakul, W.; Svendsen, C.A.; Aarestrup, F.M.; Hendriksen, R. SCCmec type IX element in methicillin resistant Staphylococcus aureus spa type t337 (CC9) isolated from pigs and pork in Thailand. Front. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Sinlapasorn, S.; Lulitanond, A.; Angkititrakul, S.; Chanawong, A.; Wilailuckana, C.; Tavichakorntrakool, R.; Chindawong, K.; Seelaget, C.; Krasaesom, M.; Chartchai, S.; et al. Sccmec IX in meticillin-resistant Staphylococcus aureus and meticillin-resistant coagulase-negative staphylococci from pigs and workers at pig farms in Khon Kaen, Thailand. J. Med. Microbiol. 2015, 64, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Patchanee, P.; Tadee, P.; Arjkumpa, O.; Love, D.; Chanachai, K.; Alter, T.; Hinjoy, S.; Tharavichitkul, P. Occurrence and characterization of livestock-associated methicillin-resistant Staphylococcus aureus in pig industries of northern Thailand. J. Vet. Sci. 2014, 15, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.H.; Tran, T.B.; Tran, T.T.; Nguyen, V.D.; Campbell, J.; Pham, H.A.; Huynh, H.T.; Nguyen, V.V.; Bryant, J.E.; Tran, T.H.; et al. Slaughterhouse pigs are a major reservoir of Streptococcus suis serotype 2 capable of causing human infection in southern Vietnam. PLoS ONE 2011, 6, e17943. [Google Scholar]
- Chukiatsiri, K.; Sasipreeyajan, J.; Blackall, P.J.; Yuwatanichsampan, S.; Chansiripornchai, N. Serovar identification, antimicrobial sensitivity, and virulence of Avibacterium paragallinarum isolated from chickens in Thailand. Avian Dis. 2012, 56, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Pakpinyo, S.; Sasipreeyajan, J. Molecular characterization and determination of antimicrobial resistance of Mycoplasma gallisepticum isolated from chickens. Vet. Microbiol. 2007, 125, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Poernomo, S.; Sutarma; Rafiee, M.; Blackall, P.J. Characterisation of isolates of Haemophilus paragallinarum from Indonesia. Aust. Vet. J. 2000, 78, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Radua, S.; Ling, O.W.; Srimontree, S.; Lulitanond, A.; Hin, W.F.; Yuherman; Lihan, S.; Rusul, G.; Mutalib, A.R. Characterization of Burkholderia pseudomallei isolated in Thailand and Malaysia. Diagn. Microbiol. Infect. Dis. 2000, 38, 141–145. [Google Scholar] [CrossRef]
- Lee, S.H.; Chong, C.E.; Lim, B.S.; Chai, S.J.; Sam, K.K.; Mohamed, R.; Nathan, S. Burkholderia pseudomallei animal and human isolates from Malaysia exhibit different phenotypic characteristics. Diagn. Microbiol. Infect. Dis. 2007, 58, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Ngamwongsatit, B.; Tanomsridachchai, W.; Suthienkul, O.; Urairong, S.; Navasakuljinda, W.; Janvilisri, T. Multidrug resistance in Clostridium perfringens isolated from diarrheal neonatal piglets in Thailand. Anaerobe 2016, 38, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.; Moutafis, G.; Istivan, T.; Tran, L.T.; Coloe, P.J. Detection of Salmonella spp. in retail raw food samples from Vietnam and characterization of their antibiotic resistance. Appl. Environ. Microbiol. 2007, 73, 6885–6890. [Google Scholar] [CrossRef] [PubMed]
- Chuanchuen, R.; Koowatananukul, C.; Khemtong, S. Characterization of class 1 integrons with unusual 3’ conserved region from Salmonella enterica isolates. Southeast. Asian J. Trop. Med. Public Health 2008, 39, 419–424. [Google Scholar] [PubMed]
- Khemtong, S.; Chuanchuen, R. Class 1 integrons and Salmonella genomic island 1 among Salmonella enterica isolated from poultry and swine. Microb. Drug Resistance 2008, 14, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Benacer, D.; Thong, K.L.; Watanabe, H.; Puthucheary, S.D. Characterization of drug resistant Salmonella enterica serotype Typhimurium by antibiograms, plasmids, integrons, resistance genes and PFGE. J. Microbiol. Biotec. 2010, 20, 1042–1052. [Google Scholar]
- Le, Q.P.; Ueda, S.; Nguyen, T.N.; Dao, T.V.; van Hoang, T.A.; Tran, T.T.; Hirai, I.; Nakayama, T.; Kawahara, R.; Do, T.H.; et al. Characteristics of extended-spectrum β-lactamase-producing Escherichia coli in retail meats and shrimp at a local market in Vietnam. Foodborne Pathog. Dis. 2015, 12, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Malhotra-Kumar, S.; Xavier, B.B.; Das, A.J.; Lammens, C.; Hoang, H.T.; Pham, N.T.; Goossens, H. Colistin-resistant Escherichia coli harbouring mcr-1 isolated from food animals in Hanoi, Vietnam. Lancet Infect. Dis. 2016, 16, 286–287. [Google Scholar] [CrossRef]
- Cavaco, L.M.; Hendriksen, R.S.; Aarestrup, F.M. Plasmid-mediated quinolone resistance determinant qnrS1 detected in Salmonella enterica serovar corvallis strains isolated in Denmark and Thailand. J. Antimicrob. Chemother. 2007, 60, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Hoa, N.T.; Chieu, T.T.; Nghia, H.D.; Mai, N.T.; Anh, P.H.; Wolbers, M.; Baker, S.; Campbell, J.I.; Chau, N.V.; Hien, T.T.; et al. The antimicrobial resistance patterns and associated determinants in Streptococcus suis isolated from humans in southern Vietnam, 1997–2008. BMC Infect. Dis. 2011. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Dalsgaard, A. Species composition and antimicrobial resistance genes of Enterococcus spp, isolated from integrated and traditional fish farms in Thailand. Environ. Microbiol. 2003, 5, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Chabou, S.; Okdah, L.; Morand, S.; Rolain, J.M. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 2016. [Google Scholar] [CrossRef]
- Wannaprasat, W.; Padungtod, P.; Chuanchuen, R. Class 1 integrons and virulence genes in Salmonella enterica isolates from pork and humans. Int. J. Antimicrob. Agents 2011, 37, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.; Moutafis, G.; Tran, L.T.; Coloe, P.J. Antibiotic resistance in food-borne bacterial contaminants in Vietnam. Appl. Environ. Microbiol. 2007, 73, 7906–7911. [Google Scholar] [CrossRef] [PubMed]
- Bodhidatta, L.; Vithayasai, N.; Eimpokalarp, B.; Pitarangsi, C.; Serichantalergs, O.; Isenbarger, D.W. Bacterial enteric pathogens in children with acute dysentery in Thailand: Increasing importance of quinolone-resistant Campylobacter. Southeast. Asian J. Trop. Med. Public Health 2002, 33, 752–757. [Google Scholar] [PubMed]
- Carrique-Mas, J.J.; Bryant, J.E. A review of foodborne bacterial and parasitic zoonoses in Vietnam. EcoHealth 2013. [Google Scholar] [CrossRef] [PubMed]
- D’Aoust, J.Y. Salmonella and the international food trade. Int. J. Food Microbiol. 1994, 24, 11–31. [Google Scholar] [CrossRef]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well known antibiotics on gram-positive bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Hombach, M.; Bottger, E.C.; Roos, M. The critical influence of the intermediate category on interpretation errors in revised EUCAST and CLSI antimicrobial susceptibility testing guidelines. Clin. Microbiol. Infect. 2013, 19, E59–E71. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2013. EFSA J. 2015. [Google Scholar] [CrossRef]
- Trung, N.V.; Carrique-Mas, J.J.; van Rooijen, W.; Nhung, N.T.; Tuyen, H.T.; Nhung, H.N.; Minh, P.V.; Mai, H.H.; Hieu, T.Q.; Campbell, J.; et al. Contribution of non-intensive chicken farming to extended-spectrum beta-lactamase producing Escherichia coli colonization in humans in southern Vietnam. Antimicrob. Agents Chemother. 2016, in press. [Google Scholar]
- Boonyasiri, A.; Tangkoskul, T.; Seenama, C.; Saiyarin, J.; Tiengrim, S.; Thamlikitkul, V. Prevalence of antibiotic resistant bacteria in healthy adults, foods, food animals, and the environment in selected areas in Thailand. Pathog. Global Health 2014, 108, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Stoesser, N.; Mathers, A.J.; Moore, C.E.; Day, N.P.; Crook, D.W. Colistin resistance gene mcr-1 and pHNSHP45 plasmid in human isolates of Escherichia coli and Klebsiella pneumoniae. Lancet Infect. Dis. 2016, 16, 285–286. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Thongmalayvong, B.; Akkhavong, K.; Somphavong, S.; Paboriboune, P.; Khounsy, S.; Morand, S.; Rolain, J.M. Clonal transmission of a colistin-resistant Escherichia coli from a domesticated pig to a human in Laos. J. Antimicrob. Chemother. 2015, 70, 3402–3404. [Google Scholar] [PubMed]
- Arcilla, M.S.; van Hattem, J.M.; Matamoros, S.; Melles, D.C.; Penders, J.; de Jong, M.D.; Schultsz, C.; COMBAT consortium. Dissemination of the mcr-1 colistin resistance gene. Lancet. Infect. Dis. 2016, 16, 147–149. [Google Scholar] [CrossRef]
- Van, T.T.; Nguyen, H.N.; Smooker, P.M.; Coloe, P.J. The antibiotic resistance characteristics of non-typhoidal Salmonella enterica isolated from food-producing animals, retail meat and humans in Southeast Asia. Int. J. Food Microbiol. 2012, 154, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D. Impact of antibiotic use in the swine industry. Curr. Opin. Microbiol. 2014, 19, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bogaard, A.E.; London, N.; Stobberingh, E.E. Antimicrobial resistance in pig faecal samples from the Netherlands (five abattoirs) and Sweden. J. Antimicrob. Chemother. 2000, 45, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O. Vancomycin resistant enterococci in farm animals—Occurrence and importance. Infect. Ecol. Epidemiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- The State of World Fisheries and Aquaculture 2012. Available online: http://www.fao.org/docrep/016/i2727e/i2727e00.htm (accessed on 1 August 2016).
- Suzuki, S.; Hoa, P.T. Distribution of quinolones, sulfonamides, tetracyclines in aquatic environment and antibiotic resistance in Indochina. Front. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The european union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2014. EFSA J. 2016. [Google Scholar] [CrossRef]
- Thamlikitkul, V.; Rattanaumpawan, P.; Boonyasiri, A.; Pumsuwan, V.; Judaeng, T.; Tiengrim, S.; Paveenkittiporn, W.; Rojanasthien, S.; Jaroenpoj, S.; Issaracharnvanich, S. Thailand antimicrobial resistance containment and prevention program. J. Global Antimicrob. Resist. 2015, 3, 290–294. [Google Scholar] [CrossRef]
- Archawakulathep, A.; Thi Kim, C.T.; Meunsene, D.; Handijatno, D.; Hassim, H.; Rovira, H.R.; Myint, K.S.; Baldrias, L.R.; Sothy, M.; Aung, M.; et al. Perspectives on antimicrobial resistance in livestock and livestock products in Asean countries. Thai J. Vet. Med. 2014, 44, 5–13. [Google Scholar]
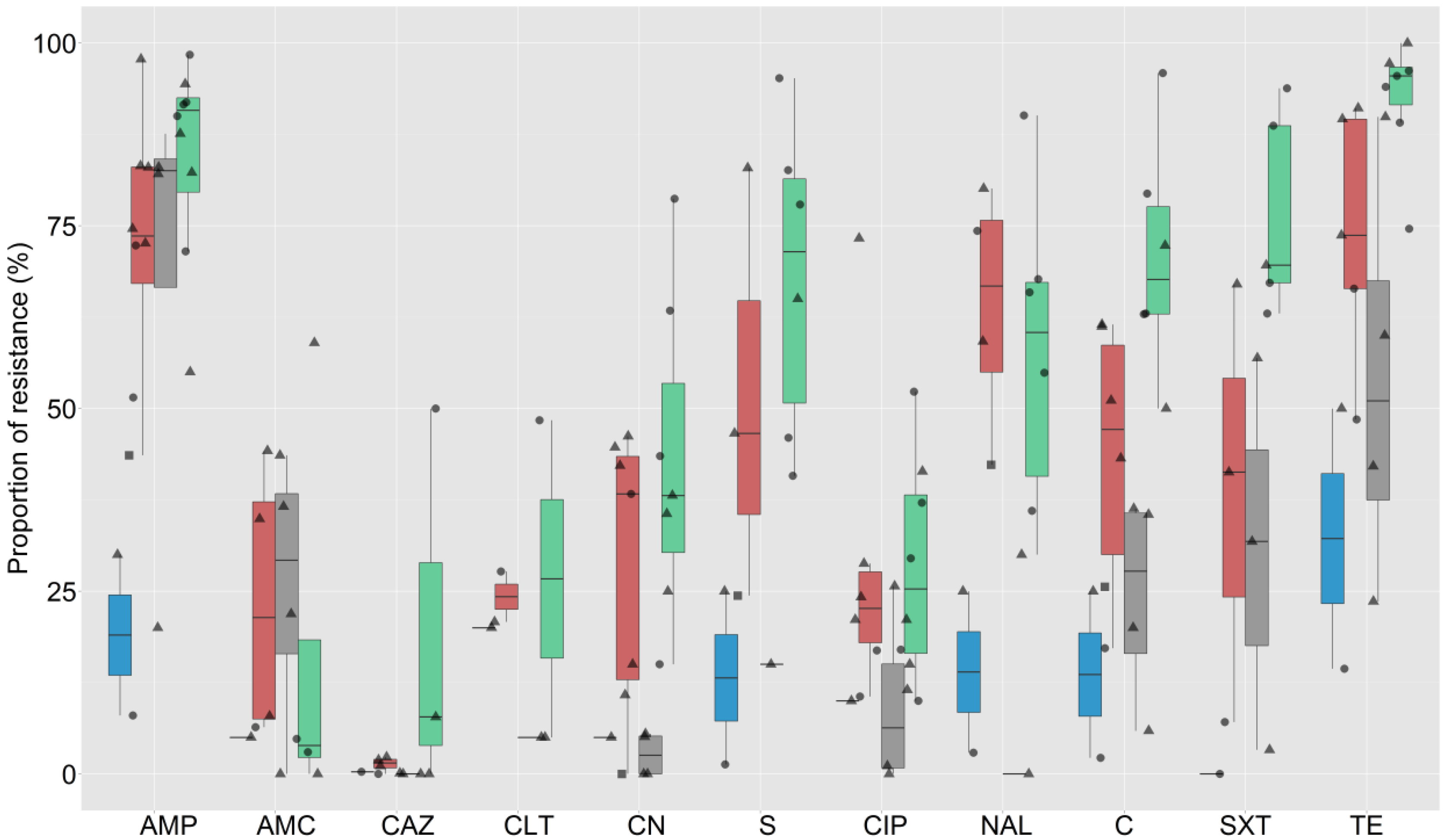
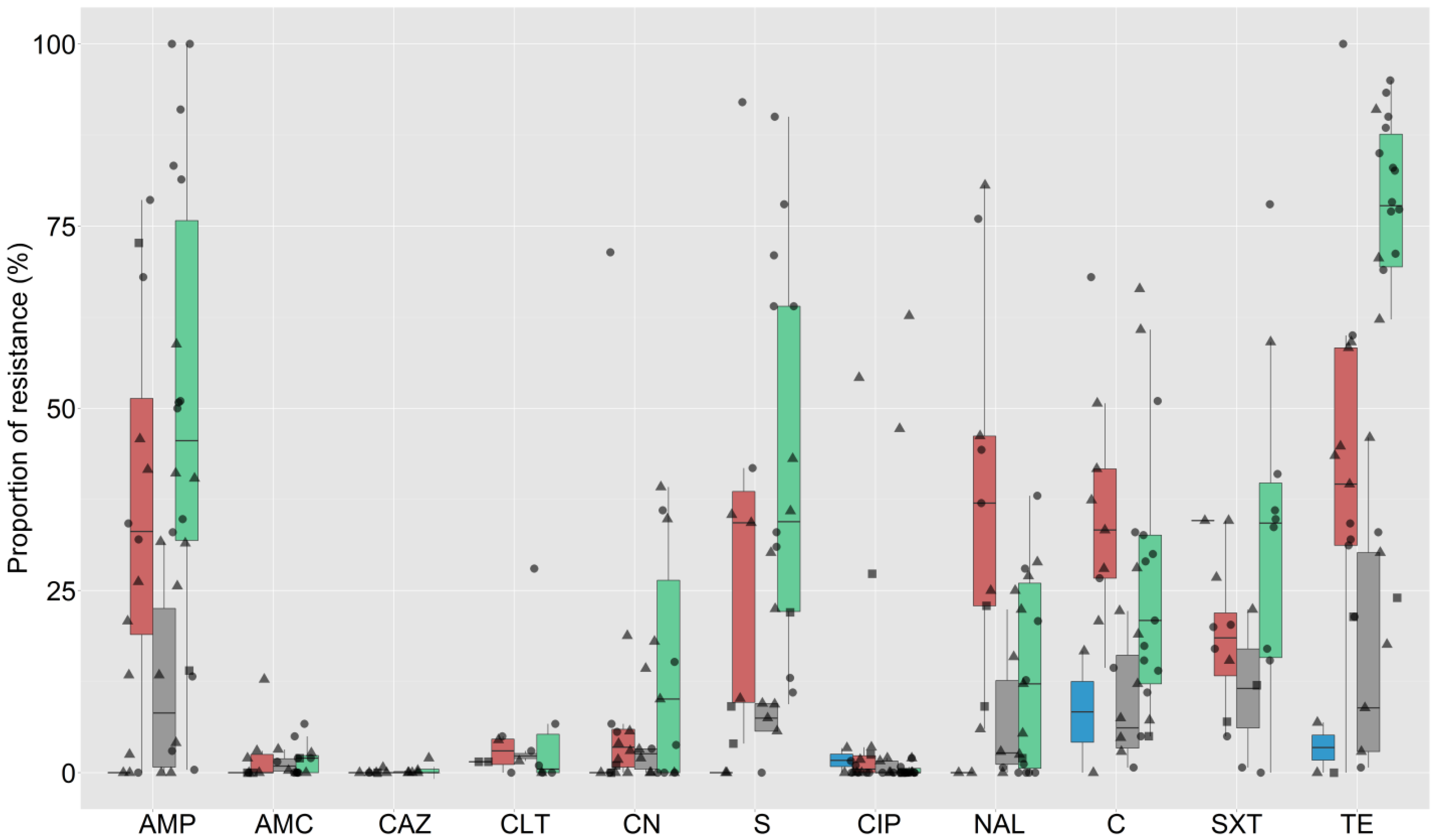

{kind=link}
{kind=link}
| Variable | Escherichia coli | Non-typhoidal Salmonella | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AMP (3294) § | CN (2667) | C (2799) | TE (3231) | CIP (2857) | AMP (3849) | CN (3144) | C (3339) | TE (3352) | CIP (4026) | |
| Host type (baseline = Chicken) | ||||||||||
| Aquaculture | 0.03 *** | 0.16 | 0.04 *** | 0.12 *** | 0.71 | NC | NC | 0.15 ** | 0.05 *** | 0.73 |
| Pig | 2.33 *** | 2.35 *** | 3.26 *** | 6.65 *** | 1.45 ** | 2.86 *** | 3.98 *** | 1.46 *** | 3.89 *** | 3.19 *** |
| Other a | 0.93 | 0.17 *** | 0.29 *** | 0.17 *** | 0.43 *** | 0.32 *** | 0.54 * | 0.31 *** | 0.21 *** | 0.33 ** |
| Country (baseline = Other b) | ||||||||||
| Thailand | 1.75 * | 1 (baseline) | 1.98 | 1 (baseline) | 1 (baseline) | 2.52 *** | 8.16 ** | 7.55 *** | 4.12 *** | 0.02 *** |
| Vietnam | 2.19 ** | 0.44 *** | 2.87 ** | 5.31 *** | 0.91 | 1.18 | 5.40 * | 11.10 *** | 4.10 *** | 0.41 |
| Period (baseline = 2007 or earlier) | ||||||||||
| 2008–2016 | 2.93 *** | 0.68 * | 1.39 * | 0.82 | 2.46 *** | 3.73 *** | 1.12 | 1.39 *** | 1.38 ** | 9.59 *** |
| Variable | Total No. Estimates | AMP | CN | C | TE | NAL | CIP | ERY | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n* | median | n* | median | n* | median | n* | median | n* | median | n* | median | n* | median | |||
| Host species | Pig | 5 | 1 | 51.2 | 4 | 10.1 | 4 | 2.0 | 4 | 84.2 | 2 | 89.6 | 5 | 84.5 | 5 | 66.0 |
| Poultry | 19 | 10 | 37.1 | 14 | 11.0 | 9 | 2.2 | 13 | 71.0 | 11 | 83.3 | 17 | 66.6 | 17 | 23.7 | |
| Country | Thailand | 15 | 5 | 31.2 | 9 | 0.0 | 8 | 0.0 | 12 | 77.2 | 7 | 79.6 | 15 | 81.2 | 13 | 50.5 |
| Vietnam | 6 | 4 | 57.9 | 5 | 16.2 | 4 | 9.9 | 2 | 73.0 | 6 | 92.0 | 6 | 44.5 | 6 | 62.5 | |
| Others** | 4 | 2 | 55.7 | 4 | 18.8 | 2 | 11.2 | 3 | 80.9 | 1 | 58.3 | 2 | 48.1 | 4 | 21.9 | |
| Total | 25 | 11 | 40.0 | 18 | 11.0 | 14 | 2.0 | 17 | 76.6 | 14 | 83.6 | 23 | 70.4 | 23 | 46.2 | |
| Ref. | Country | Species | Host Species | Location | No. Isolates | Prevalence of AMR |
|---|---|---|---|---|---|---|
| [20] | Thailand | Aeromonas spp. | Fish | Chicken-fish farm | 27 | OTC (37%), SMX (19%), ERY (11%), C (7%), CIP (4%) |
| [20] | Thailand | Aeromonas spp. | Fish | Fish farm (no chicken manure) | 45 | S (22%), OTC (13%), ERY (2%), C (2%), CIP (0%) |
| [20] | Thailand | Enterococcus spp. | Fish | Fish farm using chicken manure | 97 | S (72%), OTC (75%), ERY (91%), C (8%), CIP (15%) |
| [20] | Thailand | Enterococcus spp. | Fish | Fish farm (no chicken manure) | 69 | S (31%), ERY (23%), OTC (16%), CIP (6%), C (0%) |
| [76] | Vietnam | Edwardsiella ictaluri | Fish | Diseased fish | 64 | S (83%), OTC (81%), TMP (73.4%), FLU (8%), OA (6%), ENR (5%), C (0%), NIT (0%), AMX (0%), AMC (0%), KA (0%), CN(0%), NEO (0%), FFC (0%) |
| [77] | Malaysia and Indonesia | Streptococcus dysgalactiae | Fish | Diseased fish | 4 | OTC (100%), AMP (0%), ERY (0%), FFL (0%), LCM (0%) |
| [78] | Thailand | Klebsiella spp. | Shrimp | Market | 67 | AMP (100%), TE (100%), BAC (100%), CLI (100%), S (47%), SXT (47%), C (47%), RIF (47%), NAL (12%) |
| [79] | Vietnam | Edwardsiella ictaluri | Fish | Diseased fish | 19 | SXT (89%), FFC (47%), C (47%), OTC (31%), NIT (0%) |
| [80] | Vietnam | Pseudomonas spp. | Fish | Fish farm | 116 | AMP (99%), SXT (93%), NIT (90%), NAL (93%), C (89%), TE (30%), S (28%), DOX (25%), CN (16%), KA (12%), NOR (9%), CIP (9%), NEO (3%) |
| [80] | Vietnam | Aeromonas spp. | Fish | Fish farm | 92 | AMP (94%), SXT (61%), NAL (52%), TE (34%), C (31%), S (31%), NIT (25%), DOX (15%), KA (12%), CIP (8%), CN (6%), NEO (5%), NOR (4%) |
| [81] | Thailand | Streptococcus agalactiae | Fish | Diseased fish | 4 | AMP (100%), CN (100%), C (100%), ENR (100%), OXA (100%), NIT (100%), PEN (75%), FFC (75%), SXT (50%), ERY (50%) |
| [82] | Thailand | Streptococcus agalactiae | Fish | Diseased fish | 144 | OA (100%), CN (100%), SMX (100%), TMP (93%), OTC (12%), NOR (2%), LCM (1%), AMP (0%), C (0%), ERY (0%) |
| Antimicrobial Class | E. coli | NTS | Others | |||
|---|---|---|---|---|---|---|
| n | Gene(s) Detected | n | Gene(s) Detected | n | Gene(s) Detected and Host Bacteria | |
| Tetracycline | 4 | tetA, tetB, tetC | 5 | tetA, tetB, tetC, tetG | 11 | tetA (AC, AP), tetB (KL, AC, AP), tetD (KL), tetL (EN, SS), tetM (AP, SD, EN, SS, MRSA), tetO (CA, SS), tetS (EN), tet39 (AC) |
| Quilonone | 2 | gyrA, parC | 3 | gyrA, gyrB, parC, parE, | 4 | gyrA (CA, KL), gyrB (CA, KL), parC (KL) |
| 4 | acc(6)-Ib, qnrA, qnrS, OqxA | 1 | qnrS | |||
| Diaminopyrimidine | 7 | dfrA1, dfrA5, dfrA10, dfrA12, dfrA17, dhfr17, dhfrI, dhfrV, dhfrXII | 8 | dfrA1, dfrA7, dfrA10, dfrA12, dfrA17, dhfrXI | 1 | dfrA1, dfrA12, dfrA21 (PS, AE) |
| β-lactam | 5 | blaSHV, blaTEM, blaCTX-M | 10 | blaTEM, blaPSE-1, blaOXA-1, blaOXA-30 | 2 | blaROB-1 (AP), blaZ (MRSA) |
| 1 | MOXM, DHAM | 1 | temA, temB | |||
| Aminoglycoside | 8 | aadA1, aadA2,aadA3, aadA4, aadA5, aadA22, aadA23, aadB | 10 | aadA1, aadA2, aadA4, aadA5, aadA22, aadB | 2 | aadA1 (PS, AE), addA2 (PS, AE), aadA9 (CA), aadD (MRSA) |
| 2 | strA, strB | 3 | strA, strB | |||
| 3 | aac(3)-IV, aacA4, aphA1, aph-(3′)-IA | 3 | aac(3)-IV, aphA1-1AB, aphA2 | 2 | aac-6′-aph2″ (EN), acc-aphD (MRSA) | |
| Sulphonamide | 5 | sul1, sul2, sul3 | 4 | sul1, sul2, sul3 | 2 | sul1 (CA, AC), sul2 (AC) |
| Phenicol | 4 | catA, catB, cmlA | 3 | catA, catB, cmlA, florR | 3 | catpIP501 (EN), catB8 (PS, AE), cat, fexA (MRSA) |
| Polymixin | 3 | mcr-1 | ||||
| Lincosamide | 2 | linF, lnuF | ||||
| Macrolide | 7 | 23S rRNA (CA), ermA (AP, EN), ermB (AP, EN, SS, MRSA), ermD (BA) | ||||
| Polypeptide | 2 | vanA, vanC1, vanC2/3 (EN) | ||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nhung, N.T.; Cuong, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics 2016, 5, 37. https://doi.org/10.3390/antibiotics5040037
Nhung NT, Cuong NV, Thwaites G, Carrique-Mas J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics. 2016; 5(4):37. https://doi.org/10.3390/antibiotics5040037
Chicago/Turabian StyleNhung, Nguyen T., Nguyen V. Cuong, Guy Thwaites, and Juan Carrique-Mas. 2016. "Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review" Antibiotics 5, no. 4: 37. https://doi.org/10.3390/antibiotics5040037
APA StyleNhung, N. T., Cuong, N. V., Thwaites, G., & Carrique-Mas, J. (2016). Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics, 5(4), 37. https://doi.org/10.3390/antibiotics5040037