Abstract
Bovine mastitis, particularly that caused by Staphylococcus aureus, presents a major challenge to dairy production worldwide due to its economic impact, animal welfare concerns, and zoonotic potential. This narrative review synthesizes current literature on the epidemiology, pathogenesis, resistance patterns, and control strategies related to S. aureus-associated mastitis in dairy cattle. It highlights the pathogen’s virulence mechanisms, such as biofilm formation, immune evasion, and toxin production, that facilitate persistent infections. The review compiles global prevalence data, revealing significant geographic variation and disparities between clinical and subclinical cases. Antimicrobial resistance, especially the emergence of methicillin-resistant S. aureus (MRSA), is extensively examined alongside resistance gene profiles. Diagnostic approaches, including culture, PCR, MALDI-TOF MS, and AI-based systems, are evaluated for their sensitivity and field applicability. Additionally, the review addresses public health implications, zoonotic risks, and One Health perspectives, culminating in an exploration of prevention strategies, including improved hygiene, vaccination, dry cow therapy, and AI-driven herd management. The findings emphasize the urgent need for integrated surveillance, precision diagnostics, and targeted interventions to mitigate the burden of S. aureus mastitis.
1. Introduction
Bovine mastitis imposes a substantial economic burden on the global dairy industry, with direct losses attributed to reduced milk yield, treatment costs, and premature culling of infected animals. In southeastern Australia, the annual expenses of mastitis are estimated at AUD 150 million, driven by production losses, veterinary costs, and culling [1,2,3,4]. Globally, annual losses range between USD 19.7 and 32 billion, with subclinical mastitis contributing disproportionately due to undetected infections [5,6]. The disease also compromises animal welfare, causing udder pain, inflammation, and systemic illness, which reduces the quality of life for affected cows [4,6,7].
S. aureus is a leading causative agent of bovine mastitis worldwide, implicated in both clinical and subclinical forms. Its prevalence varies geographically: in southeastern Australia, S. aureus accounts for 10.6% of clinical and 29.1% of subclinical mastitis isolates [1]. In Ethiopia, a study of 15,000 cows across 46 herds reported a pooled prevalence of 35% for S. aureus mastitis, with subclinical cases predominant [8,9]. Similarly, in Central Oromia, Ethiopia, a 73.7% overall mastitis prevalence was observed, with S. aureus as the primary pathogen [10]. Regional disparities are evident; in China, S. aureus prevalence in clinical mastitis ranges from 10% to 66.6% across studies, influenced by herd management and diagnostic practices [11].
The pathogen’s ability to persist in mammary tissues and evade immune responses exacerbates its global burden. S. aureus biofilm formation and antimicrobial resistance (AMR) further complicate efforts to control the infection. In Ethiopia, beta-lactam resistance rates among S. aureus isolates reach 68%, linked to widespread antibiotic misuse [8]. Methicillin-resistant S. aureus (MRSA) strains, including livestock-associated variants, are increasingly reported, with mecA-mediated resistance posing zoonotic risks [12,13]. Despite low overall resistance rates in some regions, such as southeastern Australia, AMR surveillance remains crucial in mitigating therapeutic failures [1].
Diagnostic challenges compound the economic and health impacts of S. aureus mastitis. Intermittent bacterial shedding in milk complicates detection, with culture, PCR, and MALDI-TOF mass spectrometry exhibiting variable sensitivity [14,15]. Subclinical infections, often undiagnosed without tools like the California Mastitis Test, contribute to chronic herd infections and sustained transmission [7,10]. These factors underscore the need for enhanced diagnostics and tailored control strategies to address the pathogen’s epidemiological complexity.
This narrative review synthesizes structured data from scientific abstracts to examine the epidemiological aspects of S. aureus-associated bovine mastitis. The purpose is to consolidate current knowledge on global prevalence trends, geographic and herd-level distribution patterns, and clinical versus subclinical manifestations. It further explores challenges related to antimicrobial resistance, diagnostic limitations, and control strategies. The review does not interpret experimental or clinical trial data but focuses on synthesizing reported prevalence rates, risk factors, and resistance profiles. By analyzing these compiled findings, the review aims to highlight regional disparities, identify gaps in surveillance, and inform future research and public health interventions targeting S. aureus mastitis.
2. Literature Search Strategy
This narrative review employed a systematic literature search strategy to synthesize current knowledge on Staphylococcus aureus in bovine mastitis, with a focus on prevalence, antimicrobial resistance (AMR), pathogenesis, detection, and control strategies. The search was conducted across three major databases “PubMed, Scopus, and Web of Science Core Collection” spanning the last decade (January 2015 to June 2025).
The search strategy incorporated a comprehensive set of keywords and Boolean operators to capture relevant studies. Core terms included “Staphylococcus aureus” OR “S. aureus” combined with “bovine mastitis” OR “dairy cow mastitis” OR “udder infection”, and further refined using thematic subqueries: “prevalence” OR “epidemiology”, “antimicrobial resistance” OR “AMR” OR “MRSA”, “biofilm” OR “intracellular invasion” OR “virulence factors”, “detection” OR “diagnosis” OR “MALDI-TOF” OR “PCR” OR “AI”, and “control” OR “prevention” OR “vaccine” OR “One Health”. Example queries aligned with these groupings, such as (“Staphylococcus aureus”) AND (“bovine mastitis”) AND (“antimicrobial resistance”) AND (“biofilm formation”).
Studies were included if they met the following criteria: original research, systematic reviews, or meta-analyses reporting data on S. aureus prevalence, AMR profiles, virulence mechanisms, diagnostic methods, or control strategies in bovine mastitis; English-language publications or those with English abstracts; and geographic diversity covering global dairy systems. Exclusion criteria removed non-bovine studies (e.g., human or small ruminant focus), articles centered on non-S. aureus pathogens, non-peer-reviewed works (e.g., conference abstracts without full data), and publications lacking methodological rigor or statistical validation.
Screening occurred in two stages: an initial assessment of titles and abstracts for topical relevance, followed by a full-text evaluation of retained articles for methodological quality, data granularity, and alignment with the review objectives. To ensure comprehensiveness, reference lists of key publications were hand-searched for additional sources via snowballing. Extracted data encompassed prevalence rates, AMR gene profiles, diagnostic performance metrics, and intervention efficacy, and synthesized thematically to highlight regional disparities, temporal trends, and research gaps.
Study quality was evaluated based on sample representativeness, adherence to standardized diagnostic protocols (e.g., CLSI guidelines for AMR testing), and statistical robustness (e.g., confidence intervals for prevalence estimates). Limitations included potential language bias due to English-only inclusion, uneven geographic representation (e.g., scarce data from Africa and South America), and heterogeneity in diagnostic criteria (e.g., variable somatic cell count thresholds for subclinical mastitis).
3. Pathogenesis and Virulence Factors of S. aureus in Bovine Mastitis
3.1. Overview of Pathogenesis
S. aureus initiates bovine mastitis by invading the mammary gland through the teat canal (Figure 1), often exploiting mechanical damage or compromised teat keratin barriers [7,16]. Contagious transmission during milking facilitates colonization, driven by adhesins, such as fibronectin-binding proteins (FnBPs), clumping factors (ClfA/B), and collagen-binding proteins (Cna) [2,17]. These microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) enable attachment to host extracellular matrix components and mammary epithelial cells [18]. Post-colonization, the pathogen evades immune defenses via intracellular persistence within mammary epithelial cells or phagolysosomal compartments [6,19], alongside suppression of toll-like receptor (TLR)/NF-κB signaling to dampen inflammatory responses [20]. Chronicity is perpetuated by biofilm formation, toxin-mediated tissue damage, and nutrient acquisition systems, such as siderophores (staphyloferrin A/B) and the iron-responsive Isd system, which scavenge essential metals in the nutrient-restricted mammary environment [21,22].
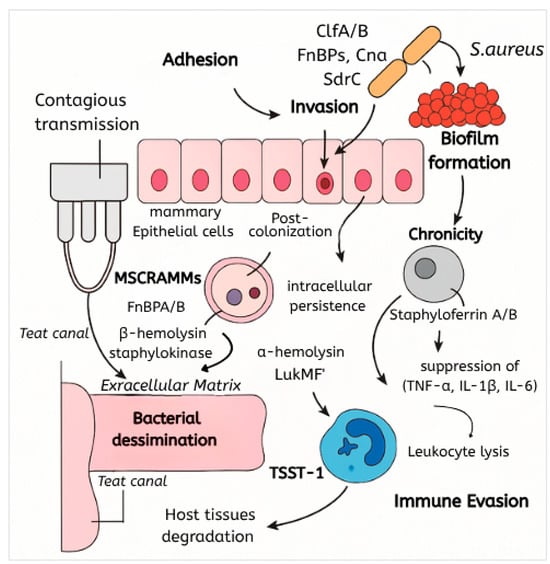
Figure 1.
Schematic of S. aureus pathogenesis and immune evasion.
3.2. Adhesion and Colonization Mechanisms
Adhesion is mediated by MSCRAMMs, including FnBPA/B, ClfA/B, Cna, and SdrC, which bind host fibronectin, collagen, and other extracellular matrix components [7,22]. Fibronectin-binding proteins (FnBPs) are crucial for initial attachment to the mammary epithelium, while clumping factors facilitate aggregation and biofilm initiation [18]. Collagen adhesin (Cna) enhances tissue specificity, particularly in damaged teat canals [23]. These interactions are foundational for establishing infection and subsequent biofilm development [24].
3.3. Toxin Production and Tissue Damage
S. aureus secretes cytolytic toxins and superantigens that exacerbate tissue damage and immune dysregulation. Pore-forming alpha-hemolysin disrupts epithelial integrity, while leukocidins, particularly LukMF’ (specific to bovine neutrophils), induce leukocyte lysis, impairing innate immunity [25]. Superantigens, including enterotoxins and toxic shock syndrome toxin-1 (TSST-1), trigger hyperinflammation through excessive cytokine release [20,26]. β-hemolysin and staphylokinase further degrade host tissues, facilitating bacterial dissemination [27,28].
3.4. Biofilm Formation and Persistence
Biofilm formation is a central virulence mechanism of S. aureus, particularly in chronic bovine mastitis [18,29,30]. These structured microbial communities are embedded in a self-produced extracellular matrix that adheres to host tissues or abiotic surfaces. In the context of the bovine mammary gland, biofilms facilitate bacterial persistence by enhancing resistance to host immune responses and antimicrobial therapies, thereby increasing the risk of chronic and recurrent intramammary infections (IMIs) [31].
The biofilm developmental process in S. aureus comprises four sequential stages: initial attachment, accumulation and maturation, maintenance, and dispersal. MSCRAMMs, such as FnBPs, ClfA/B, and Cna, primarily mediate the initial adherence to mammary epithelial surfaces. These proteins facilitate binding to host fibronectin and collagen, enabling bacterial colonization at the infection site [30,31].
Following the attachment, biofilm maturation involves the synthesis of extracellular polymeric substances, especially polysaccharide intercellular adhesin (PIA), which is encoded by the icaADBC operon [22]. PIA promotes intercellular aggregation and structural integrity of the biofilm. In some bovine strains, biofilm-associated protein (Bap) further enhances biofilm stability, especially under milk-rich environments. These mechanisms contribute to the robust biofilm phenotype observed in clinical mastitis isolates [24,32].
The accessory gene regulator (agr) quorum-sensing system governs biofilm dynamics. During early biofilm development, agr is typically repressed, which facilitates matrix accumulation and reduces immune detection [18]. In contrast, the later activation of agr promotes the production of dispersal agents, such as phenol-soluble modulins (PSMs) and proteases, allowing bacteria to exit the biofilm and colonize new sites [31]. This regulatory flexibility enables S. aureus to persist within the host while maintaining invasive potential. Environmental and host factors play critical roles in modulating biofilm formation. For example, milk components, such as caseins, enhance bacterial adhesion and matrix formation, serving as a nutrient source that supports high biomass production [33]. Notably, sub-inhibitory concentrations of antibiotics can paradoxically induce biofilm formation, complicating treatment strategies [31].
Biofilm formation significantly enhances the virulence of S. aureus through several mechanisms. First, the biofilm matrix impedes antibiotic penetration, rendering embedded bacteria up to 1000 times more resistant to treatment compared to planktonic cells [34]. Second, biofilms protect bacteria from phagocytosis and adaptive immune clearance, facilitating persistent colonization. Additionally, biofilms facilitate horizontal gene transfer, enabling the spread of AMR determinants and further complicating herd-level control [35].
Clinically, biofilm-associated mastitis is correlated with elevated somatic cell counts, reduced milk quality, and prolonged infection duration. Recent surveillance studies in dairy herds across Asia and Latin America confirm a high prevalence of strong biofilm-producing S. aureus strains, many of which also carry multidrug resistance genes [34,35].
3.5. Intracellular Invasion of S. aureus in Bovine Mastitis
S. aureus strains that cause bovine mastitis do more than colonize the mammary gland surface; many actively invade and persist inside bovine mammary epithelial cells (BMECs) and innate immune cells, creating protected reservoirs that drive chronic and recurrent infections [18]. Table 1 summarizes the key stages of S. aureus intracellular invasion in bovine mammary epithelial cells, highlighting recent mechanistic insights from molecular studies. It outlines bacterial entry mechanisms, subversion of host cellular processes (such as mitophagy), development of antibiotic-tolerant phenotypes, and the consequences for chronic infection and recurrence. These findings identify potential therapeutic targets for controlling intracellular S. aureus during mastitis.

Table 1.
Mechanisms of intracellular invasion and persistence of S. aureus in bovine mastitis.
3.6. Immune Evasion Strategies
Immune evasion involves multiple mechanisms: protein A binds to immunoglobulins, inhibiting opsonization [19], while capsular polysaccharides impede phagocytosis [6]. Intracellular survival within mammary epithelial cells and phagocytes avoids immune clearance [38,39], which is complemented by the suppression of proinflammatory cytokines (TNF-α, IL-1β, IL-6) and the induction of autophagy [20]. Leukocidin LukMF’ directly lyses neutrophils [40], and superantigens disrupt adaptive immunity by overactivating T cells [26]. Additionally, S. aureus downregulates host adhesion molecules (ICAM1, VCAM1) and counteracts nutritional immunity by acquiring metals via siderophores and the Isd system, overcoming lactoferrin-mediated iron sequestration [21,28].
4. Prevalence of S. aureus in Bovine Mastitis
4.1. Geographic Distribution
The global prevalence of S. aureus in bovine mastitis exhibits substantial regional variation, influenced by management practices and the use of antimicrobials. Table 2 synthesizes data from some studies worldwide on the prevalence of S. aureus mastitis in dairy herds, organized by continent and country.
Table 2.
Some reported prevalence of S. aureus mastitis in dairy herds.
In Africa, Cameroon reported a 67.0% prevalence (201/300 milk samples), while Egypt documented 44.4% (156/352 quarter milk samples), with MRSA detected in 23.0% and 95.0% of isolates, respectively [38,39]. Ethiopia identified S. aureus in 19% of 600 quarter milk samples, and Rwanda detected 135 isolates from 1080 quarter milk samples across 80 farms [41,42].
In Asia, Pakistan reported prevalence rates ranging from 30.32% (94/310 subclinical mastitis samples) to 42.5% (85/200 milk samples), with MRSA in 11.7–21.0% of isolates. Bangladesh documented 46.66% S. aureus prevalence (56/120 clinical mastitis samples), while Vietnam and Thailand recorded lower rates of 12.0% (48/400) and 4.76% (4/84), respectively [44,49,51]. South Korea detected MRSA in 6.1% (30/488 isolates), with resistance influenced by farm type [61]. Iran reported 24 S. aureus isolates from subclinical mastitis, and China identified a 7.0% prevalence (4/57 raw milk samples) in Shanghai [45,46].
European studies revealed a 32.95% herd-level prevalence in northern Italy (based on 88 bulk tank milk samples), 7.79% in Poland (36/462 milk samples), and 46.1% in Romania (150/325 CMT-positive samples), with MRSA detected in 25.0% of Romanian isolates [53,56]. Ireland reported annual prevalence rates of 21.37–25.59% across 7833 milk samples [52].
Mexico documented a 42% prevalence of S. aureus in 50 cows with subclinical mastitis in the Americas, while the U.S. reported 3.3% (158/4794 cows) in clinical cases. Canada observed no MRSA in 611 isolates but noted a 46.6% rate of penicillin resistance in clinical mastitis [57,59,60]. Brazil reported S. aureus in 66% of staphylococcal isolates from subclinical mastitis (507 quarter milk samples) [62].
4.2. Clinical vs. Subclinical Mastitis
S. aureus prevalence varies between clinical (CM) and subclinical mastitis (SCM). In Pakistan’s Narowal District, SCM exhibited higher prevalence (45.8%) than CM (37.5%), while Cameroonian MRSA isolates predominantly originated from clinical cases (72.5%) [38,49]. Conversely, a U.S. study linked S. aureus to 3.3% of clinical mastitis cases, correlating with elevated somatic cell counts [60]. Southern Italy reported a 59.16% prevalence of S. aureus in bulk tank milk from herds with clinical infections, compared to 29.16% for S. agalactiae [4].
Subclinical infections predominated in Ethiopia (15% of cases) and Brazil, where agr-negative strains with intracellular persistence mechanisms were associated with SCM [41,63]. A meta-analysis of 22 studies found no significant genetic differences in virulence gene prevalence between CM and SCM cases [64].
4.3. Herd-Level Risk Factors and Management Practices
Herd management systems have a significant influence on S. aureus epidemiology. Semi-intensive systems in Cameroon and conventional farming in Pakistan correlated with higher MRSA prevalence due to poor milking hygiene and antibiotic misuse [38,49]. South Korean organic farms exhibited lower antimicrobial resistance, despite higher isolation rates, which was attributed to the restricted use of antibiotics [61]. Larger Ethiopian farms with poor hygiene had elevated infection rates, while Irish pasture-based systems with seasonal calving showed persistent S. aureus infections (61.84% penicillin-resistant strains) [41,52]. Housing systems also played a role: deep litter systems in Italy correlated with higher prevalence than cubicles [4].
Clonal distribution analysis revealed limited diversity in Rwanda, dominated by CC97 (37%) and CC3666 (33%) lineages. In contrast, Brazilian isolates under CC97 displayed conserved spa types (t605, t521) [42,65].
5. Resistance Profiles and Commonly Reported Antimicrobials
5.1. Resistance Rates
S. aureus isolates from bovine mastitis exhibit extensive resistance to β-lactam antibiotics, with penicillin resistance rates ranging from 21.8% in Thailand/Cambodia to 100% in Egypt and Bangladesh [44,46,66,67]. Similarly, ampicillin resistance ranges from 45.74% to 85% in regions, such as Pakistan, Brazil, Iran, and Bangladesh [31,44,46,48]. Methicillin resistance is geographically variable, with MRSA prevalence ranging from 3.09% in China to 100% in Egypt. In contrast, Canadian and Thai/Cambodian studies report no MRSA [44,59,66,67,68].
Tetracycline resistance varies widely, from 7.3% in Thailand/Cambodia to 83.3% in Iran [46,66]. Resistance to erythromycin ranges from 2.6% in Canada to 57% in Bangladesh [44,59]. Multidrug resistance (MDR) is prevalent globally, with 50–83.3% of MRSA isolates resistant to ≥3 antibiotic classes in Pakistan, South Korea, Egypt, and Bangladesh [44,48,67,69]. Resistance to newer cephalosporins (e.g., cefquinome) and vancomycin remains low but is emerging, with vancomycin resistance genes (vanA, vanB) detected in 60.8–73.9% of Egyptian VRSA isolates [67,70].
5.2. Detection of Specific Resistance Genes
The mecA gene, which encodes methicillin resistance, is detected in 17.02–100% of S. aureus isolates, with a 100% prevalence in Egyptian MRSA and 66.67% prevalence in broader studies [41,48,67,69]. Beta-lactamase gene blaZ is prevalent in 23–73% of isolates, with higher frequencies in human-associated strains compared to bovine isolates [48,52,70,71]. Tetracycline resistance genes (tetK, tet(38)) are reported in 46.80% of Pakistani and Irish isolates, respectively (Table 3) [48,52].
Aminoglycoside resistance genes (ant(4′)-Ia, aac(6′)-Ie + aph(2″)) are prominent in South Korean MRSA (70% and 40%, respectively). In contrast, macrolide resistance genes (ermC, msrA) and efflux pump genes (norA, lmrS) are detected at lower frequencies [69,72,73]. Unique findings include the fosB fosfomycin resistance gene in U.S. CC133 isolates and phenotypic oxacillin resistance without mecA in Brazilian CC97 strains [65,73].
Table 3.
Regional prevalence of AMR genes in S. aureus mastitis isolates (reported as a percentage of isolates positive for the gene).
Key Points
- -
- mecA prevalence peaks in Egypt (100%) and Pakistan (17–44.8%), while Thailand, USA, Canada, and Brazil report 0%.
- -
- blaZ is highest in Brazil (100%), followed by Pakistan (55.3%) and New Zealand (23%).
- -
- tetK is only reported in Pakistan (46.8%); no other regions provide data.
- -
- Critical gaps exist for Africa (outside Egypt) and South America (outside Brazil). Phenotypic resistance often lacks genetic validation.
- -
- NR: not reported
5.3. Prevalence and Characteristics of MRSA
MRSA strains demonstrate high resistance to β-lactams (90–100%), kanamycin (66.7%), and non-β-lactams, with MDR rates exceeding those of methicillin-susceptible S. aureus (MSSA) [48,69]. SCCmec typing reveals geographic dominance: SCCmec IV in South Korea (ST72-t324 lineage), SCCmec I in Egypt, and SCCmec IV/V in Pakistan [47,67,69]. MRSA isolates frequently harbor biofilm-related genes (clfA, icaA/D) and virulence factors (pvl, tsst-1, hlb, lukMF), enhancing persistence and zoonotic potential [44,67,69]. Longitudinal data show rising MRSA prevalence in Germany (2% to 5% over a decade) and phenotypic-genotypic discordance in oxacillin-resistant mecA-negative Brazilian isolates [65,70]. Figure 2 shows the prevalence of MRSA in milk and S. aureus isolates across selected countries.
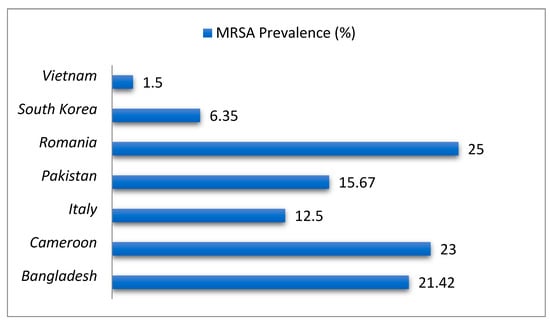
Figure 2.
Prevalence of MRSA in milk and S. aureus isolates across selected countries.
5.4. Comparative and Cumulative Resistance Patterns
Resistance to β-lactams and tetracyclines is globally widespread, driven by therapeutic overuse, whereas resistance to aminoglycosides and sulfonamides remains comparatively low [46,59]. Regional disparities in resistance gene prevalence (e.g., blaZ in 73% of human-associated vs. 2.6% of bovine isolates in Vermont) reflect divergent selection pressures [74]. Declining erythromycin resistance in Germany (53% to 8%) correlates with antimicrobial stewardship, underscoring the impact of targeted interventions [70]. Genotypic–phenotypic discordance, observed in VRSA lacking vanA/vanB and oxacillin-resistant mecA-negative isolates, highlights unresolved resistance mechanisms [65,66].
Cumulatively, recurrent intramammary infections, prophylactic antibiotic use, and horizontal gene transfer contribute to the development of MDR, with urgent needs for alternative therapies and enhanced surveillance to curb the dissemination of resistance [41,75,76].
6. Detection and Identification Methods for S. aureus in Bovine Mastitis
6.1. Conventional Culture and Biochemical Identification
Bacteriological culture remains the cornerstone for detecting S. aureus in bovine mastitis. Milk samples are cultured on selective and differential media, including blood agar, mannitol salt agar (MSA), Baird–Parker agar (with egg yolk tellurite), MacConkey agar, Columbia CNA agar, and Chromagar Mastitis, under aerobic incubation at 35–37 °C for 18–48 h [52,57,77,78]. Enrichment steps, such as pre-incubation in tryptic soy broth or nutrient broth with high salt, improve recovery rates [54]. Presumptive identification relies on colony morphology (e.g., golden-yellow pigmentation, hemolysis) and biochemical assays, including Gram staining for Gram-positive cocci, catalase positivity, coagulase activity (via tube or rapid tests), DNase production, and mannitol fermentation [57,61,77]. Chromogenic agars (e.g., Chromagar Mastitis) aid differentiation, while API test systems (e.g., ID 32 STAPH) or automated platforms, like VITEK 2 Compact, provide biochemical profiling for species confirmation [52,61,77]. The California Mastitis Test (CMT) serves as an initial on-farm screening tool for subclinical cases by detecting elevated somatic cell counts [78,79].
6.2. Molecular Detection Methods
Molecular techniques enhance specificity and speed. Conventional PCR targeting the nuc gene is the gold standard for species confirmation, achieving 100% concordance with phenotypic methods in some studies [80]. Real-time PCR assays, including probe-based systems, detect nuc with 100% sensitivity and specificity within 2 h [81]. Multiplex PCR kits (e.g., Mastit 4, bactotype HP3) concurrently identify S. aureus and co-pathogens (e.g., streptococci, Klebsiella pneumoniae), with sensitivities of 63.3–85.6% and specificities of 93.6–99.2% [45,82]. Resistance genes (mecA for methicillin resistance, vanA, blaZ) and virulence factors (clfA, fnbA, pvl, icaD, Hla) are detected via PCR or SCCmec typing [61,74,83]. Recombinase-aided amplification (RAA) coupled with lateral flow dipsticks (LFD) enables rapid (<45–85 min) field-deployable detection of nuc at sensitivities as low as 60 fg DNA [45]. Whole-genome sequencing (WGS) and spa gene typing provide strain-level resolution (e.g., ST1, ST97) and comprehensive virulence/resistance profiling [73,84]. Fluorescence in situ hybridization (FISH) with the Sau 16S69 probe visualizes S. aureus aggregates in tissues [85].
6.3. Automated and Commercial Diagnostic Systems
Automated systems streamline workflows. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) achieves rapid (<24 h), accurate species identification (confidence scores > 2.0). However, differentiation from closely related species (e.g., S. rostri) requires updated databases [2,86]. The Bacticam AI-based image classifier automates the analysis of bacterial growth on SELMA + multi-agar plates, achieving a sensitivity of over 85% and a specificity of over 91.9% compared to MALDI-TOF [87]. Commercial qPCR kits (e.g., Mastitis 4E) demonstrate high sensitivity (≥0.95) and specificity (≥0.99) for bulk milk testing [4]. Lateral flow immunoassays (LFIA) targeting EF-Tu protein offer rapid results (15 min) but require high bacterial loads (≥106 CFU/mL) [88]. Gold nanoparticle-enhanced ELISA improves sensitivity (97%) for antigen detection in milk, outperforming conventional ELISA [40]. The CombiFoss 7 DC integrates somatic cell analysis but relies on complementary culture for pathogen-specific identification [89].
6.4. Comparative Performance and Validation
As described in Table 4, culture methods exhibit variable sensitivity (50–95.5%) and specificity (79.0–91.4%), with improvements after pre-incubation [54,82]. PCR enhances detection rates by 9–18% over culture alone but faces cost and infrastructure barriers in resource-limited settings [41,90]. Real-time PCR demonstrates superior specificity (97.0–97.6%) compared to culture (89.1%) but lower sensitivity (63.3–78.1%) in pooled samples [82]. MALDI-TOF MS reduces turnaround time but cannot detect resistance genes [86]. The duplex RAA-LFD assay exhibits high concordance (κ > 0.92) with reference methods and detects S. aureus within 6 h post-enrichment [45]. Commercial qPCR kits surpass culture in specificity for contagious pathogens, while nanoparticle-enhanced ELISA reduces reliance on culture [4,40].
Table 4.
Comparative Analysis of Diagnostic Methods for S. aureus Mastitis.
7. Application of Artificial Intelligence in the Diagnosis and Management of S. aureus-Associated Bovine Mastitis
7.1. AI-Assisted Detection of S. aureus
Artificial intelligence enhances diagnostic precision for S. aureus-associated mastitis through multimodal approaches (Figure 3). Electrochemical impedance immunosensors functionalized with anti-S. aureus antibodies, combined with decision tree classifiers, achieved 95% accuracy in detecting bacterial concentrations in milk samples, enabling on-farm identification of subclinical infections [91]. Infrared thermography (IRT) paired with convolutional neural networks (CNNs) localized udder inflammation via temperature differentials, though environmental variability reduced sensitivity to 67% in field conditions [92,93]. Deep learning models trained on 16,000 teat-end images automated hyperkeratosis scoring, aiding in the detection of subclinical mastitis [94]. Automated image analysis of chromogenic agar plates improved S. aureus identification in milk samples after 24-h incubation, though sensitivity gaps persisted for low-prevalence pathogens [95].
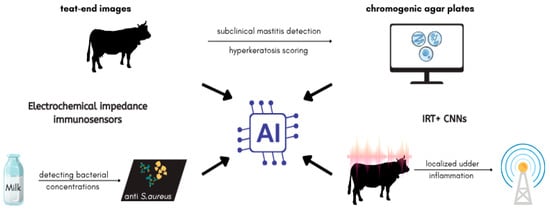
Figure 3.
AI-assisted mastitis monitoring system overview.
7.2. Predictive Modeling for Mastitis Risk and Herd Monitoring
Machine learning (ML) models integrate genomic, sensor, and management data to forecast S. aureus mastitis risks. Explainable AI (XAI) methods, including SHapley Additive exPlanations (SHAP), identified immune-related genomic markers (e.g., 204,642 SNPs) linked to mastitis susceptibility in Holstein cows, achieving 65% accuracy in classifying genetic risk [96]. Random Forest (RF) and Extreme Gradient Boosting (XGBoost) algorithms analyzed time-series sensor data (e.g., milk conductivity, yield) to predict the onset of clinical mastitis with 85% recall, although specificity varied (31–62%) [97]. Herd-level models incorporating somatic cell counts (SCC), milk yield, and climatic variables predicted mastitis outbreaks in buffalo herds with 76% accuracy, albeit with suboptimal sensitivity (67%) [98]. Network models simulating cattle movement and farm contact data further supported herd-level assessments of S. aureus transmission risk [99].
7.3. AI-Enhanced Treatment and AMR Management
AI-driven platforms address S. aureus AMR and optimize therapy. Machine learning classifiers applied to MALDI-TOF mass spectrometry data achieved 96.8% accuracy in predicting MDR S. aureus strains and 97.5% accuracy for benzylpenicillin resistance, using ribosomal and DNA-binding protein biomarkers identified in 82 isolates [100]. AI platforms that correlate protein–protein interaction networks with resistance mechanisms enable phenotype-guided antimicrobial selection, reducing the empirical use of broad-spectrum antibiotics [101]. Digital twins that integrate real-time sensor data (e.g., udder temperature, feeding behavior) dynamically update treatment recommendations, although pathogen-specific workflows require further development [102]. ML models also guided selective dry cow therapy (SDCT) by identifying cows that required antimicrobial intervention, although “black-box” limitations hindered farmer adoption [103].
7.4. Challenges and Future Directions
The integration of AI in managing S. aureus-associated bovine mastitis faces significant hurdles, primarily stemming from dataset limitations. Current models often rely on small, regionally constrained training datasets, which undermines their robustness and generalizability. For instance, Beck et al. (2024) trained anomaly-detection algorithms on only (n = 58) metagenomic milk samples, which risks overfitting and fails to capture global pathogen diversity [101]. Similarly, genomic susceptibility studies using whole-genome sequences from 52 cows struggled with biological variability, highlighting the “p >> n” problem (predictors far exceeding the number of samples) in complex trait prediction [96].
Adoption barriers further obstruct practical implementation. High costs for sensors, sequencing, and computational infrastructure limit accessibility, particularly for small-scale farms [104]. Compounding this, low farmer tech-literacy complicates interaction with AI interfaces, as seen in studies where complex data management requirements reduced user engagement [105]. Crucially, the “black-box” nature of AI models erodes stakeholder trust; Rowe et al. (2025) documented veterinarian skepticism toward ML-driven treatment recommendations due to the opacity of decision pathways [103].
Addressing these challenges necessitates rigorous validation frameworks. Multi-herd trials are crucial for evaluating generalizability across different breeds, climates, and management systems. For example, teat-end hyperkeratosis classifiers trained on images from two farms require validation in diverse geographical settings to confirm utility [94]. Similarly, MALDI-TOF resistance predictors (with 96.8% accuracy) were used on isolates from single countries, necessitating cross-border studies to verify their global applicability [100]. Longitudinal real-world testing is equally critical; early-detection algorithms (e.g., Random Forest models with 85% recall) must be assessed over extended periods to ensure durability against pathogen evolution [97].
Future advancements should prioritize federated learning to pool data across institutions without compromising privacy, enhancing dataset representativeness [106]. XAI techniques, such as SHAP value visualizations, can demystify model logic and rebuild stakeholder trust [96]. Finally, establishing open-access, multi-herd consortia will standardize validation protocols and accelerate the translation of AI innovations into herd-specific solutions [107].
8. Public Health Implications of S. aureus from Bovine Mastitis
8.1. Zoonotic Transmission and Occupational Exposure
S. aureus from bovine mastitis poses significant zoonotic risks through direct occupational exposure to infected cattle, contaminated milk, or dairy environments. Canadian studies demonstrated that bovine-derived strains invade and kill human intestinal cells (Caco-2) and cause lethal infections in Caenorhabditis elegans, highlighting cross-species pathogenicity despite lacking human adaptation genes [108]. Molecular epidemiological evidence from Tanzania, Colombia, and Pakistan confirmed bidirectional transmission, with genotypic similarities in erythromycin resistance genes (ermA, ermB, ermC) between bovine mastitis isolates and farm workers’ nasal swabs [109,110,111]. Environmental reservoirs, such as hock skin in Dutch herds and milking equipment in Brazilian farms, further amplify occupational risks, particularly with multidrug-resistant strains [112,113]. Poor hygiene practices, inadequate protective measures, and prolonged bacterial shedding in subclinical infections exacerbate colonization risks for dairy workers [111,114].
8.2. Foodborne Transmission and Enterotoxin-Related Risks
Raw milk contaminated with enterotoxigenic S. aureus strains, including MRSA variants, represents a critical food safety concern. Enterotoxin genes (sea, seb, sed, seg, tst) were identified in isolates from India, Tanzania, and Kazakhstan, with biofilm-forming strains enhancing toxin persistence in dairy products [109,115,116]. Subclinical mastitis in cows contributes to undetected pathogen shedding, particularly in regions with high consumption of raw milk. MRSA genotype B (GTB) in raw milk cheese highlights the risks associated with unprocessed dairy products [45]. While pasteurization mitigates risks, interventions, like geraniol (a natural antimicrobial) and bacteriophage therapy, show promise in reducing contamination and antibiotic residues [84,117].
8.3. MRSA Strains and Human Health Threats
MRSA from bovine mastitis carries mecA and exhibits resistance to β-lactams, tetracyclines, and lincomycin. Canadian MRSA isolates invaded human cells and resisted intracellular antibiotic clearance, raising concerns about treatment failures [108]. Strains from Argentina (ST83, spa t002) and China (S. equorum, S. saprophyticus) highlight zoonotic risks, with virulence genes (hlg, pvl, tsst-1) exacerbating pathogenicity [118,119]. Indian MRSA isolates demonstrated 100% resistance to cefoxitin and 91.3% to oxacillin, alongside biofilm-forming capabilities [115]. Synergistic therapies, such as red ginseng extract, and eradication programs in Switzerland emphasize the need for biosecurity and antibiotic stewardship to curb AMR dissemination [84,120].
8.4. One Health Interventions and Surveillance
Integrated One Health strategies are crucial for addressing zoonotic transmission (Figure 4), AMR spread, and food safety risks. Recommendations include enhanced antimicrobial stewardship, rapid diagnostics (e.g., nanopore metagenomics), and alternatives, like phage endolysins or graphene oxide, to reduce antibiotic reliance [121,122]. Surveillance of extramammary reservoirs (e.g., hock skin, milking equipment) and improved milking hygiene are essential to interrupt transmission cycles [112,123]. Successful eradication programs in Switzerland and Colombia emphasize biosecurity, segregated milking, and farmer education [111,120].
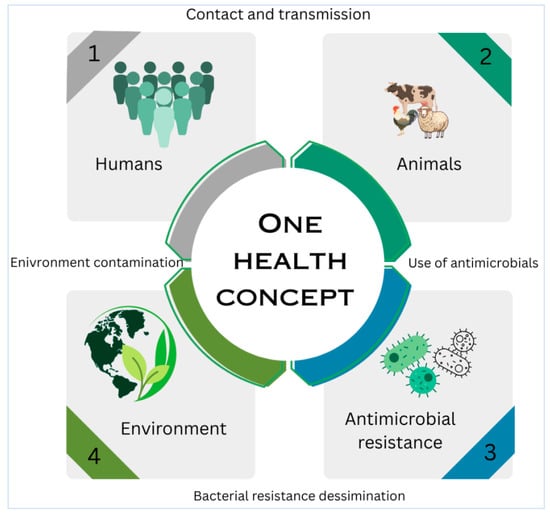
Figure 4.
One Health conceptual framework showing zoonotic transmission routes.
9. Prevention and Prophylaxis of S. aureus in Bovine Mastitis
Staphylococcus aureus mastitis remains a persistent global challenge in dairy production, characterized by significant economic losses, chronic infections, and the evolution of antimicrobial resistance. Practical control demands integrated, multi-faceted strategies due to the pathogen’s capacity for immune evasion, biofilm formation, intracellular persistence, and antigenic diversity. Table 5 synthesizes evidence-based global control approaches.
Table 5.
Global control strategies for S. aureus mastitis.
9.1. Hygienic and Management Practices
Pre- and post-milking teat disinfection is a cornerstone of S. aureus control. Polyhexamethylene biguanide (PHMB)-based teat disinfectants form a protective antimicrobial barrier, achieving >4-log reduction of S. aureus and residual efficacy for 12 h, outperforming traditional iodine-based products [124]. Silver (AgNPs) and copper nanoparticles (CuNPs) at 0.78 mg/L reduce S. aureus by 95–97%, disrupting biofilms and inducing bacterial oxidative stress [125]. Bacteriocin-producing lactic acid bacteria (LAB), such as Lactococcus cremoris FT27, demonstrate efficacy comparable to that of chemical disinfectants through nisin A production [126]. Post-milking teat dipping reduces new intramammary infections (IMIs) by 62% in field trials [127]. Environmental hygiene measures, including daily lime application, clean straw bedding, and cubicle maintenance, lower bacterial exposure. Automated milking systems (AMS) require tailored protocols, supported by farmer education programs [128]. Improved teat and udder conformation in indigenous breeds correlates with a reduced prevalence of subclinical mastitis (SCM), highlighting the genetic selection benefits. Strict aseptic techniques during dry-off, combined with PCR-based milk monitoring, enable timely interventions [4].
9.2. Vaccination and Immunoprophylaxis
Vaccination against S. aureus mastitis remains challenging due to antigenic variability, biofilm formation, and transient immune responses. Experimental vaccines targeting Staphylococcus chromogenes surface proteins (SCSP) reduced subclinical mastitis incidence by 90% at cow and quarter levels, whereas S. aureus surface protein (SASP) vaccines showed non-significant protection (48%) [129]. A trivalent inactivated vaccine (S. aureus, E. coli, M. bovis) reduced clinical mastitis signs and milk bacterial counts to undetectable levels within 96 h post-challenge in heifers [130]. Commercial bacterins and autogenous vaccines yield modest reductions in clinical severity but fail to alter the incidence of infection or shedding [131]. Novel approaches, including live-attenuated mutants (e.g., vraG-deleted strains) and nanogel-based formulations, aim to enhance Th1/Th17 immunity. Despite progress, no universal vaccine matches the efficacy of hygiene practices [132].
9.3. Antimicrobial Prophylaxis and Dry Cow Therapy
Dry cow therapy (DCT) with antibiotics remains critical for controlling subclinical S. aureus infections, although cure rates are modest (30–50%) due to intracellular persistence [128]. Selective DCT, guided by somatic cell count (SCC) thresholds (<200,000 cells/mL), reduces antibiotic use by 20–60% without compromising udder health. However, 11.8% of cows receiving internal teat sealants (ITS) alone harbored persistent S. aureus infections, particularly high-yielding cows (>15 kg/day), which exhibit elevated somatic cell counts (SCC) in early lactation. Combining ITS with antibiotics during the dry period reduces early lactation SCC and IMIs compared to ITS alone, which increases IMI odds by 6- to 7-fold [133,134]. Lactobacillus plantarum CM49 also shows broad antagonistic activity against S. aureus [135].
9.4. Selective Treatment Protocols
Tailored protocols integrating SCC trends, bacterial cultures, and algorithm-based decision support improve diagnostic accuracy for selective DCT. Segregation of infected cows delays S. aureus transmission and lowers treatment costs. Herds employing PCR pooling for routine screening report reduced antimicrobial use and improved udder health. Culling chronically infected animals and optimizing dry period management further reduce early lactation SCC. In Irish herds with high S. aureus prevalence (19.7% herd-level IMI), selective DCT efficacy depended on herd-specific management factors [4,133,136].
9.5. Resistance Concerns Linked to Preventive Antimicrobial Use
High beta-lactam resistance rates in S. aureus isolates, particularly to penicillin (75%), amoxicillin (67%), and cephalosporins (57%), are driven by beta-lactamase production (via the blaZ gene) and biofilm-mediated tolerance [8]. In Ethiopia, S. aureus isolates showed a pooled beta-lactam resistance prevalence of 35% (95% CI: 31–41%), whereas non-aureus staphylococci (NAS), such as S. chromogenes, harbored blaZ at a 22% prevalence. Molecular typing and genomic surveillance are crucial for identifying resistant genotypes and informing stewardship practices [137]. Overuse of antibiotics in non-targeted DCTs exacerbates resistance, emphasizing the need for farmer education and the development of selective protocols [11].
9.6. Herd-Level Outcomes and Program Efficacy
Integrated programs that combine hygiene, early diagnosis, and culling reduce the prevalence of S. aureus and associated economic losses. In Bavaria, a 10-year study documented a decline from 26% (in 2014) to 15% (in 2023) in culture-positive samples [127]. Herds implementing post-milking disinfection and cubicle cleaning twice daily achieved lower SCC. Modeling supports a “bang-bang” strategy, where initial low cow mixing is followed by phased productivity increases to balance biosecurity and output [136]. Welfare scoring systems correlate lower S. aureus prevalence with higher welfare outcomes. Intensive systems reported a higher prevalence of mastitis (39.5%), underscoring the need for robust hygiene [9,133].
10. Future Directions
Geographic and molecular surveillance gaps hinder a comprehensive understanding of S. aureus mastitis dynamics. Regional disparities in capsule genotype distribution (e.g., CP8) and AMR patterns, such as elevated tetracycline resistance in Jordan and ciprofloxacin resistance in Bangladesh, necessitate expanded global surveillance to address zoonotic risks and region-specific AMR trends [138,139]. Concurrently, molecular characterization of virulence factors, including ferroptosis-related genes (HMOX1, SAT1), immune pathways (TLR2, NF-kappa B), and strain-specific determinants (adlb, agr systems, MSCRAMMs), remains incomplete. Advanced genotyping (e.g., spa t2873, CC8/CC97 strains) and WGS are critical to elucidate transmission dynamics, biofilm-associated genes, and conserved antigen targets for vaccine development [63,84,140,141,142].
Subclinical mastitis caused by multidrug-resistant S. aureus in riverine buffaloes is understudied despite the high prevalence of virulence genes. Prioritizing this form through improved diagnostic accuracy and tailored interventions is essential to mitigate undetected transmission and economic losses [139].
Standardization of methodologies and diagnostic optimization is imperative for field applicability. Refining duplex RAA-LFD and AuNPs-based ELISA for sensitivity, alongside standardizing phage therapy production protocols, will enhance reproducibility [40,45,143]. Parallel advancements in AI-driven classifiers (e.g., Bacticam), lateral flow immunoassays (LFIA) with enhanced antibody specificity, and portable biosensors for real-time detection necessitate the expansion of training datasets and harmonized sample handling protocols [54,88].
Therapeutic innovation must address the resilience of biofilms and intracellular persistence. Promising strategies include phage therapy formulations, anti-biofilm agents (e.g., gemini quaternary ammonium salts), NLRP3 inflammasome inhibitors, and probiotics derived from non-aureus staphylococci (NAS). Vaccine development should prioritize adhesins, hemolysins, and biofilm components to disrupt host-pathogen interactions. [51,85,144,145].
AMR management requires region-specific AMR profiling and stewardship programs guided by molecular resistance gene tracking. Policies that enforce prudent antibiotic use, farmer education on biosecurity, and alternatives, such as phage biocontrol, are vital to curb the escalation of resistance [47,138,139,146,147].
Longitudinal and herd-level studies are crucial for validating the efficacy of mastitis control programs, particularly their environmental impacts (e.g., greenhouse gas emissions) and long-term effects on milk yield. Integrating genetic markers into breeding programs may enhance herd resilience against pathogen-specific mastitis [4,148,149].
11. Conclusions
S. aureus remains a pervasive and economically burdensome pathogen in bovine mastitis worldwide, characterized by its virulence arsenal, diagnostic challenges, and AMR. This narrative synthesis underscores substantial geographic variability in prevalence, resistance profiles, and diagnostic efficacy, reflecting disparities in herd management practices, biosecurity measures, and surveillance infrastructure. The pathogen’s ability to persist intracellularly, form biofilms, and evade immune responses contributes to chronic and subclinical infections that complicate early detection and control.
The widespread detection of MDR and MRSA strains in dairy herds across continents underscores the urgent need for targeted AMR stewardship and genomic surveillance. Diagnostic advances, including molecular assays, AI-enhanced platforms, and rapid biosensors, offer promising avenues for early and precise detection but require standardization and resource adaptation for widespread adoption.
From a One Health perspective, the zoonotic potential of S. aureus, particularly MRSA, through occupational exposure and foodborne transmission, represents a significant public health concern. Integrating antimicrobial stewardship, farmer education, hygienic milking practices, and selective dry cow therapy remains critical to mitigate infection risks and curb the dissemination of resistance.
Future interventions must prioritize vaccine development targeting conserved virulence determinants, the deployment of AI-driven predictive tools for mastitis management, and longitudinal studies assessing environmental, genetic, and herd-level dynamics. A coordinated global response, anchored in robust surveillance, translational research, and policy harmonization, is essential to contain the spread of S. aureus-associated bovine mastitis and safeguard both animal and public health.
Funding
This work was supported and funded by the Deanship of Scientific Research at Imam Mohammad Ibn Saud Islamic University (IMSIU) (grant number IMSIU-DDRSP2502).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article. Raw data supporting this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMR | Antimicrobial Resistance |
| AI | Artificial Intelligence |
| BTM | Bulk Tank Milk |
| CFU/mL | Colony Forming Units per Milliliter |
| CMT | California Mastitis Test |
| CM | Clinical Mastitis |
| CNN | Convolutional Neural Network |
| DCT | Dry Cow Therapy |
| EF-Tu | Elongation Factor Thermo-unstable |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| IRT | Infrared Thermography |
| ITS | Internal Teat Sealant |
| LFD | Lateral Flow Dipstick |
| LFIA | Lateral Flow Immunoassay |
| MALDI-TOF MS | Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry |
| MDR | Multidrug Resistance |
| MRSA | Methicillin-Resistant Staphylococcus aureus |
| MSCRAMMs | Microbial Surface Components Recognizing Adhesive Matrix Molecules |
| MSSA | Methicillin-Susceptible Staphylococcus aureus |
| NAS | Non-aureus Staphylococci |
| PCR | Polymerase Chain Reaction |
| PIA | Polysaccharide Intercellular Adhesin |
| qPCR | Quantitative Polymerase Chain Reaction |
| RAA | Recombinase-Aided Amplification |
| RF | Random Forest (Machine Learning Algorithm) |
| SCC | Somatic Cell Count |
| SCCmec | Staphylococcal Cassette Chromosome mec |
| SDCT | Selective Dry Cow Therapy |
| SHAP | SHapley Additive exPlanations (Explainable AI Technique) |
| SNP | Single Nucleotide Polymorphism |
| ST | Sequence Type (used in molecular typing) |
| TSST-1 | Toxic Shock Syndrome Toxin-1 |
| WGS | Whole-Genome Sequencing |
| XAI | Explainable Artificial Intelligence |
| XGBoost | Extreme Gradient Boosting (Machine Learning Algorithm) |
References
- Dyson, R.; Charman, N.; Hodge, A.; Rowe, S.M.; Taylor, L.F. A survey of mastitis pathogens including antimicrobial susceptibility in southeastern Australian dairy herds. J. Dairy Sci. 2022, 105, 1504–1518. [Google Scholar] [CrossRef] [PubMed]
- Langhorne, C.; Horsman, S.; Wood, C.; Clark, R.; Price, R.; Henning, J.; Grewar, J.D.; Wood, B.J.; Ranjbar, S.; McGowan, M.R.; et al. Bacterial culture and susceptibility test results for clinical mastitis samples from Australia’s subtropical dairy region. J. Dairy Sci. 2024, 107, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, Z.; Cabrera, V.E.; Hogeveen, H.; Ruegg, P.L. Economic impact of subclinical mastitis treatment in early lactation using intramammary nisin. J. Dairy Sci. 2024, 107, 4634–4645. [Google Scholar] [CrossRef]
- Zaghen, F.; Sora, V.M.; Zanirato, G.; Zecconi, A. From One Heath to One Sustainability: The Role of Contagious Mastitis Pathogens in Decreasing the Dairy Herd Sustainability. Pathogens 2024, 13, 914. [Google Scholar] [CrossRef]
- Morales-Ubaldo, A.L.; Rivero-Perez, N.; Valladares-Carranza, B.; Velázquez-Ordoñez, V.; Delgadillo-Ruiz, L.; Zaragoza-Bastida, A. Bovine mastitis, a worldwide impact disease: Prevalence, antimicrobial resistance, and viable alternative approaches. Vet. Anim. Sci. 2023, 21, 100306. [Google Scholar] [CrossRef]
- Saeed, S.I.; Kamaruzzaman, N.F.; Gahamanyi, N.; Nguyen, T.T.H.; Hossain, D.; Kahwa, I. Confronting the complexities of antimicrobial management for Staphylococcus aureus causing bovine mastitis: An innovative paradigm. Ir. Vet. J. 2024, 77, 4. [Google Scholar] [CrossRef]
- Exel, C.E.; Halasa, T.; Koop, G.; Steeneveld, W.; Lam, T.J.G.M.; Benedictus, L.; Gussmann, M. A stochastic modelling approach to determine the effect of diverse Staphylococcus aureus strains on the economic and epidemiological outcomes of mastitis intervention strategies in dairy cattle. Prev. Vet. Med. 2022, 199, 105566. [Google Scholar] [CrossRef]
- Fenta, M.D.; Tafere, F.A.; Mebratu, A.S.; Malede, B.A. Quarter-wise proportion and beta-lactam resistance rate of bovine mastitis associated- Staphylococcus aureus among infectious episodes in Ethiopia: Systematic review and meta-Analysis. Heliyon 2023, 9, e18180. [Google Scholar] [CrossRef] [PubMed]
- Tora, E.T.; Bekele, N.B.; Suresh Kumar, R.S. Bacterial profile of bovine mastitis in Ethiopia: A systematic review and meta-analysis. PeerJ 2022, 10, e13253. [Google Scholar] [CrossRef]
- Fesseha, H.; Mathewos, M.; Aliye, S.; Wolde, A. Study on Prevalence of Bovine Mastitis and Associated Risk Factors in Dairy Farms of Modjo Town and Suburbs, Central Oromia, Ethiopia. Vet. Med. Res. Rep. 2021, 12, 271–283. [Google Scholar] [CrossRef]
- Wang, K.; Cha, J.; Liu, K.; Deng, J.; Yang, B.; Xu, H.; Wang, J.; Zhang, L.; Gu, X.; Huang, C.; et al. The prevalence of bovine mastitis-associated Staphylococcus aureus in China and its antimicrobial resistance rate: A meta-analysis. Front. Vet. Sci. 2022, 9, 1006676. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, I.N.; Latorre-Fernández, J.; Reuben, R.C.; Trabelsi, I.; González-Azcona, C.; Arfaoui, A.; Usman, Y.; Lozano, C.; Zarazaga, M.; Torres, C. Beyond the Wild MRSA: Genetic Features and Phylogenomic Review of mecC-Mediated Methicillin Resistance in Non-aureus Staphylococci and Mammaliicocci. Microorganisms 2023, 12, 66. [Google Scholar] [CrossRef]
- Dendani Chadi, Z.; Dib, L.; Zeroual, F.; Benakhla, A. Usefulness of molecular typing methods for epidemiological and evolutionary studies of Staphylococcus aureus isolated from bovine intramammary infections. Saudi J. Biol. Sci. 2022, 29, 103338. [Google Scholar] [CrossRef]
- Kurban, D.; Roy, J.-P.; Kabera, F.; Fréchette, A.; Um, M.M.; Albaaj, A.; Rowe, S.; Godden, S.; Adkins, P.R.F.; Middleton, J.R.; et al. Diagnosing Intramammary Infection: Meta-Analysis and Mapping Review on Frequency and Udder Health Relevance of Microorganism Species Isolated from Bovine Milk Samples. Animals 2022, 12, 3288. [Google Scholar] [CrossRef]
- Mues, L.; Kemper, N.; Blumenberg, J.A. Occurrence and diagnostic of intermittent shedding of Staphylococcus aureus in bovine mammary infection. Front. Vet. Sci. 2025, 12, 1523698. [Google Scholar] [CrossRef]
- Rowe, S.; House, J.K.; Pooley, H.; Bullen, S.; Humphris, M.; Ingenhoff, L.; Norris, J.M.; Zadoks, R.N. Evaluation of point-of-care tests for identification of pathogens to inform clinical mastitis treatment decisions in pasture- and confinement-managed dairy cows in Australia. J. Dairy Sci. 2024, 107, 8271–8285. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Barkema, H.W.; Nobrega, D.B.; Xu, C.; Han, B.; Zhang, C.; Yang, J.; Li, X.; Gao, J. Virulence of Bacteria Causing Mastitis in Dairy Cows: A Literature Review. Microorganisms 2025, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Pickering, A.C.; Rocha, L.S.; Aguilar, A.P.; Fabres-Klein, M.H.; de Oliveira Mendes, T.A.; Fitzgerald, J.R.; de Oliveira Barros Ribon, A. Diversity and pathogenesis of Staphylococcus aureus from bovine mastitis: Current understanding and future perspectives. BMC Vet. Res. 2022, 18, 115. [Google Scholar] [CrossRef]
- Kerro Dego, O.; Vidlund, J. Staphylococcal mastitis in dairy cows. Front. Vet. Sci. 2024, 11, 1356259. [Google Scholar] [CrossRef]
- Zhang, L.; Ye, X.; Zhang, Y.; Wang, F.; Zhang, F.; Jia, Y.; Wu, D.; Tohti, K.; Cheng, M.; Zhu, J. Anti-Staphylococcus aureus Single-Chain Fragment Variables Play a Protective Anti-Inflammatory Role In Vitro and In Vivo. Vaccines 2021, 9, 1300. [Google Scholar] [CrossRef]
- Carlson, S.K.; Erickson, D.L.; Wilson, E. Staphylococcus aureus metal acquisition in the mastitic mammary gland. Microb. Pathog. 2020, 144, 104179. [Google Scholar] [CrossRef] [PubMed]
- Sabino, Y.N.V.; Cotter, P.D.; Mantovani, H.C. Anti-virulence compounds against Staphylococcus aureus associated with bovine mastitis: A new therapeutic option? Microbiol. Res. 2023, 271, 127345. [Google Scholar] [CrossRef] [PubMed]
- Touati, A.; Ibrahim, N.A.; Idres, T. Disarming Staphylococcus aureus: Review of Strategies Combating This Resilient Pathogen by Targeting Its Virulence. Pathogens 2025, 14, 386. [Google Scholar] [CrossRef] [PubMed]
- Debruyn, E.; Ghumman, N.Z.; Peng, J.; Tiwari, H.K.; Gogoi-Tiwari, J. Alternative approaches for bovine mastitis treatment: A critical review of emerging strategies, their effectiveness and limitations. Res. Vet. Sci. 2025, 185, 105557. [Google Scholar] [CrossRef]
- Pérez, V.K.C.; da Costa, G.M.; Guimarães, A.S.; Heinemann, M.B.; Lage, A.P.; Dorneles, E.M.S. Relationship between virulence factors and antimicrobial resistance in Staphylococcus aureus from bovine mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 792–802. [Google Scholar] [CrossRef]
- Algammal, A.M.; Hetta, H.F.; Elkelish, A.; Alkhalifah, D.H.H.; Hozzein, W.N.; Batiha, G.E.-S.; El Nahhas, N.; Mabrok, M.A. Methicillin-Resistant Staphylococcus aureus (MRSA): One Health Perspective Approach to the Bacterium Epidemiology, Virulence Factors, Antibiotic-Resistance, and Zoonotic Impact. Infect. Drug Resist. 2020, 13, 3255–3265. [Google Scholar] [CrossRef]
- Abebe, A.A.; Birhanu, A.G. Methicillin Resistant Staphylococcus aureus: Molecular Mechanisms Underlying Drug Resistance Development and Novel Strategies to Combat. Infect. Drug Resist. 2023, 16, 7641–7662. [Google Scholar] [CrossRef]
- Akhtar, M.; Naqvi, S.U.-A.-S.; Liu, Q.; Pan, H.; Ma, Z.; Kong, N.; Chen, Y.; Shi, D.; Kulyar, M.F.-A.; Khan, J.A.; et al. Short Chain Fatty Acids (SCFAs) Are the Potential Immunomodulatory Metabolites in Controlling Staphylococcus aureus-Mediated Mastitis. Nutrients 2022, 14, 3687. [Google Scholar] [CrossRef]
- Pedersen, R.R.; Krömker, V.; Bjarnsholt, T.; Dahl-Pedersen, K.; Buhl, R.; Jørgensen, E. Biofilm Research in Bovine Mastitis. Front. Vet. Sci. 2021, 8, 656810. [Google Scholar] [CrossRef]
- Vargová, M.; Zigo, F.; Výrostková, J.; Farkašová, Z.; Rehan, I.F. Biofilm-Producing Ability of Staphylococcus aureus Obtained from Surfaces and Milk of Mastitic Cows. Vet. Sci. 2023, 10, 386. [Google Scholar] [CrossRef]
- Fidelis, C.E.; Orsi, A.M.; Freu, G.; Gonçalves, J.L.; Santos, M.V.D. Biofilm Formation and Antimicrobial Resistance of Staphylococcus aureus and Streptococcus uberis Isolates from Bovine Mastitis. Vet. Sci. 2024, 11, 170. [Google Scholar] [CrossRef]
- Cáceres, M.E.; Ledesma, M.M.; Lombarte Serrat, A.; Vay, C.; Sordelli, D.O.; Giacomodonato, M.N.; Buzzola, F.R. Growth conditions affect biofilms of Staphylococcus aureus producing mastitis: Contribution of MALDI-TOF-MS to strain characterization. Curr. Res. Microb. Sci. 2021, 2, 100073. [Google Scholar] [CrossRef]
- Song, M.; Tang, Q.; Ding, Y.; Tan, P.; Zhang, Y.; Wang, T.; Zhou, C.; Xu, S.; Lyu, M.; Bai, Y.; et al. Staphylococcus aureus and biofilms: Transmission, threats, and promising strategies in animal husbandry. J. Anim. Sci. Biotechnol. 2024, 15, 44. [Google Scholar] [CrossRef]
- Wang, D.; Wang, L.; Liu, Q.; Zhao, Y. Virulence factors in biofilm formation and therapeutic strategies for Staphylococcus aureus: A review. Anim. Zoonoses, 2024; in press. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, X.; Feng, X.; Shang, X.; Liu, Q.; Zhang, N.; Yang, H. Molecular characteristics of Staphylococcus aureus strains isolated from subclinical mastitis of water buffaloes in Guangdong Province, China. Front. Vet. Sci. 2023, 10, 1177302. [Google Scholar] [CrossRef]
- Xu, D.; Hu, G.; Luo, J.; Cheng, J.; Wu, D.; Cheng, L.; Huang, X.; Fu, S.; Liu, J. Staphylococcus aureus induces mitophagy to promote its survival within bovine mammary epithelial cells. Vet. Microbiol. 2023, 280, 109697. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, H.; Li, F.; Zhang, Y.; Yang, J.; Shen, Y.; Hu, N.; Zou, Q.; Qin, L.; Zeng, H.; et al. Staphylococcus aureus induces mitophagy via the HDAC11/IL10 pathway to sustain intracellular survival. J. Transl. Med. 2025, 23, 156. [Google Scholar] [CrossRef]
- Esemu, S.N.; Nya’Nying, S.F.; Ndip, L.M.; Bessong, P.O.; Tanih, N.F.; Smith, S.I.; Ndip, R.N. Isolation and characterization of methicillin-resistant Staphylococcus aureus from bovine mastitis in North West Cameroon: Public health implications. BMC Res. Notes 2024, 17, 389. [Google Scholar] [CrossRef]
- Eidaroos, N.H.; Algammal, A.M.; Mohamaden, W.I.; Alenzi, A.M.; Alghamdi, S.; Kabrah, A.; El-Mahallawy, H.S.; Eid, H.M.; Algwad, A.A.; Asfor, S.A.; et al. Virulence traits, agr typing, multidrug resistance patterns, and biofilm ability of MDR Staphylococcus aureus recovered from clinical and subclinical mastitis in dairy cows. BMC Microbiol. 2025, 25, 155. [Google Scholar] [CrossRef]
- Gad, W.A.; Osman, S.A.; Abd El-Razik, K.A.; Soror, A.H.; Fouad, E.A. A novel diagnostic technique for diagnosis of Staphylococcus aureus subclinical mastitis using gold nanoparticle-based ELISA. Open Vet. J. 2024, 14, 3388–3396. [Google Scholar] [CrossRef]
- Getahun, D.D.; Tarekegn, H.T.; Azene, B.T.; Abebe, L.T.; Belete, M.A.; Tessema, T.S. Virulence genes and antibiotic resistance profiling of staphylococcus species isolated from mastitic dairy cows in and around Bahir dar, Ethiopia. BMC Microbiol. 2025, 25, 210. [Google Scholar] [CrossRef]
- Keinprecht, H.; Irimaso, E.; Rosel, A.C.; Stessl, B.; Ntakirutimana, C.; Marek, L.; Fischer, O.W.; Szostak, M.P.; Zöchbauer, J.; Wittek, T.; et al. Diversity of Staphylococcus aureus associated with mastitis from dairy cows in Rwanda. J. Glob. Antimicrob. Resist. 2024, 36, 326–335. [Google Scholar] [CrossRef]
- Jawad, A.; Dagman, S.N.; Yaseen, M.M.; Al-Karagoly, H. Phylogenetic analysis of Staphylococcus aureus enterotoxin A gene in Iraqi breed cows with bovine mastitis: Implications for disease management. Open Vet. J. 2024, 14, 1644–1657. [Google Scholar] [CrossRef]
- Islam, M.M.; Hossain, M.I.; Islam, M.S.; Azam, M.G.; Sultana, S. Prevalence, antibiotic resistance patterns, and virulence factors of Staphylococcus aureus isolates associated with bovine mastitis in northern Bangladesh. Heliyon 2025, 11, e42107. [Google Scholar] [CrossRef]
- Zhang, N.; Hou, L.; Li, D.; Lan, W.; Zhao, Y.; Sun, X. Establishment and Application of Duplex Recombinase-Aided Amplification Combined with Lateral Flow Dipsticks for Rapid and Simultaneous Visual Detection of Klebsiella pneumoniae and Staphylococcus aureus in Milk. Foods 2025, 14, 573. [Google Scholar] [CrossRef]
- Hemati, Z.; Abdolmohammadi Khiav, L.; Zahmatkesh, A. Virulence genes and antibiotic resistance profiles of Staphylococcus aureus isolated from bovine mastitis milk samples. Iran. J. Vet. Res. 2023, 24, 258–264. [Google Scholar] [CrossRef]
- Shahzad, M.A.; Yousaf, A.; Ahsan, A.; Irshad, H.; Riaz, A.; Khan, A.; Ullah, I.; Sattar, S.; Bostan, N.; Javed, S. Virulence and resistance profiling of Staphylococcus aureus isolated from subclinical bovine mastitis in the Pakistani Pothohar region. Sci. Rep. 2024, 14, 14569. [Google Scholar] [CrossRef]
- Haq, I.U.; Kamal, M.; Swelum, A.A.; Khan, S.; los Ríos-Escalante, P.R.D.; Usman, T. Alarming multidrug resistance in Staphylococcus aureus isolated from raw milk of cows with subclinical mastitis: Antibiotic resistance patterns and occurrence of selected resistance genes. PLoS ONE 2024, 19, e0301200. [Google Scholar] [CrossRef]
- Shah, A.U.; Ali Khan, J.; Avais, M.; Zaman, S.H.; Munir, Z.; Abbas, S.; Tariq, M.; ur Rahman, M.; Tariq, F.; Nawaz, S.; et al. Prevalence and chemotherapy of Staphylococcus aureus mastitis in dairy cattle. PLoS ONE 2025, 20, e0315480. [Google Scholar] [CrossRef]
- Camsing, A.; Phetburom, N.; Chopjitt, P.; Pumhirunroj, B.; Patikae, P.; Watwiengkam, N.; Yongkiettrakul, S.; Kerdsin, A.; Boueroy, P. Occurrence of antimicrobial-resistant bovine mastitis bacteria in Sakon Nakhon, Thailand. Vet. World 2024, 17, 1202–1209. [Google Scholar] [CrossRef]
- Son, H.M.; Duc, H.M. Prevalence and Phage-Based Biocontrol of Methicillin-Resistant Staphylococcus aureus Isolated from Raw Milk of Cows with Subclinical Mastitis in Vietnam. Antibiotics 2024, 13, 638. [Google Scholar] [CrossRef]
- Naranjo-Lucena, A.; Becker, P.; Madigan, G.; Cupial, R.; Byrne, B.; Johnson, A. Longitudinal Patterns in the Isolation and Antimicrobial Resistance of Bovine Mastitis-Causing Bacteria in Ireland. Antibiotics 2025, 14, 243. [Google Scholar] [CrossRef]
- Fusar Poli, S.; Locatelli, C.; Monistero, V.; Freu, G.; Cremonesi, P.; Castiglioni, B.; Lecchi, C.; Longheu, C.M.; Tola, S.; Guaraglia, A.; et al. Staphylococcus aureus and methicillin-resistant staphylococci and mammaliicocci in the bulk tank milk of dairy cows from a livestock-dense area in northern Italy. Res. Vet. Sci. 2025, 182, 105482. [Google Scholar] [CrossRef]
- Dobrut, A.; Siemińska, I.; Sroka-Oleksiak, A.; Drożdż, K.; Sobońska, J.; Mroczkowska, U.; Brzychczy-Włoch, M. Molecular and phenotypic identification of bacterial species isolated from cows with mastitis from three regions of Poland. BMC Vet. Res. 2024, 20, 193. [Google Scholar] [CrossRef]
- Zalewska, M.; Brzozowska, P.; Rzewuska, M.; Kawecka-Grochocka, E.; Urbańska, D.M.; Sakowski, T.; Bagnicka, E. The quality and technological parameters of milk obtained from dairy cows with subclinical mastitis. J. Dairy Sci. 2025, 108, 1285–1300. [Google Scholar] [CrossRef]
- Hutu, I.; Lungu, B.C.; Spataru, I.I.; Torda, I.; Iancu, T.; Barrow, P.A.; Mircu, C. Microbiological and Molecular Investigation of Antimicrobial Resistance in Staphylococcus aureus Isolates from Western Romanian Dairy Farms: An Epidemiological Approach. Animals 2024, 14, 2266. [Google Scholar] [CrossRef]
- Aguirre-Sánchez, J.R.; Castro-del Campo, N.; Medrano-Félix, J.A.; Martínez-Torres, A.O.; Chaidez, C.; Querol-Audi, J.; Castro-del Campo, N. Genomic insights of S. aureus associated with bovine mastitis in a high livestock activity region of Mexico. J. Vet. Sci. 2024, 25, e42. [Google Scholar] [CrossRef]
- Moreno, J.; Diana, L.; Martínez, M.; Iribarnegaray, V.; Puentes, R. Comprehensive analysis of antimicrobial resistance, biofilm formation and virulence factors of staphylococci isolated from bovine mastitis. Heliyon 2025, 11, e42749. [Google Scholar] [CrossRef]
- Leon, M.; Rubin, J.; Raverty, S.; Ghosh, K. Frequency and antimicrobial susceptibility of Staphylococcus aureus isolated from clinical bovine mastitis cases in British Columbia, Canada. J. Vet. Diagn. Investig. 2024, 37, 10406387241306096. [Google Scholar] [CrossRef]
- Singh, A.; Somula, H.; Wieland, M. A retrospective cohort study investigating the association of postcalving intramammary infection and milk yield, somatic cell count, clinical mastitis, and culling risk in first-lactation dairy cows. J. Dairy Sci. 2025, 108, 4234–4247. [Google Scholar] [CrossRef]
- Neri, T.A.N.; Park, H.; Kang, S.; Baek, S.H.; Nam, I.S. Comparative Antimicrobial Resistance and Prevalence of Methicillin Resistance in Coagulase-Positive Staphylococci from Conventional and Organic Dairy Farms in South Korea. Antibiotics 2024, 13, 617. [Google Scholar] [CrossRef] [PubMed]
- Eleodoro, J.I.; Muraga, L.; Vanot, R.L.; Fagnani, R. Identification and antimicrobial susceptibility of milk isolates from cows with subclinical mastitis in the northwest of Paraná State, Brazil. Vet. Ital. 2023, 59, 71–81. [Google Scholar] [CrossRef]
- Bonsaglia, E.C.R.; Rossi, B.F.; Possebon, F.S.; Silva, N.C.C.; Gonçalves, J.L.; Castilho, I.G.; Fernandes Junior, A.; dos Santos, M.V.; Rall, V.L.M. In Vitro Adhesion and Invasion Rates of Staphylococcus aureus Isolated from Mastitic Cows Are Modulated by the agr System and MSCRAMM Genes. Vet. Sci. 2025, 12, 270. [Google Scholar] [CrossRef]
- Sharifi, A.; Mahmoudi, P.; Sobhani, K. The prevalence of adhesion and biofilm genes in Staphylococcus aureus isolates from bovine mastitis: A comprehensive meta-analysis. Vet. Med. Sci. 2024, 10, e31378. [Google Scholar] [CrossRef]
- Rocha, G.D.; de Simoni Gouveia, J.J.; da Costa, M.M.; Soares, R.A.N.; Gouveia, G.V. Resistance and virulence in Staphylococcus aureus by whole-genome sequencing: A comparative approach in blaZ-positive isolates. Braz. J. Microbiol. 2024, 55, 955–967. [Google Scholar] [CrossRef]
- Na, S.; Intanon, M.; Srithanasuwan, A.; Chaisri, W.; Suriyasathaporn, W. Evidence of vancomycin-resistant Staphylococcus aureus, multidrug-resistant S. aureus, and Enterococcus faecium-causing mastitis in Thailand and Cambodia. Vet. World 2025, 18, 202–209. [Google Scholar] [CrossRef]
- Tartor, Y.H.; Enany, M.E.; Ismail, N.I.; El-Demerdash, A.S.; Eidaroos, N.H.; Algendy, R.M.; Mahmmod, Y.; Elsohaby, I. Vancomycin-resistant Staphylococcus aureus endangers Egyptian dairy herds. Sci. Rep. 2024, 14, 30606. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wang, Y.; Bai, R.; Pei, X.; Xu, H.; Zhu, K.; Wu, C. Antimicrobial resistance profiles of common mastitis pathogens on large Chinese dairy farms. JDS Commun. 2023, 5, 185–189. [Google Scholar] [CrossRef]
- Kang, H.J.; You, J.-Y.; Kim, S.H.; Moon, J.-S.; Kim, H.-Y.; Kim, J.-M.; Lee, Y.J.; Kang, H.-M. Characteristics of methicillin-resistant Staphylococcus aureus isolates from bovine mastitis milk in South Korea: Molecular characteristics, biofilm, virulence, and antimicrobial resistance. Microbiol. Spectr. 2024, 12, e01197-24. [Google Scholar] [CrossRef]
- Karell, J.; Petzl, W.; Gangl, A.; Huber-Schlenstedt, R.; Sorge, U.S. Changes in antimicrobial resistance of Staphylococcus aureus in bovine quarter milk samples from southern Germany between 2012 and 2022. J. Dairy Sci. 2024, 107, 3802–3812. [Google Scholar] [CrossRef]
- Nesaraj, J.; Grinberg, A.; Laven, R.; Chanyi, R.; Altermann, E.; Bandi, C.; Biggs, P.J. The Host Adaptation of Staphylococcus aureus to Farmed Ruminants in New Zealand, With Special Reference to Clonal Complex 1. Environ. Microbiol. Rep. 2025, 17, e70087. [Google Scholar] [CrossRef] [PubMed]
- Aouadhi, C.; Jouini, A.; Maaroufi, K.; Maaroufi, A. Antibacterial Effect of Eight Essential Oils against Bacteria Implicated in Bovine Mastitis and Characterization of Primary Action Mode of Thymus capitatus Essential Oil. Antibiotics 2024, 13, 237. [Google Scholar] [CrossRef]
- Roadcap, E.; Lichtenwalner, A.; Kennedy-Wade, B.; Adjapong, G.; Chakrawarti, A.; De Sant’Anna, F.M.; Barlow, J.W. Whole genome sequencing identifies exotoxin and antimicrobial resistance profiles of Staphylococcus aureus from Maine dairy farms. BMC Vet. Res. 2025, 21, 154. [Google Scholar] [CrossRef] [PubMed]
- Chakrawarti, A.; Casey, C.L.; Burk, A.; Mugabi, R.; Ochoa, A.; Barlow, J.W. An observational study demonstrates human-adapted Staphylococcus aureus strains have a higher frequency of antibiotic resistance compared to cattle-adapted strains isolated from dairy farms making farmstead cheese. BMC Vet. Res. 2024, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kim, D.; Jeon, H.; Somasundaram, P.; Soundrarajan, N.; Park, C. Bactericidal activities and biochemical features of 16 antimicrobial peptides against bovine-mastitis causative pathogens. Vet. Res. 2024, 55, 150. [Google Scholar] [CrossRef]
- Rahman, M.R.T.; Guay, L.-D.; Fliss, I.; Biron, E. Structure–Activity Study of the Antimicrobial Lipopeptide Humimycin A and Screening Against Multidrug-Resistant Staphylococcus aureus. Antibiotics 2025, 14, 385. [Google Scholar] [CrossRef]
- Michira, L.; Kagira, J.; Maina, N.; Waititu, K.; Kiboi, D.; Ongera, E.; Ngotho, M. Prevalence of subclinical mastitis, associated risk factors and antimicrobial susceptibility pattern of bacteria isolated from milk of dairy cattle in Kajiado Central sub-county, Kenya. Vet. Med. Sci. 2023, 9, 2885–2892. [Google Scholar] [CrossRef]
- Shahid, M.; Hussain, R.; Nawaz, Z.; Aslam, B.; Ahmad, M.Z.; Siddique, A.B.; Ahsan, H.; Fatima, A.; Khan, I.; Mustafa, B.; et al. Occurrence of Virulence Genes among Methicillin-Resistant Staphylococcus aureus Isolated from Subclinical Bovine Mastitis. ACS Omega 2023, 8, 38111–38117. [Google Scholar] [CrossRef]
- Peña-Mosca, F.; Dean, C.; Machado, V.; Fernandes, L.; Pinedo, P.; Doster, E.; Heins, B.; Sharpe, K.; Ray, T.; Feijoo, V.; et al. Investigation of intramammary infections in primiparous cows during early lactation on organic dairy farms. J. Dairy Sci. 2023, 106, 9377–9392. [Google Scholar] [CrossRef]
- Wodaje, A.; Belete, M.A.; Menkir, A.S.; Zegeye, Z.B.; Yihunie, F.B. Detection of Virulence Genes and Antimicrobial Susceptibility Profiles of Staphylococcus aureus Isolates From Bovine Mastitis in Chagni, Northwestern Ethiopia. Vet. Med. Int. 2025, 2025, 6473601. [Google Scholar] [CrossRef]
- Abd El-Razik, K.A.; Arafa, A.A.; Fouad, E.A.; Soror, A.H.; Abdalhamed, A.M.; Elgioushy, M. Phenotypic and genotypic characterization of erythromycin-resistant Staphylococcus aureus isolated from bovine subclinical mastitis in Egypt. Vet. World 2023, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Klassen, A.; Dittmar, K.; Schulz, J.; Einax, E.; Donat, K. Estimation of the performance of two real-time polymerase chain reaction assays for detection of Staphylococcus aureus, Streptococcus agalactiae, and Streptococcus dysgalactiae in pooled milk samples in a field study. J. Dairy Sci. 2023, 106, 9228–9243. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hamid, M.I.; El-Tarabili, R.M.; Bahnass, M.M.; Alshahrani, M.A.; Saif, A.; Alwutayd, K.M.; Safhi, F.A.; Mansour, A.T.; Alblwi, N.A.N.; Ghoneim, M.M.; et al. Partnering essential oils with antibiotics: Proven therapies against bovine Staphylococcus aureus mastitis. Front. Cell. Infect. Microbiol. 2023, 13, 1265027. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Mi, S.; Dari, G.; Zhang, Z.; Chen, S.; Yu, Y. Ferroptosis-Related Genes as Molecular Markers in Bovine Mammary Epithelial Cells Challenged with Staphylococcus aureus. Int. J. Mol. Sci. 2025, 26, 2506. [Google Scholar] [CrossRef]
- Doghri, I.; Jacques, M.; Nichols, S.; Roy, J.-P.; Dufour, S. Visualization of Staphylococcus aureus in the bovine mammary gland by fluorescence in situ hybridization. Res. Vet. Sci. 2025, 189, 105634. [Google Scholar] [CrossRef]
- Penati, M.; Ulloa, F.; Locatelli, C.; Monistero, V.; Pavesi, L.F.; Santandrea, F.; Piccinini, R.; Moroni, P.; Bronzo, V.; Addis, M.F. An update on non-aureus staphylococci and mammaliicocci in cow milk: Unveiling the presence of Staphylococcus borealis and Staphylococcus rostri by MALDI-TOF MS. Vet. Res. Commun. 2024, 48, 2555–2561. [Google Scholar] [CrossRef]
- Lopes, T.; Fidelis, C.E.; Silva, A.T.F.; Mota, R.A.; Rall, V.L.M.; dos Santos, M.V.; Gonçalves, J.L. MALDI–TOF bacterial subtyping for rapid detection of biomarkers in Staphylococcus aureus from subclinical bovine mastitis. J. Appl. Microbiol. 2023, 134, lxad249. [Google Scholar] [CrossRef]
- Tamminen, L.-M.; Dahlberg, J. Evaluation of an automatic image classifier for analysis of bacterial growth on a multiple-agar plate developed for bovine mastitis. PLoS ONE 2025, 20, e0318698. [Google Scholar] [CrossRef]
- Fonseca, M.; Kurban, D.; Roy, J.-P.; Santschi, D.E.; Molgat, E.; Dufour, S. Usefulness of differential somatic cell count for udder health monitoring: Association of differential somatic cell count and somatic cell score with quarter-level milk yield and milk components. J. Dairy Sci. 2025, 108, 3900–3916. [Google Scholar] [CrossRef]
- Kajdanek, A.; Kluska, M.; Matusiak, R.; Kazimierczak, J.; Dastych, J. A Rapid and Inexpensive PCR Test for Mastitis Diagnosis Based on NGS Data. Pathogens 2024, 13, 423. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.C.; Soares, A.C.; Popolin-Neto, M.; Paulovich, F.V.; Oliveira, O.N.; Mattoso, L.H.C. Detection of Staphylococcus aureus in milk samples using impedance spectroscopy and data processing with information visualization techniques and multidimensional calibration space. Sens. Actuators Rep. 2022, 4, 100083. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, X.; He, Z.; Feng, Y.; Liu, G. Accurate detection of dairy cow mastitis with deep learning technology: A new and comprehensive detection method based on infrared thermal images. Anim. Int. J. Anim. Biosci. 2022, 16, 100646. [Google Scholar] [CrossRef] [PubMed]
- Zaninelli, M.; Redaelli, V.; Luzi, F.; Bronzo, V.; Mitchell, M.; Dell’Orto, V.; Bontempo, V.; Cattaneo, D.; Savoini, G. First Evaluation of Infrared Thermography as a Tool for the Monitoring of Udder Health Status in Farms of Dairy Cows. Sensors 2018, 18, 862. [Google Scholar] [CrossRef]
- Porter, I.R.; Wieland, M.; Basran, P.S. Feasibility of the use of deep learning classification of teat-end condition in Holstein cattle. J. Dairy Sci. 2021, 104, 4529–4536. [Google Scholar] [CrossRef] [PubMed]
- Garcia, B.L.N.; Martins, C.M.d.M.R.; Porto, L.F.; Nobrega, D.B.; dos Santos, M.V. Accuracy of an AI-based automated plate reading mobile application for the identification of clinical mastitis-causing pathogens in chromogenic culture media. Sci. Rep. 2024, 14, 1208. [Google Scholar] [CrossRef]
- Kotlarz, K.; Mielczarek, M.; Biecek, P.; Wojdak-Maksymiec, K.; Suchocki, T.; Topolski, P.; Jagusiak, W.; Szyda, J. An Explainable Deep Learning Classifier of Bovine Mastitis Based on Whole-Genome Sequence Data—Circumventing the p >> n Problem. Int. J. Mol. Sci. 2024, 25, 4715. [Google Scholar] [CrossRef]
- Krolitzki, E.; Schwaminger, S.P.; Pagel, M.; Ostertag, F.; Hinrichs, J.; Berensmeier, S. Current practices with commercial scale bovine lactoferrin production and alternative approaches. Int. Dairy J. 2022, 126, 105263. [Google Scholar] [CrossRef]
- Bobbo, T.; Matera, R.; Pedota, G.; Manunza, A.; Cotticelli, A.; Neglia, G.; Biffani, S. Exploiting machine learning methods with monthly routine milk recording data and climatic information to predict subclinical mastitis in Italian Mediterranean buffaloes. J. Dairy Sci. 2023, 106, 1942–1952. [Google Scholar] [CrossRef]
- Mitsunaga, T.M.; Nery Garcia, B.L.; Pereira, L.B.R.; Costa, Y.C.B.; da Silva, R.F.; Delbem, A.C.B.; dos Santos, M.V. Current Trends in Artificial Intelligence and Bovine Mastitis Research: A Bibliometric Review Approach. Anim. Open Access J. 2024, 14, 2023. [Google Scholar] [CrossRef]
- Esener, N.; Maciel-Guerra, A.; Giebel, K.; Lea, D.; Green, M.J.; Bradley, A.J.; Dottorini, T. Mass spectrometry and machine learning for the accurate diagnosis of benzylpenicillin and multidrug resistance of Staphylococcus aureus in bovine mastitis. PLOS Comput. Biol. 2021, 17, e1009108. [Google Scholar] [CrossRef]
- Beck, K.L.; Haiminen, N.; Agarwal, A.; Carrieri, A.P.; Madgwick, M.; Kelly, J.; Pylro, V.; Kawas, B.; Wiedmann, M.; Ganda, E. Development and evaluation of statistical and artificial intelligence approaches with microbial shotgun metagenomics data as an untargeted screening tool for use in food production. mSystems 2024, 9, e00840-24. [Google Scholar] [CrossRef] [PubMed]
- Neethirajan, S.; Kemp, B. Digital Twins in Livestock Farming. Animals 2021, 11, 1008. [Google Scholar] [CrossRef] [PubMed]
- Rowe, S.M.; Zhang, E.; Godden, S.M.; Vasquez, A.K.; Nydam, D.V. Comparison of a machine learning model with a conventional rule-based selective dry cow therapy algorithm for detection of intramammary infections. J. Dairy Sci. 2025, 108, 760–772. [Google Scholar] [CrossRef]
- Michielon, A.; Litta, P.; Bonelli, F.; Don, G.; Farisè, S.; Giannuzzi, D.; Milanesi, M.; Pietrucci, D.; Vezzoli, A.; Cecchinato, A.; et al. Mind the Step: An Artificial Intelligence-Based Monitoring Platform for Animal Welfare. Sensors 2024, 24, 8042. [Google Scholar] [CrossRef]
- Voogt, A.M.; Schrijver, R.S.; Temürhan, M.; Bongers, J.H.; Sijm, D.T.H.M. Opportunities for Regulatory Authorities to Assess Animal-Based Measures at the Slaughterhouse Using Sensor Technology and Artificial Intelligence: A Review. Anim. Open Access J. 2023, 13, 3028. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Dhand, N.K.; Wang, Z.; Hu, K.; Thomson, P.C.; House, J.K.; Khatkar, M.S. Review of applications of deep learning in veterinary diagnostics and animal health. Front. Vet. Sci. 2025, 12, 1511522. [Google Scholar] [CrossRef]
- van der Voort, M.; Jensen, D.; Kamphuis, C.; Athanasiadis, I.N.; Vries, A.D.; Hogeveen, H. Invited review: Toward a common language in data-driven mastitis detection research. J. Dairy Sci. 2021, 104, 10449–10461. [Google Scholar] [CrossRef]
- Majumder, S.; Sackey, T.; Viau, C.; Park, S.; Xia, J.; Ronholm, J.; George, S. Genomic and phenotypic profiling of Staphylococcus aureus isolates from bovine mastitis for antibiotic resistance and intestinal infectivity. BMC Microbiol. 2023, 23, 43. [Google Scholar] [CrossRef]
- Mzee, T.; Kumburu, H.; Kazimoto, T.; Leekitcharoenphon, P.; van Zwetselaar, M.; Masalu, R.; Mlaganile, T.; Sonda, T.; Wadugu, B.; Mushi, I.; et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk and Humans in Eastern Tanzania: Genetic Diversity and Inter-Host Transmission. Microorganisms 2023, 11, 1505. [Google Scholar] [CrossRef]
- Rasool, Z.; Noreen, H.; Anjum, A.; Rizvi, A.; Rabaan, A.A.; Halwani, M.A.; Sabour, A.A.; Aljeldah, M.; Shammari, B.R.A.; Alhajri, S.M.; et al. Genotypic and Phenotypic Characterization of Erythromycin-Resistant Staphylococcus aureus Isolated from Bovine Mastitis and Humans in Close Contact. Trop. Med. Infect. Dis. 2022, 8, 26. [Google Scholar] [CrossRef]
- Rodríguez, M.F.; Gomez, A.P.; Parra-Giraldo, C.M.; Ceballos-Garzon, A. Proteomics and Genetic Approaches Elucidate the Circulation of Low Variability Staphylococcus aureus Strains on Colombian Dairy Farms. Microb. Ecol. 2023, 86, 2320–2329. [Google Scholar] [CrossRef]
- Exel, C.E.; Gerritsen, K.; Spaninks, M.; Duim, B.; Koop, G.; Benedictus, L. Association of Staphylococcus aureus genotypes with milk or colonization of extramammary sites in Dutch dairy cattle indicates strain variation in reservoirs for intramammary infections. Res. Vet. Sci. 2023, 154, 138–144. [Google Scholar] [CrossRef]
- Fazoli, K.G.Z.; Rey, L.M.R.; Rúbio, K.A.J.; Garcia Souza, M.A.; Oliveira, H.M.d.S.; Ribeiro, D.C.; Pereira, K.R.d.J.D.; Kawamo, D.M.; Gomes, T.K.A.; da Silva, I.B.; et al. Resistance Profile of Bovine Mastitis Isolates, Presence of the mecA Gene and Identification of ESBL Producing Strains from Small Rural Dairy Properties. Anim. Open Access J. 2023, 13, 1147. [Google Scholar] [CrossRef]
- Abebe, R.; Markos, A.; Abera, M.; Mekbib, B. Incidence rate, risk factors, and bacterial causes of clinical mastitis on dairy farms in Hawassa City, southern Ethiopia. Sci. Rep. 2023, 13, 10945. [Google Scholar] [CrossRef]
- Singh, I.; Roshan, M.; Vats, A.; Behera, M.; Gautam, D.; Rajput, S.; Rana, C.; De, S. Evaluation of Virulence, Antimicrobial Resistance and Biofilm Forming Potential of Methicillin-Resistant Staphylococcus aureus (MRSA) Isolates from Bovine Suspected with Mastitis. Curr. Microbiol. 2023, 80, 198. [Google Scholar] [CrossRef]
- Sivakumar, R.; Pranav, P.S.; Annamanedi, M.; Chandrapriya, S.; Isloor, S.; Rajendhran, J.; Hegde, N.R. Genome sequencing and comparative genomic analysis of bovine mastitis-associated Staphylococcus aureus strains from India. BMC Genom. 2023, 24, 44. [Google Scholar] [CrossRef]
- Guo, W.; Qiu, M.; Pu, Z.; Long, N.; Yang, M.; Ren, K.; Ning, R.; Zhang, S.; Peng, F.; Sun, F.; et al. Geraniol-a potential alternative to antibiotics for bovine mastitis treatment without disturbing the host microbial community or causing drug residues and resistance. Front. Cell. Infect. Microbiol. 2023, 13, 1126409. [Google Scholar] [CrossRef]
- Crespi, E.; Pereyra, A.M.; Puigdevall, T.; Rumi, M.V.; Testorelli, M.F.; Caggiano, N.; Gulone, L.; Mollerach, M.; Gentilini, E.R.; Srednik, M.E. Antimicrobial resistance studies in staphylococci and streptococci isolated from cows with mastitis in Argentina. J. Vet. Sci. 2021, 23, e12. [Google Scholar] [CrossRef]
- Yang, F.; Shi, W.; Meng, N.; Zhao, Y.; Ding, X.; Li, Q. Antimicrobial resistance and virulence profiles of staphylococci isolated from clinical bovine mastitis. Front. Microbiol. 2023, 14, 1190790. [Google Scholar] [CrossRef]
- Tsutamoto, S.; Iwasaki, Y.; Shinohara, A.; Imamiya, R.; Samukawa, K.; Kawada-Matsuo, M.; Komatsuzawa, H.; Yamada, Y.; Mandokoro, K.; Iwao, H.; et al. Triterpenoid saponin from Panax ginseng increases the sensitivity of methicillin-resistant Staphylococcus aureus to β-lactam and aminoglycoside antibiotics. Microbiol. Spectr. 2024, 12, e03227-23. [Google Scholar] [CrossRef]
- Ahmadi, A.; Khezri, A.; Nørstebø, H.; Ahmad, R. A culture-, amplification-independent, and rapid method for identification of pathogens and antibiotic resistance profile in bovine mastitis milk. Front. Microbiol. 2023, 13, 1104701. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Seo, J. A Novel Strategy to Identify Endolysins with Lytic Activity against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2023, 24, 5772. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek-Łukowska, E.; Małaczewska, J.; Sowińska, P.; Szymańska, M.; Wójcik, E.A.; Siwicki, A.K. Staphylococcus aureus from Subclinical Cases of Mastitis in Dairy Cattle in Poland, What Are They Hiding? Antibiotic Resistance and Virulence Profile. Pathogens 2022, 11, 1404. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, D.; Zhang, Y.; Hu, Y.; Lu, J.; Zeng, Z.; Zeng, D. Preparation and Antimicrobial Activity of a Film-Forming Polyhexamethylene Biguanide Teat Disinfectant. Int. J. Mol. Sci. 2023, 24, 17444. [Google Scholar] [CrossRef]
- Wierzbicki, M.; Kot, M.; Lange, A.; Kalińska, A.; Gołębiewski, M.; Jaworski, S. Evaluation of the Antimicrobial, Cytotoxic, and Physical Properties of Selected Nano-Complexes in Bovine Udder Inflammatory Pathogen Control. Nanotechnol. Sci. Appl. 2024, 17, 77–94. [Google Scholar] [CrossRef]
- Gazzola, A.; Zucali, M.; Addis, M.F.; Bava, L.; Morandi, S.; Pisanu, S.; Pagnozzi, D.; Passera, A.; Brasca, M.; Piccinini, R. Assessment of Lactococcus cremoris preparations for the pre- and post-milking teat disinfection. BMC Vet. Res. 2024, 20, 432. [Google Scholar] [CrossRef]
- Bechtold, V.; Petzl, W.; Huber-Schlenstedt, R.; Sorge, U.S. Distribution of Bovine Mastitis Pathogens in Quarter Milk Samples from Bavaria, Southern Germany, between 2014 and 2023—A Retrospective Study. Animals 2024, 14, 2504. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef]
- Vidlund, J.; Gelalcha, B.D.; Gillespie, B.E.; Agga, G.E.; Schneider, L.; Swanson, S.M.; Frady, K.D.; Kerro Dego, O. Efficacy of novel staphylococcal surface associated protein vaccines against Staphylococcus aureus and non-aureus staphylococcal mastitis in dairy cows. Vaccine 2024, 42, 1247–1258. [Google Scholar] [CrossRef]
- Zengin, H.; Alkaç, Z.K.; Vezir, Y.; Tanyıldızı, S.; Korkak, F.A.; Hark, B.D.; Dagoglu, G. Evaluation of the efficacy of a new inactivated vaccine against Staphylococcus aureus, Echerchia coli and Mycoplasma bovis mastitis in cows. Vet. Ital. 2024, 60, 1–11. [Google Scholar] [CrossRef]
- Rainard, P.; Gilbert, F.B.; Germon, P.; Foucras, G. Invited review: A critical appraisal of mastitis vaccines for dairy cows. J. Dairy Sci. 2021, 104, 10427–10448. [Google Scholar] [CrossRef]
- Côté-Gravel, J.; Malouin, F. Symposium review: Features of Staphylococcus aureus mastitis pathogenesis that guide vaccine development strategies*. J. Dairy Sci. 2019, 102, 4727–4740. [Google Scholar] [CrossRef] [PubMed]
- Clabby, C.; Valldecabres, A.; Dillon, P.; O’Sullivan, K.; Arkins, S.; Flynn, J.; McCarthy, S.; Boloña, P.S. The association between somatic cell count and selective dry cow therapy, milking routine, and dry cow management practices in early-lactation cows from 21 commercial grazing dairy herds. J. Dairy Sci. 2024, 107, 7106–7120. [Google Scholar] [CrossRef] [PubMed]
- Clabby, C.; McParland, S.; Dillon, P.; Arkins, S.; Flynn, J.; Murphy, J.; Boloña, P.S. Internal teat sealants alone or in combination with antibiotics at dry-off—The effect on udder health in dairy cows in five commercial herds. Animal 2022, 16, 100449. [Google Scholar] [CrossRef]
- Izhar, M.Z.; Nawaz, M.; Yaqub, T.; Avais, M.; Anjum, A.A. In vitro characterization of probiotic potential of Lactobacillus plantarum CM49 against selected cattle mastitogens. BMC Microbiol. 2024, 24, 310. [Google Scholar] [CrossRef]
- Stafford, E.; Kot, M. Optimal reduced-mixing for an SIS infectious-disease model. J. Biol. Dyn. 2022, 16, 746–765. [Google Scholar] [CrossRef]
- Waller, K.P.; Myrenås, M.; Börjesson, S.; Kim, H.; Widerström, M.; Monsen, T.; Sandholt, A.K.S.; Östlund, E.; Cha, W. Genotypic characterization of Staphylococcus chromogenes and Staphylococcus simulans from Swedish cases of bovine subclinical mastitis. J. Dairy Sci. 2023, 106, 7991–8004. [Google Scholar] [CrossRef]
- Gharaibeh, M.H.; Abu-Qatouseh, L.F. First molecular characterization of capsule expression and antibiotic susceptibility profile of Staphylococcus aureus isolates from bovine mastitis in Jordan. Vet. World 2022, 15, 2269–2274. [Google Scholar] [CrossRef]
- Hoque, M.N.; Talukder, A.K.; Saha, O.; Hasan, M.M.; Sultana, M.; Rahman, A.A.; Das, Z.C. Antibiogram and virulence profiling reveals multidrug resistant Staphylococcus aureus as the predominant aetiology of subclinical mastitis in riverine buffaloes. Vet. Med. Sci. 2022, 8, 2631–2645. [Google Scholar] [CrossRef]
- Maisano, A.M.; Luini, M.; Gazzola, A.; Sala, L.; Vezzoli, F.; Bertocchi, L.; Lorenzi, V.; Cremonesi, P.; Castiglioni, B.; Bergagna, S.; et al. Staphylococcus aureus adlb gene is associated with high prevalence of intramammary infection in dairy herds of northern Italy: A cross-sectional study. J. Dairy Sci. 2023, 106, 3421–3435. [Google Scholar] [CrossRef]
- Miyazawa, R.; Shimoda, S.; Matsuda, K.; Tobe, R.; Ando, T.; Yoneyama, H. Characterization of Staphylococcus aureus Isolates from Bovine Mastitis and Bulk Tank Milk: First Isolation of Methicillin-Susceptible Staphylococcus aureus in Japan. Microorganisms 2022, 10, 2117. [Google Scholar] [CrossRef]
- Singha, S.; Koop, G.; Rahman, M.M.; Ceciliani, F.; Addis, M.F.; Howlader, M.M.R.; Hossain, M.K.; Piccinini, R.; Locatelli, C.; Persson, Y.; et al. Pathogen group-specific risk factors for intramammary infection in water buffalo. PLoS ONE 2024, 19, e0299929. [Google Scholar] [CrossRef] [PubMed]
- Mohammadian, F.; Rahmani, H.K.; Bidarian, B.; Khoramian, B. Isolation and evaluation of the efficacy of bacteriophages against multidrug-resistant (MDR), methicillin-resistant (MRSA) and biofilm-producing strains of Staphylococcus aureus recovered from bovine mastitis. BMC Vet. Res. 2022, 18, 406. [Google Scholar] [CrossRef] [PubMed]
- Sikora, K.; Nowacki, A.; Szweda, P.; Woziwodzka, A.; Bartoszewska, S.; Piosik, J.; Dmochowska, B. Antimicrobial, Cytotoxic and Mutagenic Activity of Gemini QAS Derivatives of 1,4:3,6-Dianhydro-l-iditol. Molecules 2022, 27, 757. [Google Scholar] [CrossRef]
- Toledo-Silva, B.; Beuckelaere, L.; De Visscher, A.; Geeroms, C.; Meyer, E.; Piepers, S.; Thiry, D.; Haesebrouck, F.; De Vliegher, S. Novel Quantitative Assay to Describe In Vitro Bovine Mastitis Bacterial Pathogen Inhibition by Non-aureus Staphylococci. Pathogens 2022, 11, 264. [Google Scholar] [CrossRef]
- Neelam; Jain, V.K.; Singh, M.; Joshi, V.G.; Chhabra, R.; Singh, K.; Rana, Y.S. Virulence and antimicrobial resistance gene profiles of Staphylococcus aureus associated with clinical mastitis in cattle. PLoS ONE 2022, 17, e0264762. [Google Scholar] [CrossRef]
- de los Santos, R.; González-Revello, Á.; Majul, L.; Umpiérrez, A.; Aldrovandi, A.; Gil, A.; Hirigoyen, D.; Zunino, P. Subclinical bovine mastitis associated with Staphylococcus spp. in eleven Uruguayan dairy farms. J. Infect. Dev. Ctries. 2022, 16, 630–637. [Google Scholar] [CrossRef]
- Ferronato, G.; Simonetto, A.; Gilioli, G.; Zecconi, A. Modeling Mastitis Risk Management Effects on Dairy Milk Yield and Global Warming Potential. Anim. Open Access J. 2024, 15, 50. [Google Scholar] [CrossRef]
- Brito, L.F.; Bedere, N.; Douhard, F.; Oliveira, H.R.; Arnal, M.; Peñagaricano, F.; Schinckel, A.P.; Baes, C.F.; Miglior, F. Review: Genetic selection of high-yielding dairy cattle toward sustainable farming systems in a rapidly changing world. Animal 2021, 15, 100292. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).