


Characterization of Antimicrobial Resistance in Campylobacter Species from Broiler Chicken Litter
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Flock Characteristics
2.2. Bacterial Isolation and Speciation
2.3. Antimicrobial Susceptibility Assays
2.4. Genomic DNA Extraction
2.5. PCR Assay to Detect AMR Genes
2.6. Conjugation Assay
2.7. Statistical Analyses
3. Results
3.1. Distribution Frequency and Minimum Inhibition Concentration (MIC) for Campylobacter spp. Isolates from 2015 and 2016
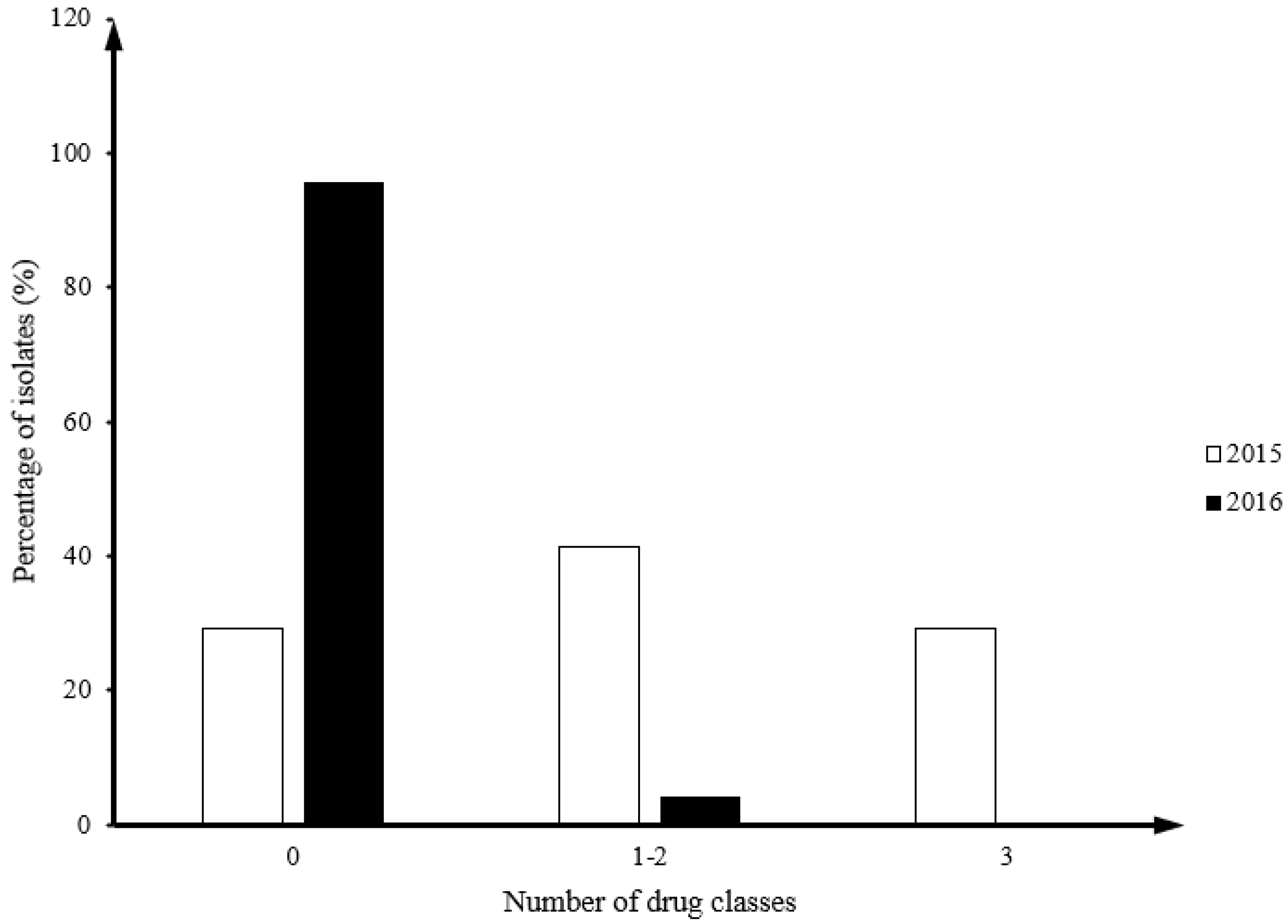
3.2. Resistance Patterns of Campylobacter Isolates from 2015 and 2016
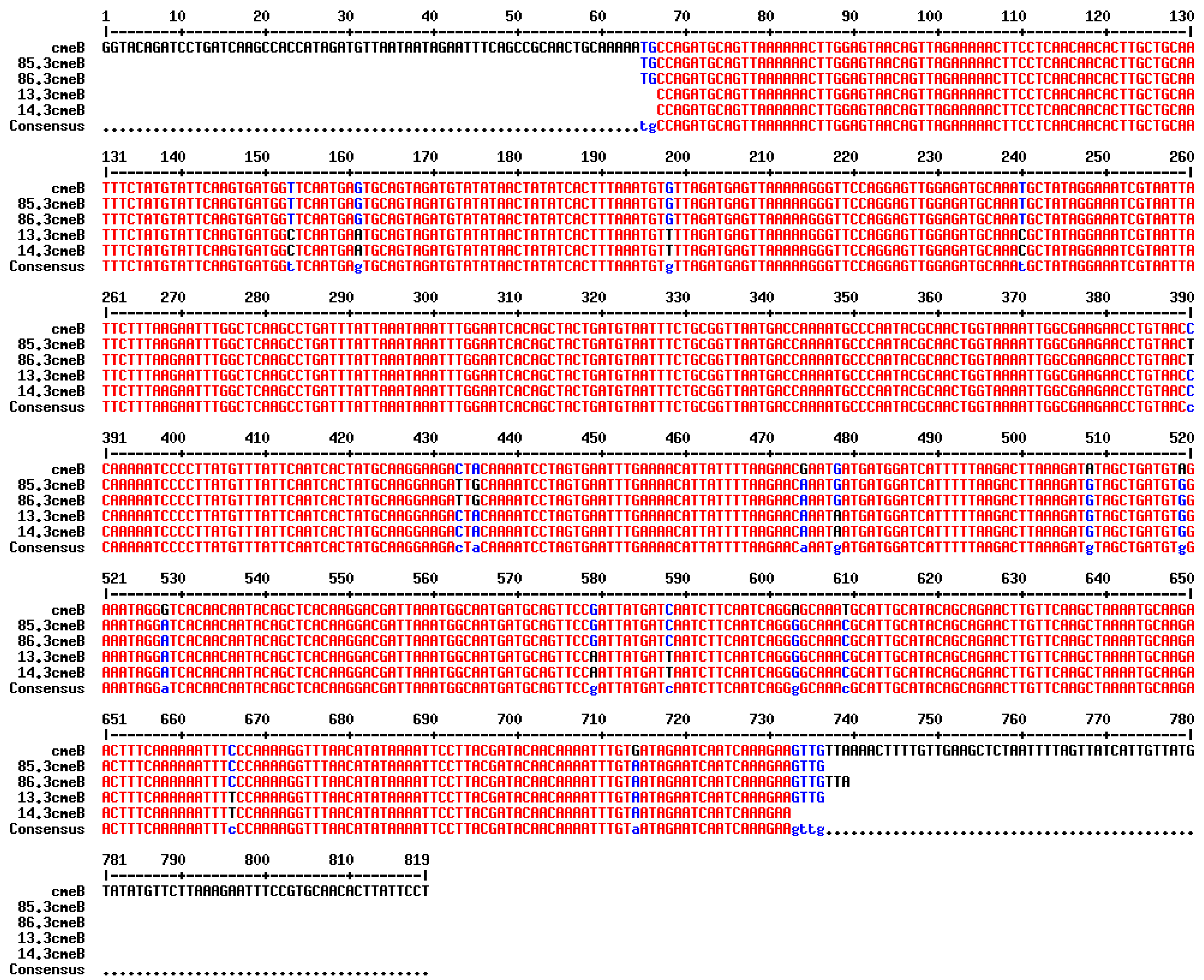
3.3. Detection of AMR Genes
3.4. Campylobacter jejuni Antimicrobial Resistance Phenotypes Transferred via In Vitro Conjugation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Luber, P.; Wagner, J.; Hahn, H.; Bartelt, E. Antimicrobial Resistance in Campylobacter jejuni and Campylobacter coli Strains Isolated in 1991 and 2001–2002 from Poultry and Humans in Berlin, Germany. Antimicrob. Agents Chemother. 2003, 47, 3825–3830. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Nelson, J.M.; Barrett, T.J.; Tauxe, R.V.; Rossiter, S.P.; Friedman, C.R.; Joyce, K.W.; Smith, K.E.; Jones, T.F.; Hawkins, M.A.; et al. Antimicrobial resistance among Campylobacter strains, United States, 1997–2001. Emerg. Infect. Dis. 2004, 10, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Canada (PHAC). Canadian Notifiable Disease Section 2015. Available online: http://diseases.canada.ca/notifiable/charts?c=pl (accessed on 15 April 2025).
- Hansson, I.; Sandberg, M.; Habib, I.; Lowman, R.; Engvall, E.O. Knowledge gaps in control of Campylobacter for prevention of campylobacteriosis. Transbound. Emerg. Dis. 2018, 65, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Butzler, J. Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect. 2004, 10, 868–876. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2010, 4, 189–200. [Google Scholar] [CrossRef]
- Di Giannatale, E.; Calistri, P.; Di Donato, G.; Decastelli, L.; Goffredo, E.; Adriano, D.; Mancini, M.E.; Galleggiante, A.; Neri, D.; Antoci, S.; et al. Thermotolerant Campylobacter spp. in chicken and bovine meat in Italy: Prevalence, level of contamination and molecular characterization of isolates. PLoS ONE 2019, 14, e0225957. [Google Scholar] [CrossRef]
- Hakeem, M.J.; Lu, X. Survival and Control of Campylobacter in Poultry Production Environment. Front. Cell. Infect. Microbiol. 2021, 10, 615049. Available online: https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2020.615049 (accessed on 13 April 2025). [CrossRef]
- Ngogang, M.P.; Ernest, T.; Kariuki, J.; Mouliom Mouiche, M.M.; Ngogang, J.; Wade, A.; van der Sande, M.A.B. Microbial Contamination of Chicken Litter Manure and Antimicrobial Resistance Threat in an Urban Area Setting in Cameroon. Antibiotics 2021, 10, 20. [Google Scholar] [CrossRef]
- Gupta, C.L.; Blum, S.E.; Kattusamy, K.; Daniel, T.; Druyan, S.; Shapira, R.; Krifucks, O.; Zhu, Y.-G.; Zhou, X.-Y.; Su, J.-Q.; et al. Longitudinal study on the effects of growth-promoting and therapeutic antibiotics on the dynamics of chicken cloacal and litter microbiomes and resistomes. Microbiome 2021, 9, 178. [Google Scholar] [CrossRef]
- Narvaez-Bravo, C.; Taboada, E.N.; Mutschall, S.K.; Aslam, M. Epidemiology of antimicrobial resistant Campylobacter spp. isolated from retail meats in Canada. Int. J. Food Microbiol. 2017, 253, 43–47. Available online: https://www.sciencedirect.com/science/article/pii/S0168160517301782 (accessed on 17 April 2025). [CrossRef]
- Rath, A.; Rautenschlein, S.; Rzeznitzeck, J.; Breves, G.; Hewicker-Trautwein, M.; Waldmann, K.H.; von Altrock, A. Impact of Campylobacter spp. on the Integrity of the Porcine Gut. Animals 2021, 11, 2742. [Google Scholar] [CrossRef]
- Altekruse, S.F.; Stern, N.J.; Fields, P.I.; Swerdlow, D.L. Campylobacter jejuni—An emerging foodborne pathogen. Emerg. Infect. Dis. 1999, 5, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lehtopolku, M.; Kotilainen, P.; Puukka, P.; Nakari, U.M.; Siitonen, A.; Eerola, E.; Huovinen, P.; Hakanen, A.J. Inaccuracy of the disk diffusion method compared with the agar dilution method for susceptibility testing of Campylobacter spp. J. Clin. Microbiol. 2012, 50, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Government of Canada. Categorization of Antimicrobial Drugs Based on Importance in Human Medicine. 2009. Available online: https://www.canada.ca/en/health-canada/services/drugs-health-products/veterinary-drugs/antimicrobial-resistance/categorization-antimicrobial-drugs-based-importance-human-medicine.html (accessed on 17 April 2025).
- Chicken Farmers of Canada. 2016 Annual Report. 2016. Available online: https://epe.lac-bac.gc.ca/100/201/300/ar_chicken_farmers/2016.pdf?nodisclaimer=1 (accessed on 17 April 2025).
- Golz, J.C.; Stingl, K. Natural Competence and Horizontal Gene Transfer in Campylobacter BT—Fighting Campylobacter Infections: Towards a One Health Approach; Backert, S., Ed.; Springer International Publishing: Cham, Switzerland, 2021; Volume 431, pp. 265–292. [Google Scholar] [CrossRef]
- Jonsdottir, I.; Given, C.; Penttinen, R.; Jalasvuori, M. Preceding Host History of Conjugative Resistance Plasmids Affects Intra- and Interspecific Transfer Potential from Biofilm. mSphere 2023, 8, e0010723. [Google Scholar] [CrossRef] [PubMed]
- Alonso-del Valle, A.; Toribio-Celestino, L.; Quirant, A.; Pi, C.T.; DelaFuente, J.; Canton, R.; Rocha, E.P.; Ubeda, C.; Peña-Miller, R.; San Millan, A. Antimicrobial resistance level and conjugation permissiveness shape plasmid distribution in clinical enterobacteria. Proc. Natl. Acad. Sci. USA 2023, 120, e2314135120. [Google Scholar] [CrossRef]
- Yang, M.; Wang, X.; Zheng, L.; Zhu, Y. Genomic analysis and antimicrobial resistance in human- and poultry-derived Campylobacter jejuni isolates from Hangzhou, China. Front. Microbiol. 2025, 16, 1599555. [Google Scholar] [CrossRef]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef]
- Tang, Y.; Fang, L.; Xu, C.; Zhang, Q. Antibiotic resistance trends and mechanisms in the foodborne pathogen, Campylobacter. Anim. Health Res. Rev. 2017, 18, 87–98. Available online: https://www.cambridge.org/core/product/3CA9C0DA6FA8AE1F0EE12AB80D372380 (accessed on 12 January 2025). [CrossRef]
- Dai, L.; Wu, Z.; Sahin, O.; Zhao, S.; Yu, E.W.; Zhang, Q. Mutation-based mechanism and evolution of the potent multidrug efflux pump RE-CmeABC in Campylobacter. Proc. Natl. Acad. Sci. USA 2024, 121, e2415823121. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. Canadian Integrated Program for Antimicrobial (CIPARS) Annual Report 2015. 2015. Available online: http://publications.gc.ca/collections/collection_2017/aspc-phac/HP2-4-2015-eng.pdf (accessed on 10 January 2025).
- Persson, S.; Olsen, K.E.P. Multiplex PCR for identification of Campylobacter coli and Campylobacter jejuni from pure cultures and directly on stool samples. J. Med. Microbiol. 2005, 54, 1043–1047. [Google Scholar] [CrossRef]
- Abdi-Hachesoo, B.; Khoshbakht, R.; Sharifiyazdi, H.; Tabatabaei, M.; Hosseinzadeh, S.; Asasi, K. Tetracycline resistance genes in Campylobacter jejuni and C. coli isolated from poultry carcasses. Jundishapur J. Microbiol. 2014, 7, 7–11. [Google Scholar] [CrossRef]
- Bacon, D.J.; Alm, R.; Burr, D.H.; Hu, L.; Kopecko, D.J.; Ewing, C.P.; Trust, T.J.; Guerry, P.; Barbieri, J.T. Involvement of a plasmid in virulence of Campylobacter jejuni 81–176. Infect. Immun. 2000, 68, 4384–4390. [Google Scholar] [CrossRef]
- Lin, J.; Michel, L.O.; Zhang, Q. CmeABC Functions as a Multidrug Efflux System in Campylobacter jejuni. Society 2002, 46, 2124–2131. [Google Scholar] [CrossRef]
- M45-A2; Performance Standards for Antimicrobial Susceptibility. Clinical and Laboratory Standards Institute (CLSI): Wayne, PL, USA, 2017.
- Meade, H.M.; Long, S.R.; Ruvkun, G.B.; Brown, S.E.; Ausubel, M. Physical and Genetic Characterization of Symbiotic and Auxotrophic Mutants of Rhizobium meliloti Induced by Transposon Tn5 Mutagenesis. J. Bacteriol. 1982, 149, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Checkley, S.; Caffrey, N.; Mainali, C.; Gow, S. Genetic Characterization of AmpC and Extended-Spectrum Beta- Lactamase Phenotypes in Escherichia coli and Salmonella From Alberta Broiler Chickens. Front. Cell. Infect. Microbiol. 2021, 11, 622195. [Google Scholar] [CrossRef] [PubMed]
- Reed, W.; Adelumola, O.; Dinku, E.; Timothy, S.; Jodie, P.L.; Zaid, A. Virulence factors and antimicrobial resistance profiles of Campylobacter isolates recovered from consecutively reused broiler litter. Microbiol. Spectr. 2023, 11, e03223. [Google Scholar] [CrossRef] [PubMed]
- Emberland, K.E.; Wensaas, K.A.; Litleskare, S.; Iversen, A.; Hanevik, K.; Langeland, N.; Rortveit, G. Clinical features of gastroenteritis during a large waterborne Campylobacter outbreak in Askøy, Norway. Infection 2022, 50, 343–354. [Google Scholar] [CrossRef]
- Bolinger, H.; Kathariou, S. The Current State of Macrolide Resistance in Campylobacter spp.: Trends and Impacts of Resistance Mechanisms. Appl. Environ. Microbiol. 2017, 83, e00416–e00417. [Google Scholar] [CrossRef]
- Lynch, C.T.; Lynch, H.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; Lucey, B. Antimicrobial resistance of Campylobacter isolates recovered from broilers in the Republic of Ireland in 2017 and 2018, an update. Br. Poult. Sci. 2020, 61, 550–556. [Google Scholar] [CrossRef]
- Dasti, J.I.; Groß, U.; Pohl, S.; Lugert, R.; Weig, M.; Schmidt-Ott, R. Role of the plasmid-encoded tet(O) gene in tetracycline-resistant clinical isolates of Campylobacter jejuni and Campylobacter coli. J. Med. Microbiol. 2007, 56, 833–837. [Google Scholar] [CrossRef]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrob. Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef]
- Zilhao, R.; Papadopoulou, B.; Courvalin, P. Occurrence of the Campylobacter resistance gene tetO in Enterococcus and Streptococcus spp. Antimicrob. Agents Chemother. 1988, 32, 1793–1796. [Google Scholar] [CrossRef] [PubMed]
- Nirdnoy, W.; Mason, C.J.; Guerry, P. Mosaic structure of a multiple-drug-resistant, conjugative plasmid from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2454–2459. [Google Scholar] [CrossRef] [PubMed]
- Marasini, D.; Karki, A.B.; Bryant, J.M.; Sheaff, R.J.; Fakhr, M.K. Molecular characterization of megaplasmids encoding the type VI secretion system in Campylobacter jejuni isolated from chicken livers and gizzards. Sci. Rep. 2020, 10, 12514. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Sahin, O.; Lin, J.; Zhang, Q. Role of the CmeABC efflux pump in the emergence of fluoroquinolone-resistant Campylobacter under selection pressure. J. Antimicrob. Chemother. 2006, 58, 1154–1159. [Google Scholar] [CrossRef]
- Ge, B.; Mcdermott, P.F.; White, D.G.; Meng, J. Role of Efflux Pumps and Topoisomerase Mutations in Fluoroquinolone Resistance in Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 2005, 49, 3347–3354. [Google Scholar] [CrossRef]
- Aksomaitiene, J.; Ramonaite, S.; Olsen, J.E.; Malakauskas, M. Prevalence of Genetic Determinants and Phenotypic Resistance to Ciprofloxacin in Campylobacter jejuni from Lithuania. Front. Microbiol. 2018, 9, 203. [Google Scholar] [CrossRef]
- Hakanen, A.; Jalava, J.; Kotilainen, P.; Jousimies-somer, H.; Siitonen, A.; Huovinen, P. gyrA Polymorphism in Campylobacter jejuni: Detection of gyrA Mutations in 162 C. jejuni Isolates by Single-Strand Conformation Polymorphism and DNA Sequencing. Antimicrob. Agents Chemother. 2002, 46, 2644–2647. [Google Scholar] [CrossRef]
- San Millan, A.; Maclean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spectr. 2017, 5, 65–79. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Gene Target (Size) | Primers | Reference |
|---|---|---|
| 16S universal ribosomal RNA sequence (1062 bp) | 16s-F: 5′-GGAGGCAGCAGTAGGGAATA-3′ 16s-R: 5′-TGACGGGCGGTGAGTACAAG-3′ | [26] |
| Aspartokinase to detect C. coli (500 bp) | CC18-F: 5′- GGTATGATTTCTACAAGCGAG-3′ CC519-R: 5′-ATAAAAGACTATCGTCGCGTG-3′ | [26] |
| Hippuricase to detect C. jejuni | hipO-F: 5′-GACTTCGTGCAGATATGGATGCTT-3′ hipO-R: 5′-GCTATAACTATCCGAAGAAGCCATCA-3′ | [26] |
| tetA gene (888 bp) | tetA_F: 5′-GTGAAACCCAACATACCCC-3′ tetA_R: 5′-GAAGGCAAGCAGGATGTAG-3′ | [27] |
| tetO gene (559 bp) | tetO_F: 5′GGCGTTTTGTTTATGTGCG-3′ tetO_R: 5′-ATGGACAACCCGACAGAAGC-3′ | [28] |
| cmeB gene (819 bp) | cmeB_F: 5′-GGTACAGATCCTGATCAAGCC-3′ cmeB_R: 5′-AGGAATAAGTGTTGCACGGAAATT-3′ | [29] |
| Year | Antimicrobial | Distribution of Isolates (Count) Across MIC Range | ||||||||||||||
| <0.015 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | >64 | ||
| Gentamicin | 1 | 40 | ||||||||||||||
| Telithromycin | 21 | 8 | 12 | |||||||||||||
| Clindamycin | 6 | 22 | 1 | 5 | 7 | |||||||||||
| Nalidixic acid | 27 | 13 | 1 | |||||||||||||
| 2015 | Ciprofloxacin | 40 | 1 | |||||||||||||
| Erythromycin | 18 | 11 | 12 | |||||||||||||
| Azithromycin | 1 | 14 | 9 | 4 | 1 | 12 | ||||||||||
| Florfenicol | 3 | 38 | ||||||||||||||
| Tetracycline | 2 | 5 | 17 | 6 | 2 | 9 | ||||||||||
| Gentamicin | 6 | 18 | ||||||||||||||
| Telithromycin | 9 | 7 | 1 | 3 | 4 | |||||||||||
| Clindamycin | 2 | 13 | 2 | 7 | ||||||||||||
| Nalidixic acid | 7 | 7 | 9 | 1 | ||||||||||||
| 2016 | Ciprofloxacin | 9 | 15 | |||||||||||||
| Erythromycin | 1 | 12 | 3 | 2 | 6 | |||||||||||
| Azithromycin | 13 | 3 | 3 | 5 | ||||||||||||
| Florfenicol | 20 | 4 | ||||||||||||||
| Tetracycline | 14 | 1 | 1 | 7 | 1 | |||||||||||
| Strains | MIC (µg/mL) | Interpretation | AMR Patterns | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AZM | CIP | CLI | ERY | FLR | GEN | NAL | TEL | TET | AZM | CIP | CLI | ERY | FLR | GEN | NAL | TEL | TET | ||
| C. jejuni ATCC 33250 a | 0.03 | 0.06 | 0.25 | <0.25 | 1 | 0.5 | ≤4 | <0.5 | <0.12 | S | S | S | S | S | S | S | S | S | Susceptible |
| 6.3 | 0.12 | 0.12 | 0.12 | 0.5 | 1 | 1 | ≤4 | 1 | >64 | S | S | S | S | S | S | S | S | R | Te |
| 85.3 | 0.12 | 0.12 | 0.12 | 0.5 | 1 | 0.5 | ≤4 | 0.5 | >64 | S | S | S | S | S | S | S | S | R | Te |
| 96.3 | 0.06 | 8 | 0.12 | 0.25 | 1 | 1 | >64 | 0.5 | >64 | S | R | S | S | S | S | R | S | R | CiNaTe |
| 13.3 | >64 | 0.12 | 16 | >64 | 1 | 1 | 8 | >8 | 0.25 | R | S | R | R | S | S | S | R | S | AzClErTl |
| 113.3 | >64 | 0.12 | 8 | >64 | 1 | 1 | 8 | >8 | 0.25 | R | S | R | R | S | S | S | R | S | AzClErTl |
| 117.3 | >64 | 0.12 | 16 | >64 | 1 | 1 | 8 | >8 | 0.25 | R | S | R | R | S | S | S | R | S | AzClErTl |
| 13.6 b | >64 | 0.12 | 8 | >64 | 1 | 0.25 | 8 | >8 | >64 | R | S | R | R | S | S | S | R | R | AzClErTlTe |
| 13.85 b | >64 | 0.12 | 8 | >64 | 1 | 0.25 | 8 | 8 | >64 | R | S | R | R | S | S | S | R | R | AzClErTlTe |
| 13.96 b | >64 | 0.12 | 8 | >64 | 2 | 0.25 | 8 | >8 | >64 | R | S | R | R | S | S | S | R | R | AzClErTlTe |
| 96.13 c | >64 | 16 | 16 | >64 | 1 | 0.5 | >64 | >8 | 0.12 | R | R | R | R | S | S | R | R | S | CiNaAzClErTl |
| 96.113 c | >64 | 16 | 16 | >64 | 2 | 0.5 | >64 | >8 | 0.25 | R | R | R | R | S | S | R | R | S | CiNaAzClErTl |
| 96.117 c | >64 | 8 | 16 | >64 | 1 | 0.25 | >64 | >8 | 0.12 | R | R | R | R | S | S | R | R | S | CiNaAzClErTl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, T.T.; Checkley, S.; Caffrey, N.; Mainali, C.; Gow, S.; Agunos, A.; Liljebjelke, K. Characterization of Antimicrobial Resistance in Campylobacter Species from Broiler Chicken Litter. Antibiotics 2025, 14, 759. https://doi.org/10.3390/antibiotics14080759
Tran TT, Checkley S, Caffrey N, Mainali C, Gow S, Agunos A, Liljebjelke K. Characterization of Antimicrobial Resistance in Campylobacter Species from Broiler Chicken Litter. Antibiotics. 2025; 14(8):759. https://doi.org/10.3390/antibiotics14080759
Chicago/Turabian StyleTran, Tam T., Sylvia Checkley, Niamh Caffrey, Chunu Mainali, Sheryl Gow, Agnes Agunos, and Karen Liljebjelke. 2025. "Characterization of Antimicrobial Resistance in Campylobacter Species from Broiler Chicken Litter" Antibiotics 14, no. 8: 759. https://doi.org/10.3390/antibiotics14080759
APA StyleTran, T. T., Checkley, S., Caffrey, N., Mainali, C., Gow, S., Agunos, A., & Liljebjelke, K. (2025). Characterization of Antimicrobial Resistance in Campylobacter Species from Broiler Chicken Litter. Antibiotics, 14(8), 759. https://doi.org/10.3390/antibiotics14080759