
Challenges of Carbapenem-Resistant Enterobacteriaceae in the Development of New β-Lactamase Inhibitors and Antibiotics



Abstract
1. Introduction
2. Carbapenem-Resistant Enterobacteriaceae and the Diversity of β-Lactamases
3. Therapeutic Strategies, β-Lactamase Inhibitors, and New Antibiotics
4. An Active and Multifaceted Clinical Research Landscape
5. An Emerging and Promising Avenue of Exploratory Research
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleming, A. Classics in Infectious Diseases: On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to Their Use in the Isolation of B. Influenzae by Alexander Fleming, Reprinted from the British Journal of Experimental Pathology 10:226–236, 1929. Rev. Infect. Dis. 1980, 2, 129–139. [Google Scholar] [PubMed]
- Hoffman, P.S. Antibacterial Discovery: 21st Century Challenges. Antibiotics 2020, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Blaskovich, M.A.; Cooper, M.A. Antibiotics in the Clinical Pipeline at the End of 2015. J. Antibiot. 2017, 70, 3–24. [Google Scholar] [CrossRef]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug Resistance: An Emerging Crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 541340. [Google Scholar] [CrossRef]
- GBD 2021 Antimicrobial Resistance Collaborators. Global Burden of Bacterial Antimicrobial Resistance 1990–2021: A Systematic Analysis with Forecasts to 2050. Lancet Lond. Engl. 2024, 404, 1199–1226. [Google Scholar] [CrossRef] [PubMed]
- Stemming the Superbug Tide. Available online: https://www.oecd.org/en/publications/stemming-the-superbug-tide_9789264307599-en.html (accessed on 11 March 2025).
- Drug-Resistant Infections: A Threat to Our Economic Future. Available online: https://www.worldbank.org/en/topic/health/publication/drug-resistant-infections-a-threat-to-our-economic-future (accessed on 11 March 2025).
- Lobanovska, M.; Pilla, G. Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future? Yale J. Biol. Med. 2017, 90, 135–145. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 2. [Google Scholar] [CrossRef]
- One Health. Available online: https://www.who.int/news-room/fact-sheets/detail/one-health (accessed on 25 March 2025).
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for Bad Bugs: Confronting the Challenges of Antibacterial Discovery. Nat. Rev. Drug Discov. 2007, 6, 29–40. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical Analysis of Antibacterial Agents in Clinical Development. Nat. Rev. Microbiol. 2020, 18, 286–298. [Google Scholar] [CrossRef]
- WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance. Available online: https://www.who.int/publications/i/item/9789240093461 (accessed on 11 March 2025).
- Lewis, K. The Science of Antibiotic Discovery. Cell 2020, 181, 29–45. [Google Scholar] [CrossRef]
- Monserrat-Martinez, A.; Gambin, Y.; Sierecki, E. Thinking Outside the Bug: Molecular Targets and Strategies to Overcome Antibiotic Resistance. Int. J. Mol. Sci. 2019, 20, 1255. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.-S.; Harnod, D.; Hsueh, P.-R. Global Threat of Carbapenem-Resistant Gram-Negative Bacteria. Front. Cell. Infect. Microbiol. 2022, 12, 823684. [Google Scholar] [CrossRef] [PubMed]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Barber, K.E.; Wagner, J.L.; Larry, R.C.; Stover, K.R. Frequency of and Risk Factors for Carbapenem-Resistant Enterobacteriaceae. J. Med. Microbiol. 2021, 70, 001286. [Google Scholar] [CrossRef]
- Lee, I.; Jo, J.-W.; Woo, H.-J.; Suk, K.T.; Lee, S.S.; Kim, B.-S. Proton Pump Inhibitors Increase the Risk of Carbapenem-Resistant Enterobacteriaceae Colonization by Facilitating the Transfer of Antibiotic Resistance Genes among Bacteria in the Gut Microbiome. Gut Microbes 2024, 16, 2341635. [Google Scholar] [CrossRef]
- Yip, A.Y.G.; King, O.G.; Omelchenko, O.; Kurkimat, S.; Horrocks, V.; Mostyn, P.; Danckert, N.; Ghani, R.; Satta, G.; Jauneikaite, E.; et al. Antibiotics Promote Intestinal Growth of Carbapenem-Resistant Enterobacteriaceae by Enriching Nutrients and Depleting Microbial Metabolites. Nat. Commun. 2023, 14, 5094. [Google Scholar] [CrossRef]
- Baek, M.S.; Kim, S.; Kim, W.-Y.; Kweon, M.-N.; Huh, J.W. Gut Microbiota Alterations in Critically Ill Patients with Carbapenem-Resistant Enterobacteriaceae Colonization: A Clinical Analysis. Front. Microbiol. 2023, 14, 1140402. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Reuter, S.; Wille, J.; Xanthopoulou, K.; Stefanik, D.; Grundmann, H.; Higgins, P.G.; Seifert, H. A Global View on Carbapenem-Resistant Acinetobacter Baumannii. mBio 2023, 14, e0226023. [Google Scholar] [CrossRef]
- Ferous, S.; Anastassopoulou, C.; Pitiriga, V.; Vrioni, G.; Tsakris, A. Antimicrobial and Diagnostic Stewardship of the Novel β-Lactam/β-Lactamase Inhibitors for Infections Due to Carbapenem-Resistant Enterobacterales Species and Pseudomonas Aeruginosa. Antibiotics 2024, 13, 285. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Zuccari, G. Recommendations to Synthetize Old and New β-Lactamases Inhibitors: A Review to Encourage Further Production. Pharmaceuticals 2022, 15, 384, Erratum in Pharmaceuticals 2022, 15, 526. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. Interplay between β-Lactamases and New β-Lactamase Inhibitors. Nat. Rev. Microbiol. 2019, 17, 295–306. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-Resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid Detection of Carbapenemase-Producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Boutal, H.; Vogel, A.; Bernabeu, S.; Devilliers, K.; Creton, E.; Cotellon, G.; Plaisance, M.; Oueslati, S.; Dortet, L.; Jousset, A.; et al. A Multiplex Lateral Flow Immunoassay for the Rapid Identification of NDM-, KPC-, IMP- and VIM-Type and OXA-48-like Carbapenemase-Producing Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 909–915. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Chen, H.-M.; Hii, I.-M.; Hsueh, P.-R. Carbapenemase-Producing Enterobacterales Infections: Recent Advances in Diagnosis and Treatment. Int. J. Antimicrob. Agents 2022, 59, 106528. [Google Scholar] [CrossRef]
- Ambler, R.P. The Structure of Beta-Lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Ma, J.; Song, X.; Li, M.; Yu, Z.; Cheng, W.; Yu, Z.; Zhang, W.; Zhang, Y.; Shen, A.; Sun, H.; et al. Global Spread of Carbapenem-Resistant Enterobacteriaceae: Epidemiological Features, Resistance Mechanisms, Detection and Therapy. Microbiol. Res. 2023, 266, 127249. [Google Scholar] [CrossRef]
- Bonomo, R.A.; Burd, E.M.; Conly, J.; Limbago, B.M.; Poirel, L.; Segre, J.A.; Westblade, L.F. Carbapenemase-Producing Organisms: A Global Scourge. Clin. Infect. Dis. 2018, 66, 1290–1297. [Google Scholar] [CrossRef]
- Potter, R.F.; D’Souza, A.W.; Dantas, G. The Rapid Spread of Carbapenem-Resistant Enterobacteriaceae. Drug Resist. Updates 2016, 29, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef]
- Brolund, A.; Lagerqvist, N.; Byfors, S.; Struelens, M.J.; Monnet, D.L.; Albiger, B.; Kohlenberg, A.; European Antimicrobial Resistance Genes Surveillance Network (EURGen-Net) Capacity Survey Group. Worsening Epidemiological Situation of Carbapenemase-Producing Enterobacteriaceae in Europe, Assessment by National Experts from 37 Countries, July 2018. Eurosurveillance 2019, 24, 1900123. [Google Scholar] [CrossRef]
- Boyd, S.E.; Holmes, A.; Peck, R.; Livermore, D.M.; Hope, W. OXA-48-Like β-Lactamases: Global Epidemiology, Treatment Options, and Development Pipeline. Antimicrob. Agents Chemother. 2022, 66, e0021622. [Google Scholar] [CrossRef]
- Sheu, C.-C.; Chang, Y.-T.; Lin, S.-Y.; Chen, Y.-H.; Hsueh, P.-R. Infections Caused by Carbapenem-Resistant Enterobacteriaceae: An Update on Therapeutic Options. Front. Microbiol. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Koenig, C.; Kuti, J.L. Evolving Resistance Landscape in Gram-Negative Pathogens: An Update on β-Lactam and β-Lactam-Inhibitor Treatment Combinations for Carbapenem-Resistant Organisms. Pharmacotherapy 2024, 44, 658–674. [Google Scholar] [CrossRef]
- Doi, Y. Treatment Options for Carbapenem-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2019, 69, S565–S575. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Ucha, J.C.; Arca-Suárez, J.; Bou, G.; Beceiro, A. New Carbapenemase Inhibitors: Clearing the Way for the β-Lactams. Int. J. Mol. Sci. 2020, 21, 9308. [Google Scholar] [CrossRef]
- Wong, D.; van Duin, D. Novel Beta-Lactamase Inhibitors: Unlocking Their Potential in Therapy. Drugs 2017, 77, 615–628. [Google Scholar] [CrossRef]
- Karaiskos, I.; Galani, I.; Daikos, G.L.; Giamarellou, H. Breaking Through Resistance: A Comparative Review of New Beta-Lactamase Inhibitors (Avibactam, Vaborbactam, Relebactam) Against Multidrug-Resistant Superbugs. Antibiotics 2025, 14, 528. [Google Scholar] [CrossRef]
- Cruz-López, F.; Martínez-Meléndez, A.; Morfin-Otero, R.; Rodriguez-Noriega, E.; Maldonado-Garza, H.J.; Garza-González, E. Efficacy and In Vitro Activity of Novel Antibiotics for Infections With Carbapenem-Resistant Gram-Negative Pathogens. Front. Cell. Infect. Microbiol. 2022, 12, 884365. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, K.; van Duin, D. Treatment for Carbapenem-Resistant Enterobacterales Infections: Recent Advances and Future Directions. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2053–2068. [Google Scholar] [CrossRef]
- Brennan-Krohn, T.; Manetsch, R.; O’Doherty, G.A.; Kirby, J.E. New Strategies and Structural Considerations in Development of Therapeutics for Carbapenem-Resistant Enterobacteriaceae. Transl. Res. J. Lab. Clin. Med. 2020, 220, 14–32. [Google Scholar] [CrossRef]
- Lasko, M.J.; Nicolau, D.P. Carbapenem-Resistant Enterobacterales: Considerations for Treatment in the Era of New Antimicrobials and Evolving Enzymology. Curr. Infect. Dis. Rep. 2020, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Karakonstantis, S.; Rousaki, M.; Kritsotakis, E.I. Cefiderocol: Systematic Review of Mechanisms of Resistance, Heteroresistance and In Vivo Emergence of Resistance. Antibiotics 2022, 11, 723. [Google Scholar] [CrossRef]
- Trecarichi, E.M.; Tumbarello, M. Therapeutic Options for Carbapenem-Resistant Enterobacteriaceae Infections. Virulence 2017, 8, 470–484. [Google Scholar] [CrossRef]
- Bou Zerdan, M.; Al Hassan, S.; Shaker, W.; El Hajjar, R.; Allam, S.; Bou Zerdan, M.; Naji, A.; Zeineddine, N. Carbapenemase Inhibitors: Updates on Developments in 2021. J. Clin. Med. Res. 2022, 14, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U.; Jumde, R.P.; Hennessy, A.; Cohn, J.; Piddock, L.J.V. Global Health Perspectives on Antibacterial Drug Discovery and the Preclinical Pipeline. Nat. Rev. Microbiol. 2025. Online ahead of print. [Google Scholar] [CrossRef]
- Fratoni, A.J.; Berry, A.V.; Liu, X.; Chen, X.; Wu, Y.; Nicolau, D.P.; Abdelraouf, K. Imipenem/Funobactam (Formerly XNW4107) in Vivo Pharmacodynamics against Serine Carbapenemase-Producing Gram-Negative Bacteria: A Novel Modelling Approach for Time-Dependent Killing. J. Antimicrob. Chemother. 2023, 78, 2343–2353. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Castanheira, M.; Lindley, J.; Rubio-Aparicio, D.; Nelson, K.; Tsivkovski, R.; Sun, D.; Totrov, M.; Loutit, J.; Dudley, M. In Vitro Potency of Xeruborbactam in Combination with Multiple β-Lactam Antibiotics in Comparison with Other β-Lactam/β-Lactamase Inhibitor (BLI) Combinations against Carbapenem-Resistant and Extended-Spectrum β-Lactamase-Producing Enterobacterales. Antimicrob. Agents Chemother. 2023, 67, e0044023. [Google Scholar] [CrossRef]
- Durand-Réville, T.F.; Comita-Prevoir, J.; Zhang, J.; Wu, X.; May-Dracka, T.L.; Romero, J.A.C.; Wu, F.; Chen, A.; Shapiro, A.B.; Carter, N.M.; et al. Discovery of an Orally Available Diazabicyclooctane Inhibitor (ETX0282) of Class A, C, and D Serine β-Lactamases. J. Med. Chem. 2020, 63, 12511–12525. [Google Scholar] [CrossRef] [PubMed]
- Outeda-García, M.; Arca-Suárez, J.; Lence, E.; Rodriguez-Coello, A.; Maceiras, R.; Blanco-Martin, T.; Guijarro-Sánchez, P.; Gonzalez-Pinto, L.; Alonso-Garcia, I.; García-Pose, A.; et al. Advancements in the Fight against Globally Distributed OXA-48 Carbapenemase: Evaluating the New Generation of Carbapenemase Inhibitors. Antimicrob. Agents Chemother. 2025, 69, e0161424. [Google Scholar] [CrossRef] [PubMed]
- Tooke, C.L.; Hinchliffe, P.; Krajnc, A.; Mulholland, A.J.; Brem, J.; Schofield, C.J.; Spencer, J. Cyclic Boronates as Versatile Scaffolds for KPC-2 β-Lactamase Inhibition. RSC Med. Chem. 2020, 11, 491–496. [Google Scholar] [CrossRef]
- Everett, M.; Sprynski, N.; Coelho, A.; Castandet, J.; Bayet, M.; Bougnon, J.; Lozano, C.; Davies, D.T.; Leiris, S.; Zalacain, M.; et al. Discovery of a Novel Metallo-β-Lactamase Inhibitor That Potentiates Meropenem Activity against Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e00074-18. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, Y.-H.; Schofield, C.J.; McNally, A.; Zong, Z.; Li, G.-B. Metallo-β-Lactamase-Mediated Antimicrobial Resistance and Progress in Inhibitor Discovery. Trends Microbiol. 2023, 31, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Nahar, L.; Hagiya, H.; Gotoh, K.; Asaduzzaman, M.; Otsuka, F. New Delhi Metallo-Beta-Lactamase Inhibitors: A Systematic Scoping Review. J. Clin. Med. 2024, 13, 4199. [Google Scholar] [CrossRef]
- Chen, C.; Oelschlaeger, P.; Wang, D.; Xu, H.; Wang, Q.; Wang, C.; Zhao, A.; Yang, K.-W. Structure and Mechanism-Guided Design of Dual Serine/Metallo-Carbapenemase Inhibitors. J. Med. Chem. 2022, 65, 5954–5974. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Prester, A.; Perbandt, M.; Galchenkova, M.; Oberthuer, D.; Werner, N.; Henkel, A.; Maracke, J.; Yefanov, O.; Hakanpää, J.; Pompidor, G.; et al. Time-Resolved Crystallography of Boric Acid Binding to the Active Site Serine of the β-Lactamase CTX-M-14 and Subsequent 1,2-Diol Esterification. Commun. Chem. 2024, 7, 152. [Google Scholar] [CrossRef]
- Lu, S.; Hu, L.; Lin, H.; Judge, A.; Rivera, P.; Palaniappan, M.; Sankaran, B.; Wang, J.; Prasad, B.V.V.; Palzkill, T. An Active Site Loop Toggles between Conformations to Control Antibiotic Hydrolysis and Inhibition Potency for CTX-M β-Lactamase Drug-Resistance Enzymes. Nat. Commun. 2022, 13, 6726. [Google Scholar] [CrossRef]
- Krajnc, A.; Gobec, S. Conjugates of Monocyclic β-Lactams and Siderophore Mimetics: A Patent Evaluation (WO2023023393). Expert Opin. Ther. Pat. 2023, 33, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Hyte, M.L.; Arphai, L.J.; Vaughn, C.J.; Durham, S.H. The Role of Bezlotoxumab for the Prevention of Recurrent Clostridioides Difficile Infections: A Review of the Current Literature and Paradigm Shift after 2021. Antibiotics 2022, 11, 1211. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F.; Dedrick, R.M.; Schooley, R.T. Phage Therapy for Antibiotic-Resistant Bacterial Infections. Annu. Rev. Med. 2022, 73, 197–211. [Google Scholar] [CrossRef]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef]
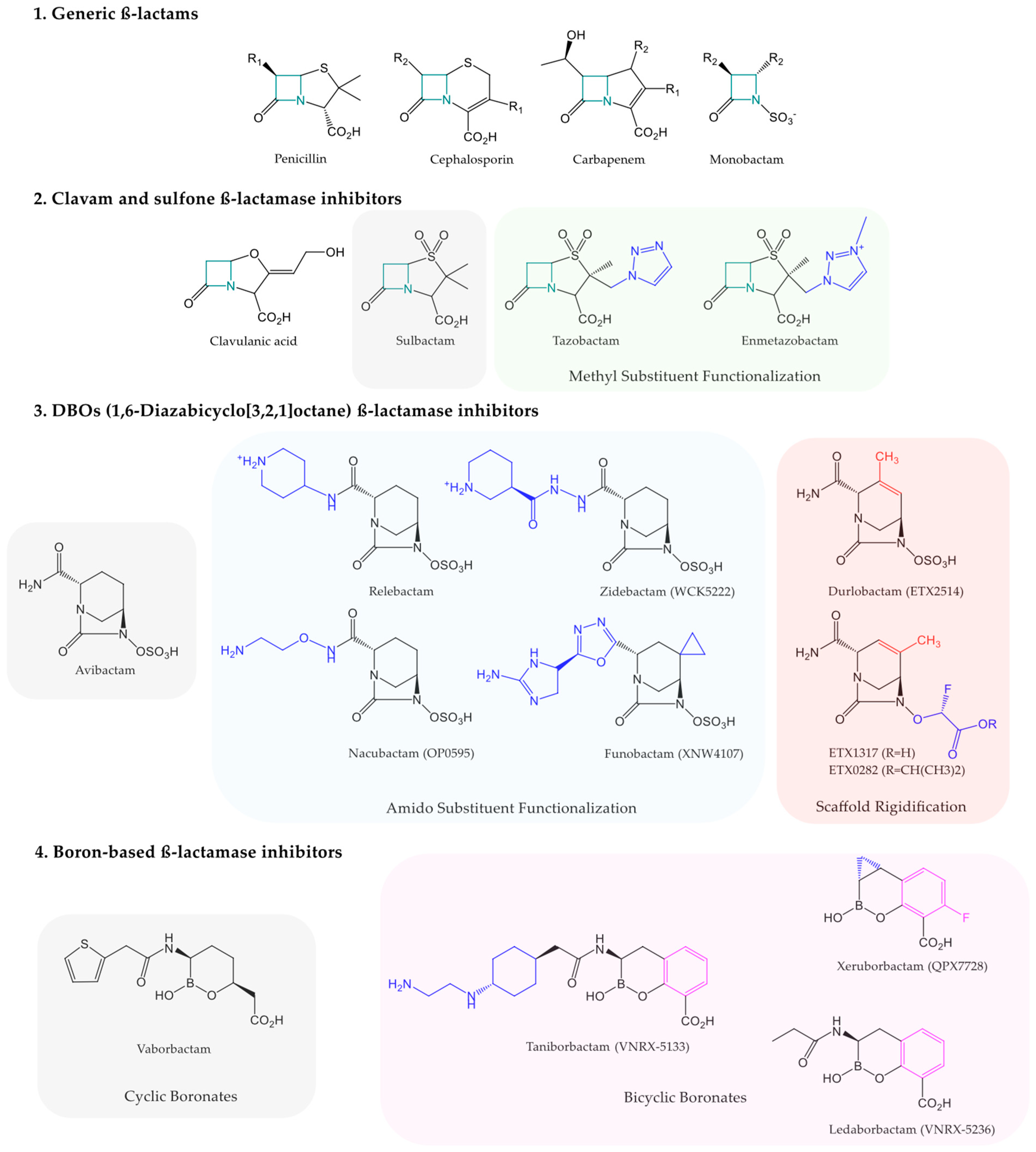

{kind=link}
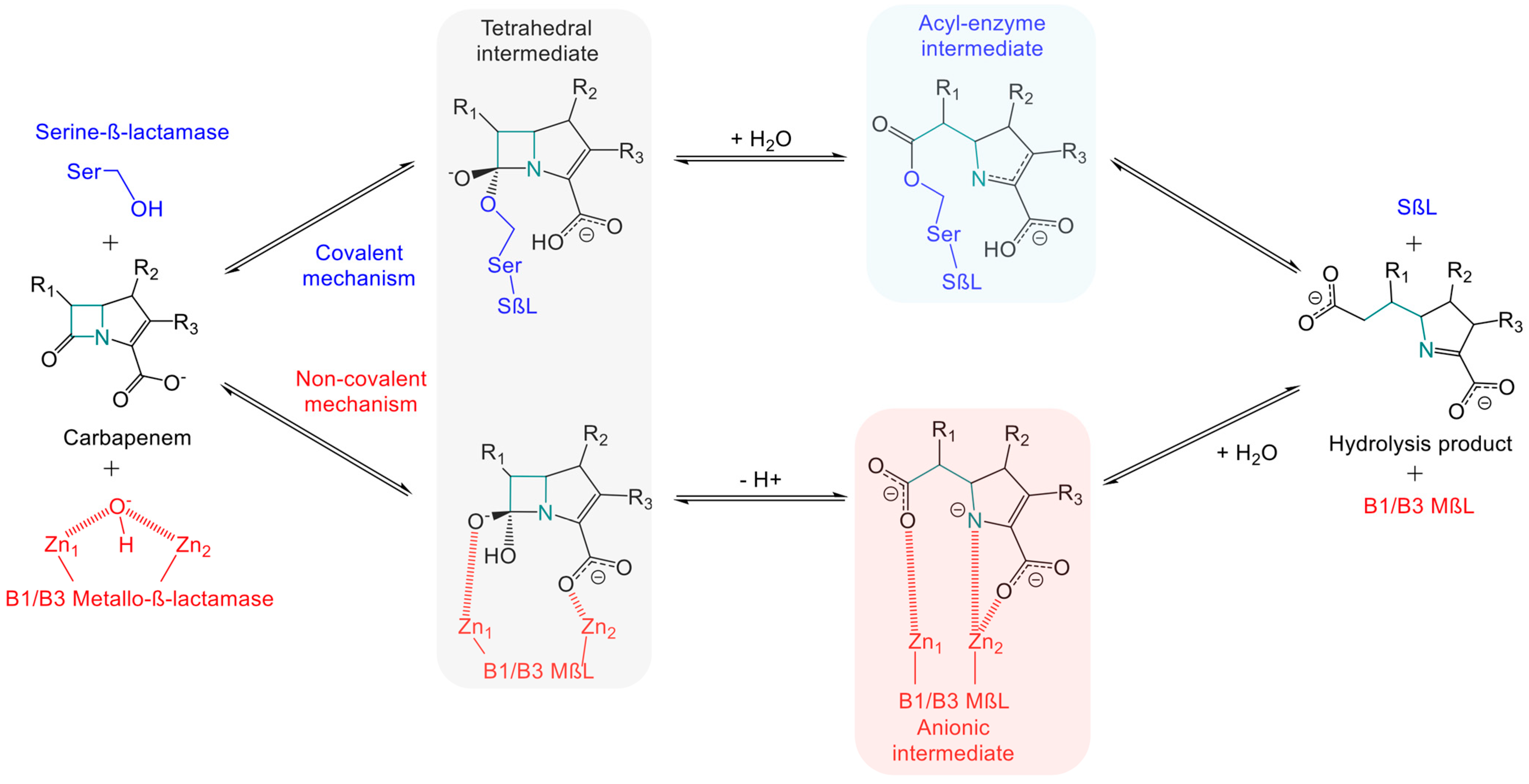
{kind=link}
| Carbapenemases | KPC | MβL (NDM, VIM, IMP) | OXA-48 | ||
|---|---|---|---|---|---|
| Ambler classification | A | B | D | ||
| Active site | Serine | Zn2+ | Serine | ||
| Geographical areas with high prevalence | America | Asia, Australia Middle East | Europe | ||
| β-lactamase inhibitor drugs | |||||
| Clavam | Clavulanic acid | ✕ | ✕ | ✕ | |
| Penicillanic acid sulfone | Tazobactam | ✕ | ✕ | ✕ | |
| Sulbactam | ✕ | ✕ | ✕ | ||
| Enmetazobactam | ≈ | ✕ | ✕ | ||
| Diazabicyclooctanone | Avibactam | ✓ | ✕ | ✓ | |
| Relebactam | ✓ | ✕ | ✕ | ||
| Boronic acid | Vaborbactam | ✓ | ✕ | ✕ | |
| New antibiotic drugs | |||||
| Cephalosporin | Cefiderocol | ✓ | ≈ (NDM) | ✓ (VIM, IMP) | ✓ |
| Tetracycline | Evaracycline | ✓ | ✓ | ✓ | |
| Aminoglycoside | Plazomicin | ✓ | ≈ (NDM, VIM) | ✓ (IMP) | ✓ |
| β-lactamase inhibitor drug candidates | |||||
| Diazabicyclooctanone | Nacubactam (Phase III) | ✓ | ✕ | ≈ | |
| Zidebactam (Phase III) | ✓ | ✕ | ✓ | ||
| Durlobactam (Phase III) | ✓ | ✕ | ✓ | ||
| Funobactam (Phase III) | ✓ | ✕ | ✓ | ||
| ETX0282 (Phase I) | ✓ | ✕ | ✓ | ||
| Boronic acid | Taniborbactam (Phase III) | ✓ | ✓ (NDM, VIM) | ✕ (IMP) | ✓ |
| Xeruborbactam (Phase I) | ✓ | ✓ (NDM, VIM) | ≈ (IMP) | ✓ | |
| Ledaborbactam (Phase I) | ✓ | ✕ | ✓ | ||
| Antibiotic drug candidates | |||||
| Ancremonam LYS228 (Phase II) | ✓ | ✓ | ✓ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leroux, P.; Bornet, C.; Bolla, J.-M.; Cohen, A. Challenges of Carbapenem-Resistant Enterobacteriaceae in the Development of New β-Lactamase Inhibitors and Antibiotics. Antibiotics 2025, 14, 587. https://doi.org/10.3390/antibiotics14060587
Leroux P, Bornet C, Bolla J-M, Cohen A. Challenges of Carbapenem-Resistant Enterobacteriaceae in the Development of New β-Lactamase Inhibitors and Antibiotics. Antibiotics. 2025; 14(6):587. https://doi.org/10.3390/antibiotics14060587
Chicago/Turabian StyleLeroux, Pierre, Charleric Bornet, Jean-Michel Bolla, and Anita Cohen. 2025. "Challenges of Carbapenem-Resistant Enterobacteriaceae in the Development of New β-Lactamase Inhibitors and Antibiotics" Antibiotics 14, no. 6: 587. https://doi.org/10.3390/antibiotics14060587
APA StyleLeroux, P., Bornet, C., Bolla, J.-M., & Cohen, A. (2025). Challenges of Carbapenem-Resistant Enterobacteriaceae in the Development of New β-Lactamase Inhibitors and Antibiotics. Antibiotics, 14(6), 587. https://doi.org/10.3390/antibiotics14060587