
Surviving Colonies of Pseudomonas aeruginosa Isolated In Vivo from Infected, Antibiotic-Treated Galleria mellonella Larvae Acquire an Antibiotic-Tolerant Phenotype


Abstract
1. Introduction
2. Results
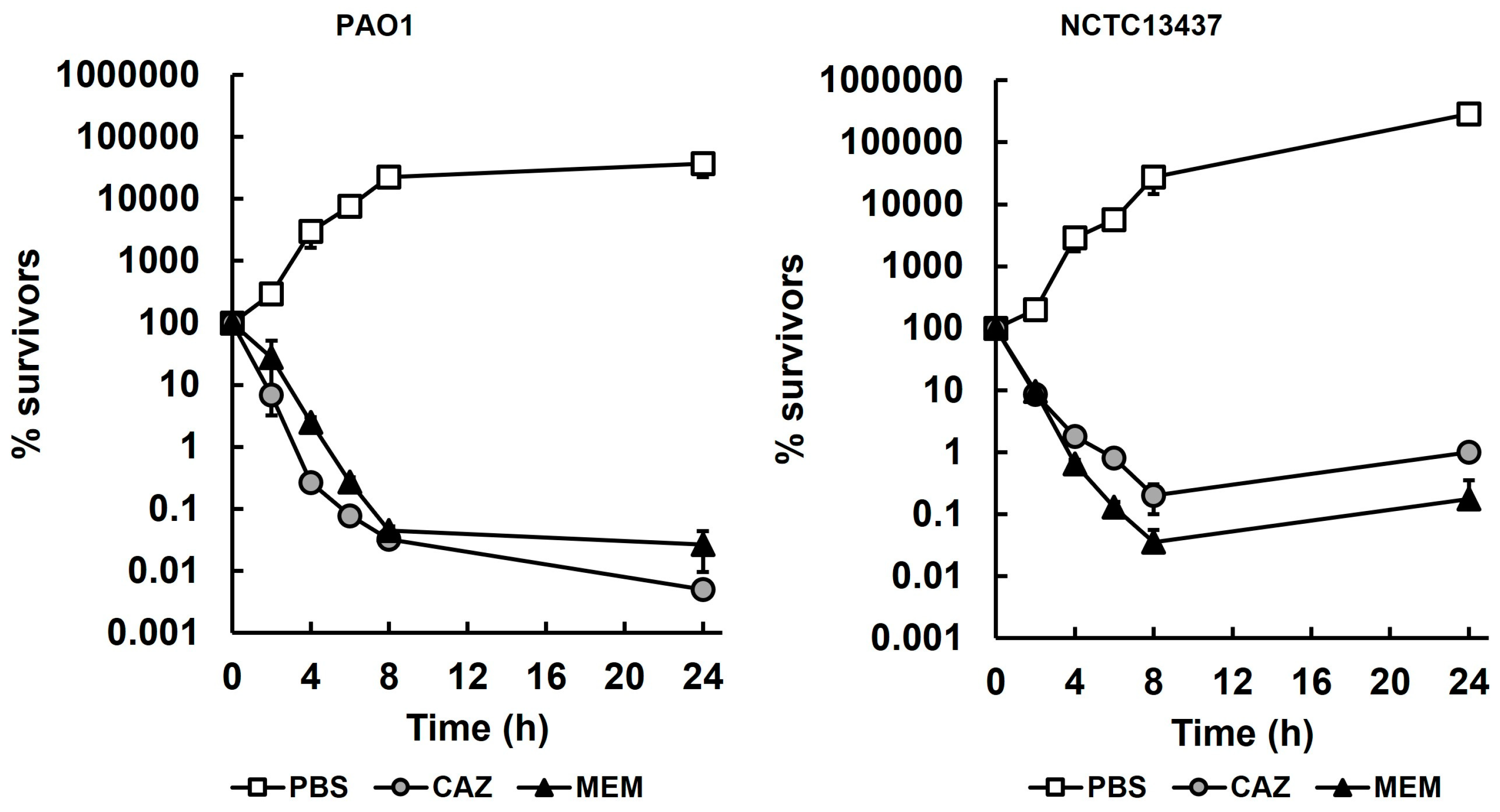
2.1. Susceptibility and Killing of P. aeruginosa Strains by Ceftazidime and Meropenem
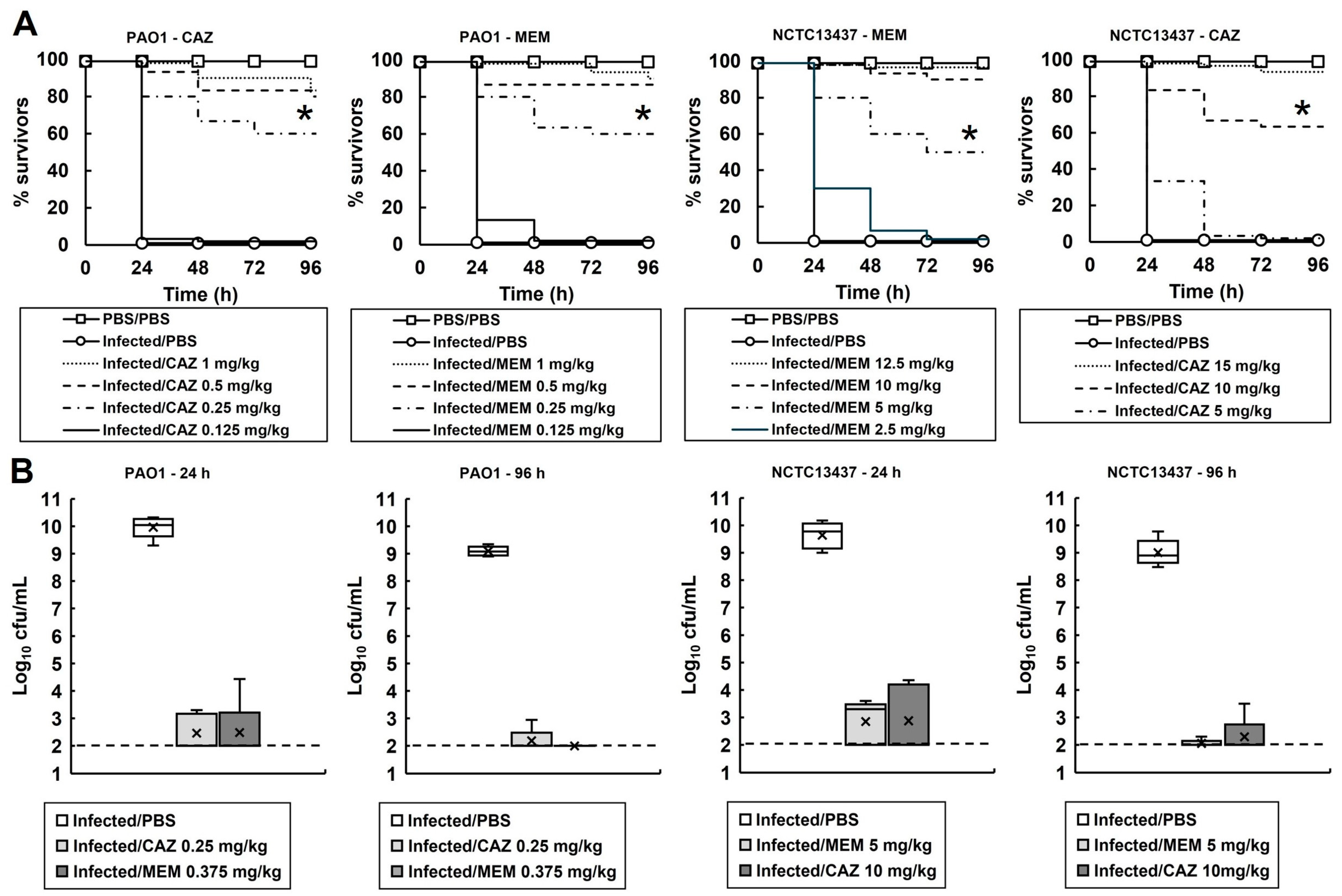
2.2. G. mellonella Larvae Infected with P. aeruginosa and Treated with Meropenem or Ceftazidime Show Enhanced Survival Despite the Persistence of Low Numbers of Infecting Bacteria
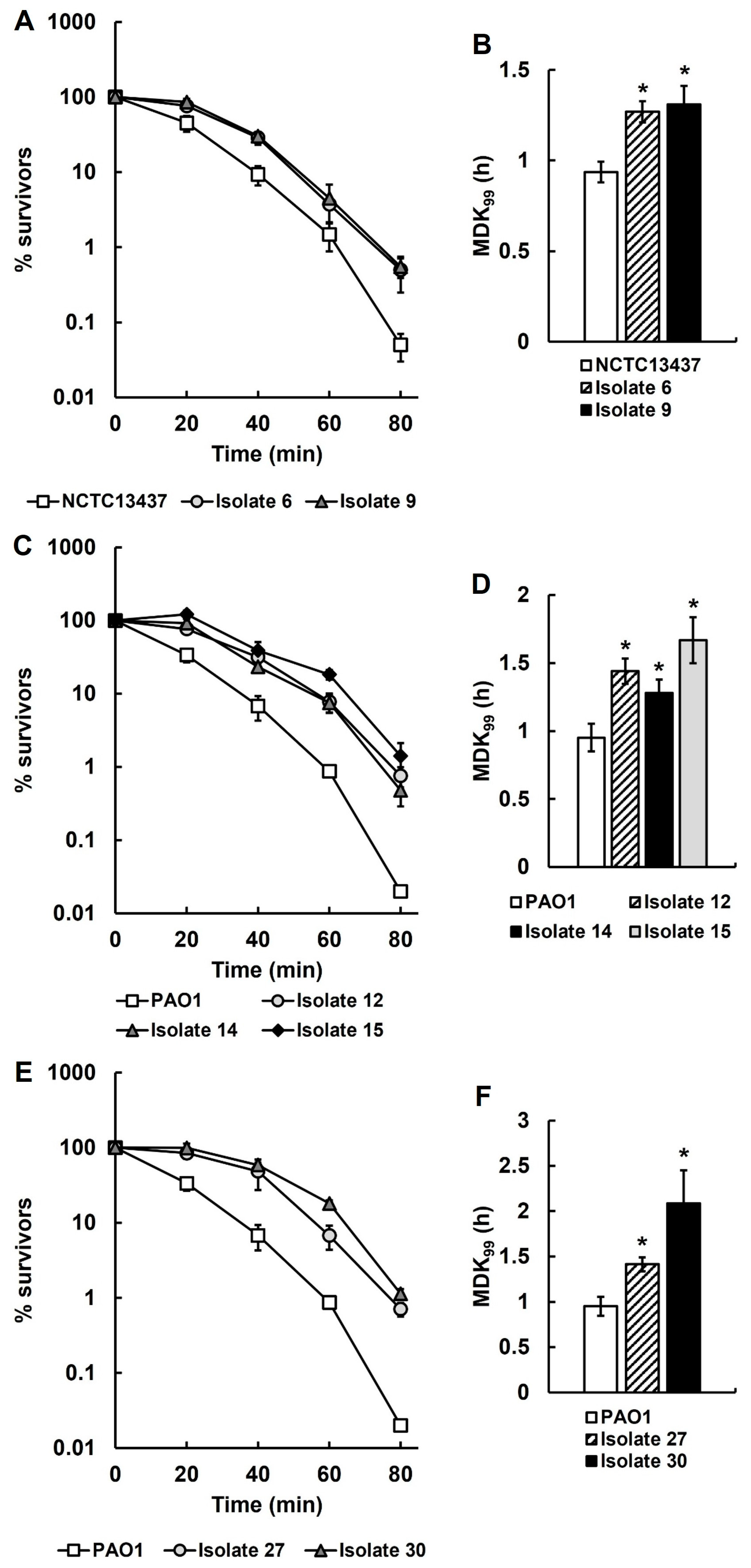
2.3. Cells Derived from P. aeruginosa PAO1 and NCTC13437 and Isolated as Single Colonies from Antibiotic-Treated Larvae Display Phenotypic Differences Compared to Their Parent Strains
2.4. Short-Read Whole Genome Sequencing of P. aeruginosa Isolates Recovered After Antibiotic Exposure In Vivo Detects No Changes When Compared with the Untreated Parental Strains
2.5. G. mellonella Larvae Infected with P. aeruginosa PAO1 and Treated with Three Doses of Ceftazidime Show Enhanced Survival Despite the Persistence of Small Numbers of Infecting Bacteria
2.6. P. aeruginosa PAO1 Isolates Recovered from Infected Larvae Treated with Three Doses of Ceftazidime Have Antibiotic-Tolerant Phenotypes Compared to Their Untreated Parent Strain
2.7. Long-Read Whole Genome Sequencing of P. aeruginosa PAO1 Isolates Recovered After Exposure to Three Doses of Ceftazidime In Vivo Reveals a SNP in the bkdB Gene Causing a Missense Variation
2.8. Mutant P. aeruginosa Strains with a bkdB Gene Knockout Do Not Display an Antibiotic-Tolerant Phenotype Compared to the Parent P. aeruginosa MPAO1
2.9. The Recovered Antibiotic-Tolerant P. aeruginosa Isolates Are Significantly More Thermotolerant than Their Untreated Parent Strains
3. Discussion
4. Materials and Methods
4.1. Bacteria and Growth Media
4.2. Antibiotics and G. mellonella Larvae
4.3. Antibiotic Susceptibility Testing
4.4. In Vitro Time-Kill Assays
4.5. G. mellonella Infection Model
4.6. Burden of P. aeruginosa in G. mellonella Larvae and Isolation of Surviving Cells
4.7. Measurement of the MIC, Growth Rate, MDK99 and Recovery, Virulence and Thermotolerance of Antibiotic-Treated P. aeruginosa Isolates
4.8. Whole Genome Sequencing and Bioinformatic Analysis of P. aeruginosa Antibiotic-Treated Isolates
4.9. Analysis of P. aeruginosa MPAO1 with bdkB Knockout
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynolds, D.; Kollef, M. The epidemiology and pathogenesis and treatment of Pseudomonas aeruginosa infections: An update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef]
- Weiner-Lastinger, L.M.; Abner, S.; Edwards, J.R.; Kallen, A.J.; Karlsson, M.; Magill, S.S.; Pollock, D.; See, I.; Soe, M.M.; Walters, M.S.; et al. Antimicrobial-resistant pathogens associated with adult healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network, 2015–2017. Infect. Control. Hosp. Epidemiol. 2020, 41, 1–18. [Google Scholar] [CrossRef]
- Micek, S.T.; Kollef, M.H.; Torres, A.; Chen, C.; Rello, J.; Chastre, J.; Antonelli, M.; Welte, T.; Clair, B.; Ostermann, H.; et al. Pseudomonas aeruginosa nosocomial pneumonia: Impact of pneumonia classification. Infect. Control. Hosp. Epidemiol. 2015, 36, 1190–1197. [Google Scholar] [CrossRef]
- Scotet, V.; L’Hostis, C.; Férec, C. The changing epidemiology of cystic fibrosis: Incidence, survival and impact of the CFTR gene discovery. Genes 2020, 11, 589. [Google Scholar] [CrossRef]
- Ratjen, F.; Munck, A.; Kho, P.; Angyalosi, G.; ELITE Study Group. Treatment of early Pseudomonas aeruginosa infection in patients with cystic fibrosis: The ELITE trial. Thorax 2010, 65, 286–291. [Google Scholar] [CrossRef]
- Deventer, A.T.; Stevens, C.E.; Stewart, A.; Hobbs, J.K. Antibiotic tolerance among clinical isolates: Mechanisms, detection, prevalence, and significance. Clin. Microbiol. Rev. 2024, 37, e0010624. [Google Scholar] [CrossRef]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef]
- Gollan, B.; Grabe, G.; Michaux, C.; Helaine, S. Bacterial persisters and infection: Past, present, and progressing. Annu. Rev. Microbiol. 2019, 73, 359–385. [Google Scholar] [CrossRef] [PubMed]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.; Michiels, J.; Wenseleers, T.; Windels, E.M.; Vanden Boer, P.; Kestemont, D.; De Meester, L.; Verstrepen, K.J.; Verstraeten, N.; Fauvart, M.; et al. Frequency of antibiotic application drives rapid evolutionary adaptation of Escherichia coli persistence. Nat. Microbiol. 2016, 1, 16020. [Google Scholar] [CrossRef]
- Michiels, J.E.; Van den Bergh, B.; Verstraeten, N.; Fauvart, M.; Michiels, J. In vitro evolution of high antibiotic tolerance by periodic aminoglycoside challenge in the ESKAPE pathogens. Antimicrob. Agents Chemother. 2016, 60, 4630–4637. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.R.; Burns, J.L.; Lory, S.; Lewis, K. Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J. Bacteriol. 2010, 192, 6191–6199. [Google Scholar] [CrossRef]
- Claudi, B.; Spröte, P.; Chirkova, A.; Personnic, N.; Zankl, J.; Schürmann, N.; Schmidt, A.; Bumann, D. Phenotypic variation of salmonella in host tissues delays eradication by antimicrobial chemotherapy. Cell 2014, 158, 722–733. [Google Scholar] [CrossRef]
- Bartell, J.A.; Cameron, D.R.; Mojsoska, B.; Haagensen, J.A.J.; Pressler, T.; Sommer, L.M.; Lewis, K.; Molin, S.; Johansen, H.K. Bacterial persisters in long-term infection: Emergence and fitness in a complex host environment. PLoS Pathog. 2020, 16, e1009112. [Google Scholar] [CrossRef]
- Santi, I.; Manfredi, P.; Maffei, E.; Egli, A.; Jenal, U. Evolution of antibiotic tolerance shapes resistance development in chronic Pseudomonas aeruginosa infections. mBio 2021, 12, 10–1128. [Google Scholar] [CrossRef]
- Ghoul, M.; Andersen, S.B.; Marvig, R.L.; Johansen, H.K.; Jelsbak, L.; Molin, S.; Perron, G.; Griffin, A.S. Long-term evolution of antibiotic tolerance in Pseudomonas aeruginosa lung infections. Evol. Lett. 2023, 7, 389–400. [Google Scholar] [CrossRef]
- Schumacher, M.A.; Balani, P.; Min, J.; Chinnam, N.B.; Hansen, S.; Vulić, M.; Lewis, K.; Brennan, R.G. HipBA–promoter structures reveal the basis of heritable multidrug tolerance. Nature 2015, 524, 59–64. [Google Scholar] [CrossRef]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef]
- Radzikowski, J.L.; Vedelaar, S.; Siegel, D.; Ortega, Á.D.; Schmidt, A.; Heinemann, M. Bacterial persistence is an active σS stress response to metabolic flux limitation. Mol. Syst. Biol. 2016, 12, 882. [Google Scholar] [CrossRef] [PubMed]
- Baharoglu, Z.; Mazel, D. SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol. Rev. 2014, 38, 1126–1145. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Gandt, A.B.; Rowe, S.E.; Deisinger, J.P.; Conlon, B.P.; Lewis, K. ATP-dependent persister formation in Escherichia coli. mBio 2017, 8, e02267-16. [Google Scholar] [CrossRef] [PubMed]
- Rowe, S.E.; Wagner, N.J.; Li, L.; Beam, J.E.; Wilkinson, A.D.; Radlinski, L.C.; Zhang, Q.; Miao, E.A.; Conlon, B.P. Reactive oxygen species induce antibiotic tolerance during systemic Staphylococcus aureus infection. Nat. Microbiol. 2020, 5, 282–290. [Google Scholar] [CrossRef]
- Ledger, E.V.K.; Mesnage, S.; Edwards, A.M. Human serum triggers antibiotic tolerance in Staphylococcus aureus. Nat. Commun. 2022, 13, 2041. [Google Scholar] [CrossRef]
- Siriyong, T.; Voravuthikunchai, S.P.; Coote, P.J. Steroidal alkaloids and conessine from the medicinal plant Holarrhena antidysenterica restore antibiotic efficacy in a Galleria mellonella model of multidrug-resistant Pseudomonas aeruginosa infection. BMC Complement. Altern. Med. 2018, 18, 285. [Google Scholar] [CrossRef]
- Mackay, B.; Parcell, B.J.; Shirran, S.L.; Coote, P.J. Carbapenem-only combination therapy against multi-drug resistant Pseudomonas aeruginosa: Assessment of in vitro and in vivo efficacy and mode of action. Antibiotics 2022, 11, 1467. [Google Scholar] [CrossRef]
- Fletcher, M.; McCormack, A.; Parcell, B.J.; Coote, P.J. Combination therapy with ciprofloxacin and pentamidine against multidrug-resistant Pseudomonas aeruginosa: Assessment of in vitro and in vivo efficacy and the role of resistance–nodulation–division (RND) efflux pumps. Antibiotics 2023, 12, 1236. [Google Scholar] [CrossRef]
- Svedholm, E.; Bruce, B.; Parcell, B.J.; Coote, P.J. Repurposing mitomycin C in combination with pentamidine or gentamicin to treat infections with multi-drug-resistant (MDR) Pseudomonas aeruginosa. Antibiotics 2024, 13, 177. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 14.0, 2024. Available online: http://www.eucast.org (accessed on 5 December 2024).
- Zou, J.; Kou, S.-H.; Xie, R.; VanNieuwenhze, M.S.; Qu, J.; Peng, B.; Zheng, J. Non-walled spherical Acinetobacter baumannii is an important type of persister upon β-lactam antibiotic treatment. Emerg. Microbes Infect. 2020, 9, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B. Bugs on drugs: A Drosophila melanogaster gut model to study in vivo antibiotic tolerance of E. coli. Microorganisms 2022, 10, 119. [Google Scholar] [CrossRef]
- Wolfson, J.S.; Hooper, D.C.; McHugh, G.L.; Bozza, M.A.; Swartz, M.N. Mutants of Escherichia coli K-12 exhibiting reduced killing by both quinolone and beta-lactam antimicrobial agents. Antimicrob. Agents Chemother. 1990, 34, 1938–1943. [Google Scholar] [CrossRef]
- Lopatkin, A.J.; Stokes, J.M.; Zheng, E.J.; Yang, J.H.; Takahashi, M.K.; You, L.; Collins, J.J. Bacterial metabolic state more accurately predicts antibiotic lethality than growth rate. Nat. Microbiol. 2019, 4, 2109–2117. [Google Scholar] [CrossRef] [PubMed]
- Dawan, J.; Ahn, J. Bacterial stress responses as potential targets in overcoming antibiotic resistance. Microorganisms 2022, 10, 1385. [Google Scholar] [CrossRef] [PubMed]
- Burns, G.; Brown, T.; Hatter, K.; Sokatch, J.R. Comparison of the amino acid sequences of the transacylase components of branched chain oxoacid dehydrogenase of Pseudomonas putida, and the pyruvate and 2-oxoglutarate dehydrogenases of Escherichia coli. Eur. J. Biochem. 1988, 176, 165–169. [Google Scholar] [CrossRef]
- Ward, D.E.; Ross, R.P.; van der Weijden, C.C.; Snoep, J.L.; Claiborne, A. Catabolism of branched-chain α-keto acids in Enterococcus faecalis: The bkd gene cluster, enzymes, and metabolic route. J. Bacteriol. 1999, 181, 5433–5442. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Wynn, R.M.; Chuang, J.L.; Brautigam, C.A.; Custorio, M.; Chuang, D.T. A synchronized substrate-gating mechanism revealed by cubic-core structure of the bovine branched-chain α-ketoacid dehydrogenase complex. EMBO J. 2006, 25, 5983–5994. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, H.; Chen, X.; Liu, L. Engineering microbial membranes to increase stress tolerance of industrial strains. Metab. Eng. 2019, 53, 24–34. [Google Scholar] [CrossRef]
- Sinensky, M. Homeoviscous adaptation—A homeostatic process that regulates the viscosity of membrane lipids in Escherichia coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–525. [Google Scholar] [CrossRef]
- Van de Vossenberg, J.L.C.M.; Driessen, A.J.M.; da Costa, M.S.; Konings, W.M. Homeostasis of the membrane proton permeability in Bacillus subtilis grown at different temperatures. Biochim. Biophys. Acta-Biomemb 1999, 1419, 97–104. [Google Scholar] [CrossRef]
- Silvius, J.R.; Mak, N.; McElhaney, R.N. Lipid and protein composition and thermotropic lipid phase transitions in fatty acid-homogeneous membranes of Acholeplasma laidlawii. Biochim. Biophys. Acta-Biomemb 1980, 597, 199–215. [Google Scholar] [CrossRef]
- Marinus, M.G.; Casadesus, J. Roles of DNA adenine methylation in host–pathogen interactions: Mismatch repair, transcriptional regulation, and more. FEMS Microbiol. Rev. 2009, 33, 488–503. [Google Scholar] [CrossRef]
- Adhikari, S.; Curtis, P.D. DNA methyltransferases and epigenetic regulation in bacteria. FEMS Microbiol. Rev. 2016, 40, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Rolando, M.; Di Silvestre, C.; Gomez-Valero, L.; Buchrieser, C. Bacterial methyltransferases: From targeting bacterial genomes to host epigenetics. Microlife 2022, 3, uqac014. [Google Scholar] [CrossRef] [PubMed]
- Jen, F.E.C.; Seib, K.L.; Jennings, M.P. Phasevarions mediate epigenetic regulation of antimicrobial susceptibility in Neisseria meningitidis. Antimicrob. Agents Chemother. 2014, 58, 4219–4221. [Google Scholar] [CrossRef]
- Braaten, B.A.; Nou, X.; Kaltenbach, L.S.; Low, D.A. Methylation patterns in pap regulatory DNA control pyelonephritis-associated pili phase variation in E. coli. Cell 1994, 76, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Casadesús, J.; Low, D. Epigenetic gene regulation in the bacterial world. Microbiol. Mol. Biol. Rev. 2006, 70, 830–856. [Google Scholar] [CrossRef]
- Ghosh, D.; Veeraraghavan, B.; Elangovan, R.; Vivekanandan, P. Antibiotic resistance and epigenetics: More to it than meets the eye. Antimicrob. Agents Chemother. 2020, 64, e02225–19. [Google Scholar] [CrossRef]
- Riber, L.; Hansen, L.H. Epigenetic memories: The hidden drivers of bacterial persistence? Trends Microbiol. 2021, 29, 190–194. [Google Scholar] [CrossRef]
- Woodford, N.; Zhang, J.; Kaufmann, M.E.; Yarde, S.; Tomas, M.M.; Faris, C.; Vardhan, M.S.; Dawson, S.; Cotterill, S.L.; Livermore, D.M. Detection of Pseudomonas aeruginosa isolates producing VEB-type extended-spectrum β-lactamases in the United Kingdom. J. Antimicrob. Chemother. 2008, 62, 1265–1268. [Google Scholar] [CrossRef]
- Hill, L.; Veli, N.; Coote, P.J. Evaluation of Galleria mellonella larvae for measuring the efficacy and pharmacokinetics of antibiotic therapies against Pseudomonas aeruginosa infection. Int. J. Antimicrob. Agents 2014, 43, 254–261. [Google Scholar] [CrossRef]
- Krezdorn, J.; Adams, S.; Coote, P.J. A Galleria mellonella infection model reveals double and triple antibiotic combination therapies with enhanced efficacy versus a multidrug-resistant strain of Pseudomonas aeruginosa. J. Med. Microbiol. 2014, 63, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Adamson, D.H.; Krikstopaityte, V.; Coote, P.J. Enhanced efficacy of putative efflux pump inhibitor/antibiotic combination treatments versus MDR strains of Pseudomonas aeruginosa in a Galleria mellonella in vivo infection model. J. Antimicrob. Chemother. 2015, 70, 2271–2278. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. Survival probabilities (the Kaplan-Meier method). Brit Med. J. 1998, 317, 1572. [Google Scholar] [CrossRef]
- Bland, J.M. The logrank test. Brit Med. J. 2004, 328, 1073. [Google Scholar] [CrossRef]
- Wick, R.; Volkening, J. Porechop: Adapter Trimmer for Oxford Nanopore Reads (0.2.4). GitHub, 2018. Available online: https://github.com/rrwick/Porechop (accessed on 11 May 2025).
- Wick, R. Filtlong: Quality Filtering Tool for Long Reads (v0.2.1). GitHub, 2021. Available online: https://github.com/rrwick/Filtlong (accessed on 11 May 2025).
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Hunt, M.; Silva, N.D.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesman, J. Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Snippy: Rapid Haploid Variant Calling and Core SNP Phylogeny. 2020. Available online: https://github.com/tseemann/snippy (accessed on 11 May 2025).
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Smolka, M.; Paulin, L.F.; Grochowski, C.M.; Horner, D.W.; Mahmoud, M.; Behera, S.; Kalef-Ezra, E.; Gandhi, M.; Hong, K.; Pehlivan, D.; et al. Detection of mosaic and population-level structural variants with Sniffles2. Nat. Biotechnol. 2024, 42, 1571–1580. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. aeruginosa Strain | Antibiotic Treatment | MDK99 (h) ± SEM |
|---|---|---|
| PAO1 | MEM (2 mg/L) | 4.7 ± 0.3 |
| PAO1 | CAZ (4 mg/L) | 3.8 ± 0.1 |
| NCTC13437 | MEM (128 mg/L) | 4.0 ± 0.1 |
| NCTC13437 | CAZ (2500 mg/L) | 5.4 ± 0.2 |
| P. aeruginosa PAO1 and In Vivo Antibiotic Treatment | Growth Rate (ΔOD600/h) | Reduction in Growth Rate (%) | MDK99 (h) | Increase in MDK99 (%) | Recovery at 24 h (Log10 cfu/mL) | Change in MIC | Change in Virulence |
|---|---|---|---|---|---|---|---|
| Untreated parent control | 0.327 | 0 | 3.8 | 0 | 0 | 0 | 0 |
| Isolate 26 CAZ—0.25 mg/kg | 0.261 | 20.2 | 5.1 | 34 | No (2.0) | No | No |
| Isolate 27 CAZ—0.25 mg/kg | 0.259 | 20.8 | 5.3 | 40 | No (2.3) | No | No |
| Isolate 28 CAZ—0.25 mg/kg | 0.249 | 23.8 | 5.4 | 42 | No (3.0) | No | No |
| Isolate 29 CAZ—0.25 mg/kg | 0.254 | 22.3 | 5.3 | 40 | No (2.8) | No | No |
| Isolate 30 CAZ—0.25 mg/kg | 0.240 | 26.6 | 6.0 | 58 | No (3.3) | No | No |
| Recovered Isolate | NCBI Locus Tag | Position of Mutation | Type of Mutation | Parental Genotype | Isolate Genotype | % of Isolate Population Carrying Mutation | Gene | Biological Function |
|---|---|---|---|---|---|---|---|---|
| Isolate 27 | PA2249 | 3038686 | snp | C | T | 70 | bdkB; lipoamide acyltransferase component of branched-chain alpha-keto acid dehydrogenase complex | Catalyses the conversion of alpha keto acids into acyl-CoA and CO2 |
| Isolate 30 | PA2249 | 3038686 | snp | C | T | 51 | As above | As above |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCormack, A.; Hobbs, J.K.; Johnston, P.R.; Coote, P.J. Surviving Colonies of Pseudomonas aeruginosa Isolated In Vivo from Infected, Antibiotic-Treated Galleria mellonella Larvae Acquire an Antibiotic-Tolerant Phenotype. Antibiotics 2025, 14, 507. https://doi.org/10.3390/antibiotics14050507
McCormack A, Hobbs JK, Johnston PR, Coote PJ. Surviving Colonies of Pseudomonas aeruginosa Isolated In Vivo from Infected, Antibiotic-Treated Galleria mellonella Larvae Acquire an Antibiotic-Tolerant Phenotype. Antibiotics. 2025; 14(5):507. https://doi.org/10.3390/antibiotics14050507
Chicago/Turabian StyleMcCormack, Alex, Joanne K. Hobbs, Paul R. Johnston, and Peter J. Coote. 2025. "Surviving Colonies of Pseudomonas aeruginosa Isolated In Vivo from Infected, Antibiotic-Treated Galleria mellonella Larvae Acquire an Antibiotic-Tolerant Phenotype" Antibiotics 14, no. 5: 507. https://doi.org/10.3390/antibiotics14050507
APA StyleMcCormack, A., Hobbs, J. K., Johnston, P. R., & Coote, P. J. (2025). Surviving Colonies of Pseudomonas aeruginosa Isolated In Vivo from Infected, Antibiotic-Treated Galleria mellonella Larvae Acquire an Antibiotic-Tolerant Phenotype. Antibiotics, 14(5), 507. https://doi.org/10.3390/antibiotics14050507