

Antimicrobial Susceptibility Profiles of Staphylococcus aureus and Streptococcus spp. Isolates from Clinical Cases of Waterfowl in Hungary Between 2022 and 2023



Abstract
1. Introduction
2. Results
2.1. Origin of the Strains
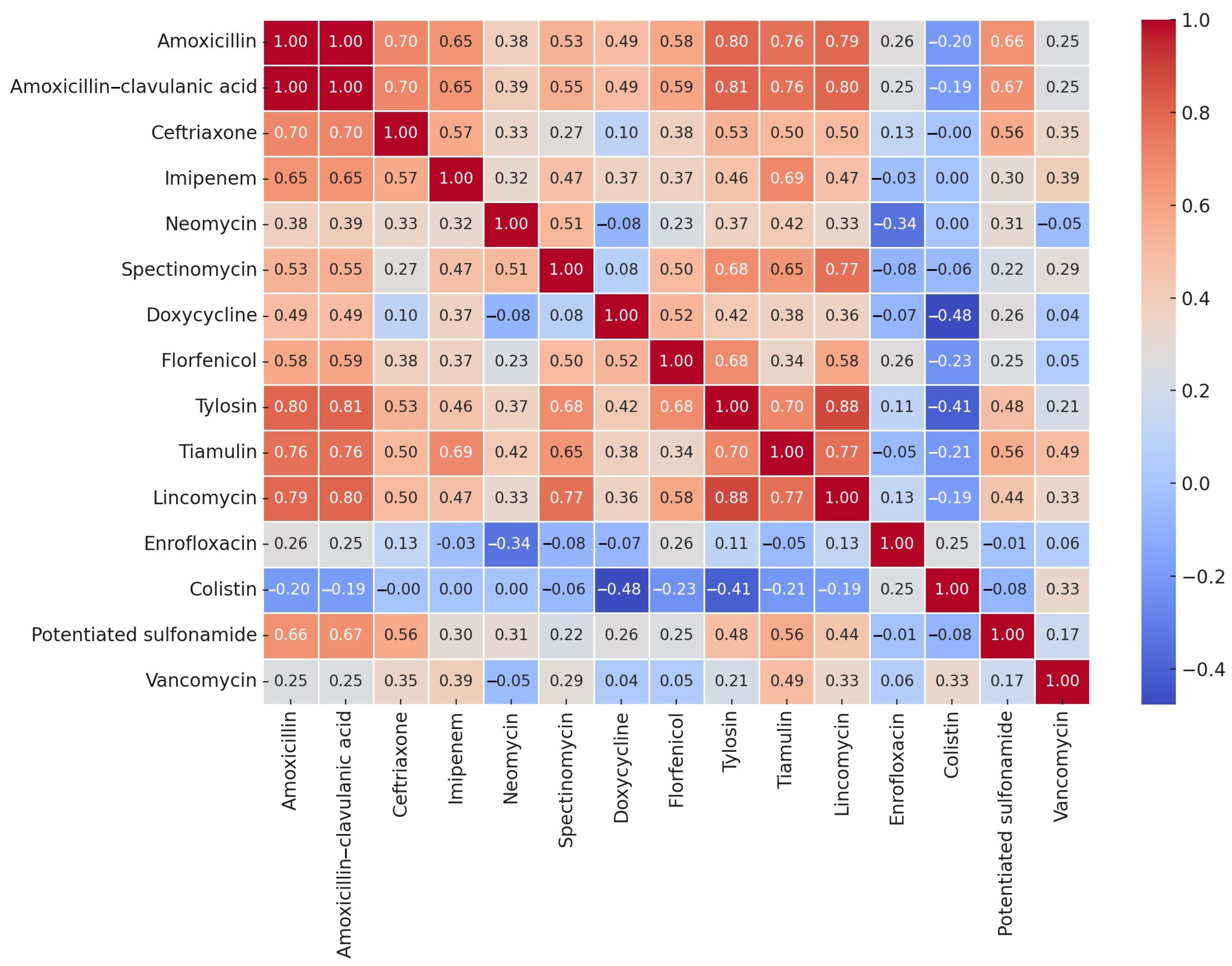
2.2. Minimum Inhibitory Concentration Values
3. Discussion
4. Materials and Methods
4.1. Origin of Strains
4.2. Preparation of Antibiotic Stock Solutions
4.3. Determination of Minimum Inhibitory Concentration
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMR | Antimicrobial resistance |
| CLSI | Clinical and Laboratory Standards Institute |
| ECOFF | Epidemiological cut-off values |
| MHB | Mueller–Hinton broth |
| MIC | Minimum inhibitory concentration |
References
- Jamali, H.; Radmehr, B.; Ismail, S. Prevalence and Antimicrobial Resistance of Listeria, Salmonella, and Yersinia Species Isolates in Ducks and Geese. Poult. Sci. 2014, 93, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Reddy, B.; Singh, V.K.; Singh, A.K.; Kesari, K.K.; Tripathi, P.; Kumar, P.; Tripathi, V.; Simal-Gandara, J. Potential Environmental and Human Health Risks Caused by Antibiotic-Resistant Bacteria (ARB), Antibiotic Resistance Genes (ARGs) and Emerging Contaminants (ECs) from Municipal Solid Waste (MSW) Landfill. Antibiotics 2021, 10, 374. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Chaudhury, R.R. Antibiotic Resistance in India: Drivers and Opportunities for Action. PLoS Med. 2016, 13, e1001974. [Google Scholar] [CrossRef]
- Witte, W. Selective Pressure by Antibiotic Use in Livestock. Int. J. Antimicrob. Agents 2000, 16 (Suppl. 1), S19–S24. [Google Scholar] [CrossRef]
- Hetényi, N.; Bersényi, A.; Hullár, I. Physiological Effects of Medium-Chain Fatty Acids and Triglycerides, and Their Potential Use in Poultry and Swine Nutrition: A Literature Review. Magy. Állatorvosok Lapja 2024, 146, 651–659. [Google Scholar] [CrossRef]
- Jerzsele, Á.; Somogyi, Z.; Szalai, M.; Kovács, D. Effects of Fermented Wheat Germ Extract on Artificial Salmonella Typhimurium Infection in Broiler Chickens. Magy. Állatorvosok Lapja 2020, 142, 77–85. [Google Scholar]
- Kerek, Á.; Csanády, P.; Jerzsele, Á. Antibacterial Efficiency of Propolis—Part 1. Magy. Állatorvosok Lapja 2022, 144, 285–298. [Google Scholar]
- Kerek, Á.; Csanády, P.; Jerzsele, Á. Antiprotozoal and Antifungal Efficiency of Propolis—Part 2. Magy. Állatorvosok Lapja 2022, 144, 691–704. [Google Scholar]
- Kovács, L.; Nagy, D.; Könyves, L.; Jerzsele, Á.; Kerek, Á. Antimicrobial Properties of Essential Oils—Animal Health Aspects. Magy. Állatorvosok Lapja 2023, 145, 497–510. [Google Scholar] [CrossRef]
- Olasz, Á.; Jerzsele, Á.; Balta, L.; Dobra, P.F.; Kerek, Á. In Vivo Efficacy of Different Extracts of Propolis in Broiler Salmonellosis. Magy. Állatorvosok Lapja 2023, 145, 461–475. [Google Scholar] [CrossRef]
- Petrilla, J.; Mátis, G.; Molnár, A.; Jerzsele, Á.; Pál, L.; Gálfi, P.; Neogrády, Z.; Dublecz, K. In Vitro Investigation of the Antibacterial Efficacy of Butyrate on Various Campylobacter jejuni Strains. MÁL 2021, 143, 57–64. [Google Scholar]
- Acikgoz, Z.; Yucel, B.; Altan, O. The Effects of Propolis Supplementation on Broiler Performance and Feed Digestibility. Arch. Geflugelkd. 2005, 69, 117–122. [Google Scholar]
- Sebők, C.; Márton, R.A.; Meckei, M.; Neogrády, Z.; Mátis, G. Antimicrobial Peptides as New Tools to Combat Infectious Diseases. Magy. Állatorvosok Lapja 2024, 146, 181–191. [Google Scholar] [CrossRef]
- Kovács, L.; Hejel, P.; Farkas, M.; László, L.; Könyves, L. Study Report on the Effect of a Litter Treatment Product Containing Bacillus licheniformis and Zeolite in Male Fattening Turkey Flock. Magy. Állatorvosok Lapja 2024, 146, 291–305. [Google Scholar] [CrossRef]
- Such, N.; Molnár, A.; Pál, L.; Farkas, V.; Menyhárt, L.; Husvéth, F.; Dublecz, K. The Effect of Pre- and Probiotic Treatment on the Gumboro-Titer Values of Broilers. Magy. Állatorvosok Lapja 2021, 143, 119–127. [Google Scholar]
- Abd El-Ghany, W.A. Paraprobiotics and Postbiotics: Contemporary and Promising Natural Antibiotics Alternatives and Their Applications in the Poultry Field. Open Vet. J. 2020, 10, 323–330. [Google Scholar] [CrossRef]
- Essősy, M.; Fodor, I.; Ihnáth, Z.; Karancsi, Z.; Kovács, D.; Szalai, K.V.; Szentmiklósi, D.; Jerzsele, Á. The Possibilities of Antibiotic-Free Broiler-Hen Fattening, with Special Reference to the Use of Pre- and Probiotics. Magy. Állatorvosok Lapja 2020, 142, 397–407. [Google Scholar]
- Kovács, D.; Palkovicsné Pézsa, N.; Farkas, O.; Jerzsele, Á. Usage of Antibiotic Alternatives in Pig Farming: Literature Review. Magy. Állatorvosok Lapja 2021, 143, 281–282. [Google Scholar]
- Bager, F.; Madsen, M.; Christensen, J.; Aarestrup, F.M. Avoparcin Used as a Growth Promoter Is Associated with the Occurrence of Vancomycin-Resistant Enterococcus faecium on Danish Poultry and Pig Farms. Prev. Vet. Med. 1997, 31, 95–112. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, S.; Rehman, M.U.; Yang, H.; Yang, Z.; Wang, M.; Jia, R.; Chen, S.; Liu, M.; Zhu, D.; et al. Distribution and Association of Antimicrobial Resistance and Virulence Traits in Escherichia coli Isolates from Healthy Waterfowls in Hainan, China. Ecotoxicol. Environ. Saf. 2021, 220, 112317. [Google Scholar] [CrossRef]
- Albarella, U. Alternate Fortunes? The Role of Domestic Ducks and Geese from Roman to Medieval Times in Britain. Doc. Archaeobiologiae 2005, III, 249–258. [Google Scholar]
- Farkas, M.; Könyves, L.; Csorba, S.; Farkas, Z.; Józwiák, Á.; Süth, M.; Kovács, L. Biosecurity Situation of Large-Scale Poultry Farms in Hungary According to the Databases of National Food Chain Safety Office Centre for Disease Control and Biosecurity Audit System of Poultry Product Board of Hungary in the Period of 2021–2022. Magy. Állatorvosok Lapja 2024, 146, 723–742. [Google Scholar] [CrossRef]
- Adebowale, O.; Makanjuola, M.; Bankole, N.; Olasoju, M.; Alamu, A.; Kperegbeyi, E.; Oladejo, O.; Fasanmi, O.; Adeyemo, O.; Fasina, F.O. Multi-Drug Resistant Escherichia coli, Biosecurity and Anti-Microbial Use in Live Bird Markets, Abeokuta, Nigeria. Antibiotics 2022, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Mag, P.; Németh, K.; Somogyi, Z.; Jerzsele, Á. Antibacterial therapy based on pharmacokinetic/ pharmacodynamic models in small animal medicine-1. Literature review. Magy. Állatorvosok Lapja 2023, 145, 419–438. [Google Scholar] [CrossRef]
- Adams, H.R. Veterinary Pharmacology and Therapeutics; Iowa State University Press: Ames, Iowa, 2001; ISBN 978-0-8138-1743-9. [Google Scholar]
- KSH Baromfiállomány (19.1.1.29.). Available online: https://www.ksh.hu/stadat_files/mez/hu/mez0029.html (accessed on 18 May 2023).
- Szoke, Z.; Fauszt, P.; Mikolas, M.; David, P.; Szilagyi-Tolnai, E.; Pesti-Asboth, G.; Homoki, J.R.; Kovacs-Forgacs, I.; Gal, F.; Stundl, L.; et al. Comprehensive Analysis of Antimicrobial Resistance Dynamics among Broiler and Duck Intensive Production Systems. Sci. Rep. 2025, 15, 4673. [Google Scholar] [CrossRef]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus Epidemiology in Wildlife: A Systematic Review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Yuko, S. Staphylococcosis in Poultry–Poultry. Available online: https://www.merckvetmanual.com/poultry/staphylococcosis/staphylococcosis-in-poultry (accessed on 3 May 2025).
- Teresa, Y.M. Streptococcosis in Poultry–Poultry. Available online: https://www.msdvetmanual.com/poultry/streptococcosis/streptococcosis-in-poultry (accessed on 3 May 2025).
- Fulde, M.; Valentin-Weigand, P. Epidemiology and Pathogenicity of Zoonotic Streptococci. Curr. Top. Microbiol. Immunol. 2013, 368, 49–81. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, J.; He, Y.; Lv, Z.; Liang, Z.; Chen, J.; Li, P.; Liu, J.; Yang, H.; Tao, A.; et al. Exploring the Role of Staphylococcus aureus in Inflammatory Diseases. Toxins 2022, 14, 464. [Google Scholar] [CrossRef]
- Abbasi, K.; Tajbakhsh, E.; Momtaz, H. Antimicrobial Resistance, Virulence Genes, and Biofilm Formation in Staphylococcus aureus Strains Isolated from Meat and Meat Products. J. Food Saf. 2021, 41, e12933. [Google Scholar] [CrossRef]
- Bortolaia, V.; Espinosa-Gongora, C.; Guardabassi, L. Human Health Risks Associated with Antimicrobial-Resistant Enterococci and Staphylococcus aureus on Poultry Meat. Clin. Microbiol. Infect. 2016, 22, 130–140. [Google Scholar] [CrossRef]
- Benrabia, I.; Hamdi, T.M.; Shehata, A.A.; Neubauer, H.; Wareth, G. Methicillin-Resistant Staphylococcus aureus (MRSA) in Poultry Species in Algeria: Long-Term Study on Prevalence and Antimicrobial Resistance. Vet. Sci. 2020, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2020/2021. EFSA J. 2023, 21, e07867. [Google Scholar] [CrossRef]
- Abdullahi, I.N.; Zarazaga, M.; Campaña-Burguet, A.; Eguizábal, P.; Lozano, C.; Torres, C. Nasal Staphylococcus aureus and S. pseudintermedius Carriage in Healthy Dogs and Cats: A Systematic Review of Their Antibiotic Resistance, Virulence and Genetic Lineages of Zoonotic Relevance. J. Appl. Microbiol. 2022, 133, 3368–3390. [Google Scholar] [CrossRef]
- Zehra, A.; Gulzar, M.; Singh, R.; Kaur, S.; Gill, J.P.S. Prevalence, Multidrug Resistance and Molecular Typing of Methicillin-Resistant Staphylococcus aureus (MRSA) in Retail Meat from Punjab, India. J. Glob. Antimicrob. Resist. 2019, 16, 152–158. [Google Scholar] [CrossRef]
- de Boer, E.; Zwartkruis-Nahuis, J.T.M.; Wit, B.; Huijsdens, X.W.; de Neeling, A.J.; Bosch, T.; van Oosterom, R.A.A.; Vila, A.; Heuvelink, A.E. Prevalence of Methicillin-Resistant Staphylococcus aureus in Meat. Int. J. Food Microbiol. 2009, 134, 52–56. [Google Scholar] [CrossRef]
- AVEC. Annual Report 2022; Association of Poultry Processors and Poultry Trade in the EU Countries: Brussels, Belgium, 2022; Available online: http://avec-poultry.eu/wp-content/uploads/2022/09/AVEC-annual-report-2022_FINAL-WEB.pdf (accessed on 3 May 2025).
- Bourély, C.; Cazeau, G.; Jarrige, N.; Jouy, E.; Haenni, M.; Lupo, A.; Madec, J.-Y.; Leblond, A.; Gay, E. Co-Resistance to Amoxicillin and Tetracycline as an Indicator of Multidrug Resistance in Escherichia coli Isolates From Animals. Front. Microbiol. 2019, 10, 2288. [Google Scholar] [CrossRef] [PubMed]
- Fessler, A.T.; Kadlec, K.; Hassel, M.; Hauschild, T.; Eidam, C.; Ehricht, R.; Monecke, S.; Schwarz, S. Characterization of Methicillin-Resistant Staphylococcus aureus Isolates from Food and Food Products of Poultry Origin in Germany. Appl. Environ. Microbiol. 2011, 77, 7151–7157. [Google Scholar] [CrossRef] [PubMed]
- Dahms, C.; Huebner, N.-O.; Kossow, A.; Mellmann, A.; Dittmann, K.; Kramer, A. Occurrence of ESBL-Producing Escherichia coli in Livestock and Farm Workers in Mecklenburg-Western Pomerania, Germany. PLoS ONE 2015, 10, e0143326. [Google Scholar] [CrossRef] [PubMed]
- Krupa, P.; Bystroń, J.; Bania, J.; Podkowik, M.; Empel, J.; Mroczkowska, A. Genotypes and Oxacillin Resistance of Staphylococcus aureus from Chicken and Chicken Meat in Poland. Poult. Sci. 2014, 93, 3179–3186. [Google Scholar] [CrossRef]
- Pyzik, E.; Marek, A.; Stępień-Pyśniak, D.; Urban-Chmiel, R.; Jarosz, Ł.S.; Jagiełło-Podębska, I. Detection of Antibiotic Resistance and Classical Enterotoxin Genes in Coagulase -Negative Staphylococci Isolated from Poultry in Poland. J. Vet. Res. 2019, 63, 183–190. [Google Scholar] [CrossRef]
- Boost, M.V.; Wong, A.; Ho, J.; O’Donoghue, M. Isolation of Methicillin-Resistant Staphylococcus aureus (MRSA) from Retail Meats in Hong Kong. Foodborne Pathog. Dis. 2013, 10, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-K.; Nam, H.-M.; Park, H.-J.; Lee, H.-S.; Choi, M.-J.; Jung, S.-C.; Lee, J.-Y.; Kim, Y.-C.; Song, S.-W.; Wee, S.-H. Prevalence and Characterization of Methicillin-Resistant Staphylococcus aureus in Raw Meat in Korea. J. Microbiol. Biotechnol. 2010, 20, 775–778. [Google Scholar] [CrossRef]
- Cha, S.-Y.; Kang, M.; Yoon, R.-H.; Park, C.-K.; Moon, O.-K.; Jang, H.-K. Prevalence and Antimicrobial Susceptibility of Salmonella Isolates in Pekin Ducks from South Korea. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; He, S.; Tan, A.; Guo, X.; Jiang, L.; Liu-Fu, C.; Deng, Y.; Zhang, R. Isolation, Identification and Character Analysis of Streptococcus dysgalactiae from Megalobrama Terminalis. J. Fish Dis. 2020, 43, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Andam, C.P.; Hanage, W.P. Mechanisms of Genome Evolution of Streptococcus. Infect. Genet. Evol. 2015, 33, 334–342. [Google Scholar] [CrossRef]
- Hakenbeck, R.; Grebe, T.; Zähner, D.; Stock, J.B. Beta-Lactam Resistance in Streptococcus pneumoniae: Penicillin-Binding Proteins and Non-Penicillin-Binding Proteins. Mol. Microbiol. 1999, 33, 673–678. [Google Scholar] [CrossRef]
- Dewachter, L.; Dénéréaz, J.; Liu, X.; de Bakker, V.; Costa, C.; Baldry, M.; Sirard, J.-C.; Veening, J.-W. Amoxicillin-Resistant Streptococcus pneumoniae Can Be Resensitized by Targeting the Mevalonate Pathway as Indicated by sCRilecs-Seq. eLife 2022, 11, e75607. [Google Scholar] [CrossRef]
- McMullin, P. A Pocket Guide to Poultry Health and Disease; 5M Books Ltd.: Sheffield, UK, 2004; ISBN 978-0-9530150-5-4. [Google Scholar]
- Hogg, R.; Pearson, A. Streptococcus gallolyticus subspecies gallolyticus Infection in Ducklings. Vet. Rec. 2009, 165, 297–298. [Google Scholar] [CrossRef]
- Logue, C.M.; Andreasen, C.B.; Borst, L.B.; Eriksson, H.; Hampson, D.J.; Sanchez, S.; Fulton, R.M. Other Bacterial Diseases. In Diseases of Poultry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 995–1085. ISBN 978-1-119-37119-9. [Google Scholar]
- Mark, P.; Paul, M.; Janet, M.B.; Dennis, A. Poultry Diseases; Elsevier Health Sciences: Edingurg, UK, 2007; ISBN 978-0-7020-2862-5. [Google Scholar]
- Rodrigo, M.K.D.; Saiganesh, A.; Hayes, A.J.; Wilson, A.M.; Anstey, J.; Pickering, J.L.; Iwasaki, J.; Hillas, J.; Winslow, S.; Woodman, T.; et al. Host-Dependent Resistance of Group A Streptococcus to Sulfamethoxazole Mediated by a Horizontally-Acquired Reduced Folate Transporter. Nat. Commun. 2022, 13, 6557. [Google Scholar] [CrossRef]
- Abreu, R.; Semedo-Lemsaddek, T.; Cunha, E.; Tavares, L.; Oliveira, M. Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control. Microorganisms 2023, 11, 953. [Google Scholar] [CrossRef] [PubMed]
- Al-Hadidi, S.H.; Al Mana, H.; Almoghrabi, S.Z.; El-Obeid, T.; AlAli, W.Q.; Eltai, N.O. Retail Chicken Carcasses as a Reservoir of Multidrug-Resistant Salmonella. Microb. Drug Resist. 2022, 28, 824–831. [Google Scholar] [CrossRef] [PubMed]
- da Costa, P.M.; Loureiro, L.; Matos, A.J.F. Transfer of Multidrug-Resistant Bacteria between Intermingled Ecological Niches: The Interface between Humans, Animals and the Environment. Int. J. Environ. Res. Public Health 2013, 10, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Hafez, H.M.; Attia, Y.A. Challenges to the Poultry Industry: Current Perspectives and Strategic Future After the COVID-19 Outbreak. Front. Vet. Sci. 2020, 7, 516. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EU Body or Agency). Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2022: Trends from 2010 to 2022: Thirteenth ESVAC Report; Publications Office of the European Union: Luxembourg, 2023; ISBN 978-92-9155-072-2. [Google Scholar]
- Bank, W. People, Pathogens and Our Planet: The Economics of One Health; World Bank: Washington, DC, USA, 2012. [Google Scholar]
- Awad, N.F.S.; Abd El-Hamid, M.I.; Nabil, N.M.; Tawakol, M.M.; Eid, S.; Al-Zaban, M.I.; Farouk, H.; Zakai, S.A.; Elkelish, A.; Ibrahim, M.S.; et al. Multidrug Resistant and Multivirulent Avian Bacterial Pathogens: Tackling Experimental Leg Disorders Using Phytobiotics and Antibiotics Alone or in Combination. Poult. Sci. 2023, 102, 102889. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Llorens, J.; Monroy, I.; Torres-Boncompte, J.; Soriano, J.M.; Catalá-Gregori, P.; Sevilla-Navarro, S. Tracking the Prevalence of Antibiotic Resistance in Enterococcus Within the Spanish Poultry Industry: Insights from a One Health Approach. Antibiotics 2024, 14, 16. [Google Scholar] [CrossRef]
- Wang, J.; Meng, Y.; Zhang, R.; Yan, H.; Xu, G.; Zhu, Y.; Xie, Z.; Jiang, S. Coagulase-Negative Staphylococci Are the Main Causes of Bacterial Meningitis in Duck. Poult. Sci. 2024, 103, 103592. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.R.N.; Rosa, I.M.L. Biodiversity, Traditional Medicine and Public Health: Where Do They Meet? J. Ethnobiol. Ethnomed. 2007, 3, 14. [Google Scholar] [CrossRef]
- Eid, H.M.; Algammal, A.M.; Elfeil, W.K.; Youssef, F.M.; Harb, S.M.; Abd-Allah, E.M. Prevalence, Molecular Typing, and Antimicrobial Resistance of Bacterial Pathogens Isolated from Ducks. Vet. World 2019, 12, 677–683. [Google Scholar] [CrossRef]
- Xu, Z.; Shah, H.N.; Misra, R.; Chen, J.; Zhang, W.; Liu, Y.; Cutler, R.R.; Mkrtchyan, H.V. The Prevalence, Antibiotic Resistance and mecA Characterization of Coagulase Negative Staphylococci Recovered from Non-Healthcare Settings in London, UK. Antimicrob. Resist. Infect. Control 2018, 7, 73. [Google Scholar] [CrossRef]
- Kolár, M.; Urbánek, K.; Látal, T. Antibiotic Selective Pressure and Development of Bacterial Resistance. Int. J. Antimicrob. Agents 2001, 17, 357–363. [Google Scholar] [CrossRef]
- Leclercq, R.; Courvalin, P. Bacterial Resistance to Macrolide, Lincosamide, and Streptogramin Antibiotics by Target Modification. Antimicrob. Agents Chemother. 1991, 35, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Okitsu, N.; Kaieda, S.; Yano, H.; Nakano, R.; Hosaka, Y.; Okamoto, R.; Kobayashi, T.; Inoue, M. Characterization of ermB Gene Transposition by Tn1545 and Tn917 in Macrolide-Resistant Streptococcus pneumoniae Isolates. J. Clin. Microbiol. 2005, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Gurjar, M. Colistin for Lung Infection: An Update. J. Intensive Care 2015, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute CLSI. Methods for Antimicrobial Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria Isolated From Animals, 1st ed.; CLSI Standards VET06; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical and Laboratory Standards Institute CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 12th ed.; CLSI standards M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2024. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Heck, D.W.; Moshagen, M. RRreg: Correlation and Regression Analyses for Randomized Response Data; R package Version 1.1.0, 2022. Available online: https://CRAN.R-project.org/package=RRreg (accessed on 3 May 2025).



{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Pericardium | Lungs | Liver | Articulation |
|---|---|---|---|---|
| n (%) | ||||
| Staphylococcus aureus (n = 8) | 0 (0.0) | 0 (0.0) | 6 (75.0) | 2 (25.0) |
| Streptococcus spp. (n = 19) | 2 (10.5) | 1 (5.3) | 15 (78.9) | 1 (5.3) |
| Total (n = 27) | 2 (7.4) | 1 (3.7) | 21 (77.8) | 3 (11.1) |
| Antibiotics | Breakpoint 1 (μg/mL) | Distribution of Strains by MIC (μg/mL) | MIC50 | MIC90 | ECOFF 4 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.015 | 0.03 | 0.06 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | µg/mL | ||||
| Amoxicillin | 0.5 | 2 | 0 | 1 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0.5 | 64 | 0.5 | |||
| Amoxicillin–clavulanic acid 2 | 1 | 2 | 1 | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0.25 | 4 | 0.5 | |||||||
| Ceftriaxone | - | 3 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 16 | 8 | ||||
| Colistin | - | 1 | 4 | 0 | 0 | 0 | 1 | 2 | 32 | 1024 | - | ||||||||||
| Doxycycline | 0.5 | 3 | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 1 | 2 | 8 | 0.5 | ||||||||
| Enrofloxacin | 4 | 1 | 0 | 2 | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 4 | - | ||||||
| Florfenicol | - | 2 | 1 | 1 | 2 | 1 | 0 | 0 | 0 | 0 | 1 | 4 | 16 | 8 | |||||||
| Imipenem | 8 | 1 | 3 | 2 | 1 | 0 | 0 | 0 | 1 | 0.03 | 0.125 | 0.125 | |||||||||
| Lincomycin | - | 2 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 4 | 512 | 2 | ||||||
| Neomycin | - | 1 | 3 | 2 | 0 | 1 | 1 | 32 | 256 | 1 | |||||||||||
| Trimethoprim–sulfamethoxazole 3 | 4 | 4 | 2 | 0 | 0 | 0 | 1 | 1 | 1 | 32 | 0.25 | ||||||||||
| Spectinomycin | - | 1 | 4 | 0 | 1 | 1 | 1 | 64 | 512 | 128 | |||||||||||
| Tiamulin | 4 | 1 | 1 | 0 | 2 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 4 | 128 | 2 | |||||
| Tilozin | 64 | 3 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 2 | 128 | - | |||
| Vancomycin | 32 | 4 | 1 | 1 | 2 | 0.5 | 4 | 2 | |||||||||||||
| Antibiotics | Breakpoint 1 (μg/mL) | Distribution of Strains by MIC (μg/mL) | MIC50 | MIC90 | ECOFF 4 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.015 | 0.03 | 0.06 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | µg/mL | ||||
| Amoxicillin | - | 1 | 2 | 3 | 0 | 5 | 3 | 2 | 0 | 3 | 0.5 | 8 | 0.06 5 | ||||||||
| Amoxicillin–clavulanic acid 2 | 1 | 1 | 2 | 3 | 0 | 5 | 2 | 0 | 0 | 6 | 0.5 | 8 | 0.06 5 | ||||||||
| Ceftriaxone | - | 4 | 6 | 2 | 0 | 1 | 1 | 1 | 3 | 0 | 0 | 1 | 0.06 | 4 | 0.06 6 | ||||||
| Colistin | - | 1 | 1 | 0 | 0 | 8 | 6 | 0 | 2 | 0 | 0 | 1 | 16 | 128 | - | ||||||
| Doxycycline | 0.5 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 6 | 7 | 0 | 1 | 16 | 32 | 0.5 7 | ||||
| Enrofloxacin | 4 | 2 | 6 | 4 | 2 | 2 | 1 | 1 | 1 | 1 | 16 | 2 8 | |||||||||
| Florfenicol | 8 | 1 | 0 | 4 | 2 | 5 | 0 | 1 | 1 | 2 | 1 | 2 | 8 | 512 | 4 7 | ||||||
| Imipenem | - | 8 | 3 | 4 | 0 | 0 | 3 | 1 | 0.06 | 1 | - | ||||||||||
| Lincomycin | - | 1 | 0 | 0 | 1 | 2 | 0 | 1 | 1 | 1 | 1 | 3 | 0 | 3 | 5 | 128 | 1024 | 1 9 | |||
| Neomycin | - | 14 | 1 | 1 | 3 | 64 | 512 | - | |||||||||||||
| Trimethoprim–sulfamethoxazole 3 | 4 | 2 | 0 | 1 | 1 | 5 | 2 | 3 | 0 | 0 | 5 | 4 | 64 | 0.25 7 | |||||||
| Spectinomycin | - | 10 | 2 | 3 | 1 | 3 | 64 | 1024 | - | ||||||||||||
| Tiamulin | - | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | 2 | 8 | 1 | 0 | 3 | 128 | 1024 | - | ||||
| Tilozin | - | 4 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 3 | 3 | 64 | 1024 | 1 8 | |||
| Vancomycin | 2 | 2 | 3 | 13 | 0 | 0 | 1 | 0.5 | 0.5 | 1 6 | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerek, Á.; Szabó, Á.; Jerzsele, Á. Antimicrobial Susceptibility Profiles of Staphylococcus aureus and Streptococcus spp. Isolates from Clinical Cases of Waterfowl in Hungary Between 2022 and 2023. Antibiotics 2025, 14, 496. https://doi.org/10.3390/antibiotics14050496
Kerek Á, Szabó Á, Jerzsele Á. Antimicrobial Susceptibility Profiles of Staphylococcus aureus and Streptococcus spp. Isolates from Clinical Cases of Waterfowl in Hungary Between 2022 and 2023. Antibiotics. 2025; 14(5):496. https://doi.org/10.3390/antibiotics14050496
Chicago/Turabian StyleKerek, Ádám, Ábel Szabó, and Ákos Jerzsele. 2025. "Antimicrobial Susceptibility Profiles of Staphylococcus aureus and Streptococcus spp. Isolates from Clinical Cases of Waterfowl in Hungary Between 2022 and 2023" Antibiotics 14, no. 5: 496. https://doi.org/10.3390/antibiotics14050496
APA StyleKerek, Á., Szabó, Á., & Jerzsele, Á. (2025). Antimicrobial Susceptibility Profiles of Staphylococcus aureus and Streptococcus spp. Isolates from Clinical Cases of Waterfowl in Hungary Between 2022 and 2023. Antibiotics, 14(5), 496. https://doi.org/10.3390/antibiotics14050496