
Antimicrobial Resistance and Phylogenetic Analysis of Multidrug-Resistant Non-Typhoidal Salmonella Isolates from Different Sources in Southern Vietnam

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Results
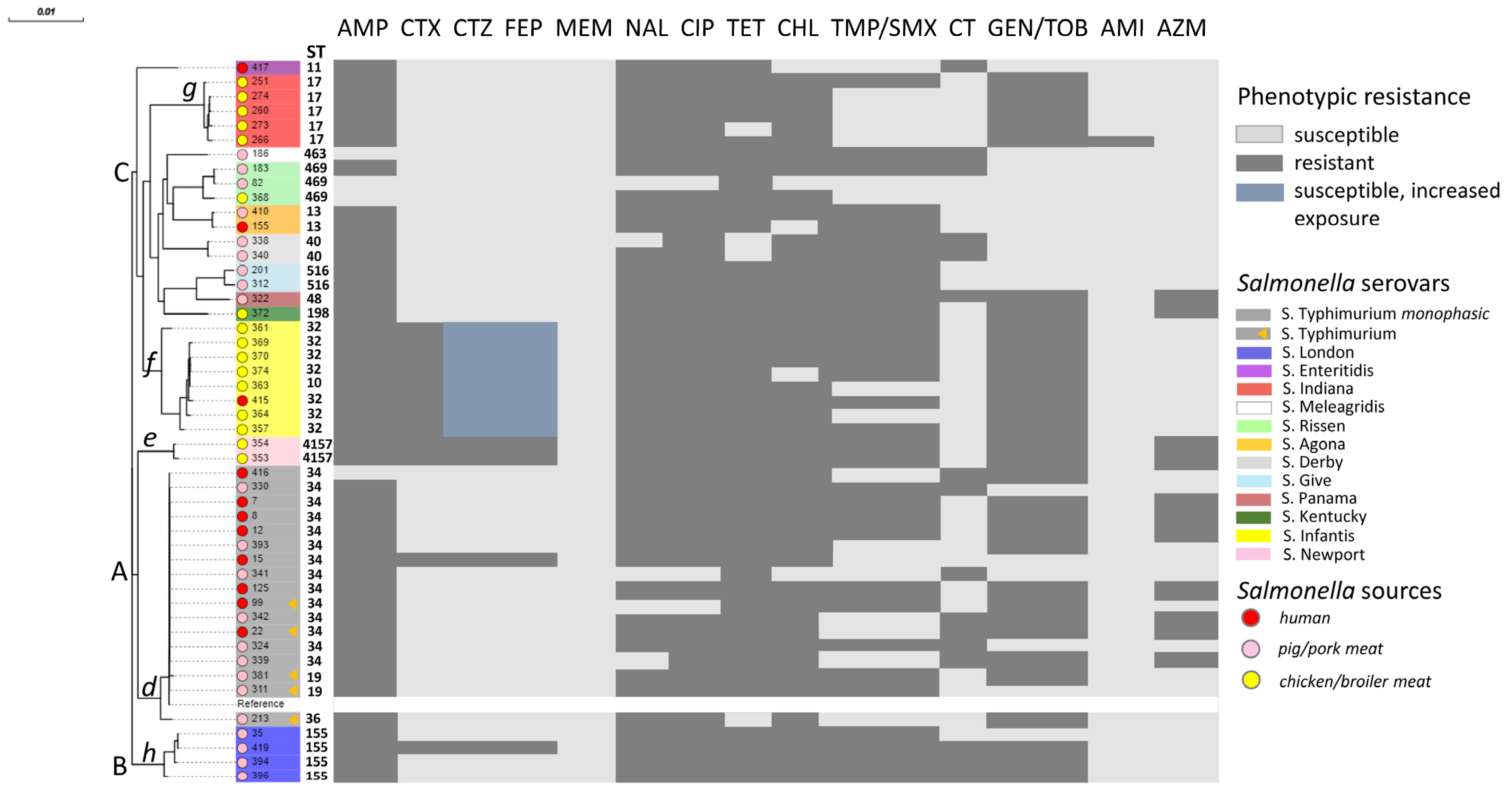
2.1. NTS Serotype Identification
2.2. Antimicrobial Susceptibility Testing (AST) of NTS
2.3. Correlation Between Phenotypic and Genotypic Antimicrobial Resistance of Salmonella Isolates
2.3.1. Beta-Lactam Antibiotics
2.3.2. Aminoglycosides
2.3.3. Quinolones
2.3.4. Other Antibiotics
2.4. Plasmid Analysis
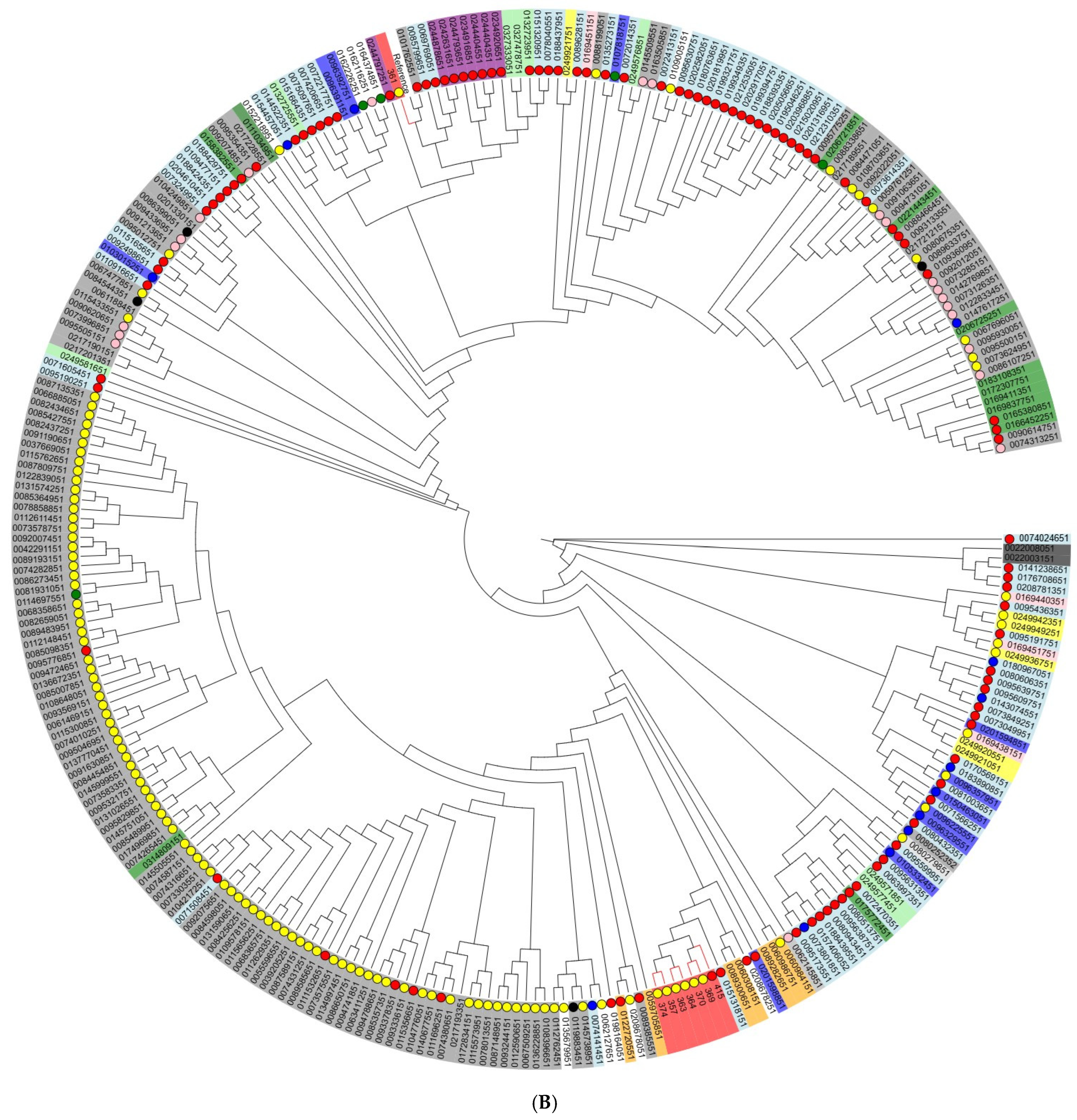
2.5. Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Study Design, Sampling Collection, and Bacterial Isolation
4.2. Phenotypic Antimicrobial Susceptibility Testing (AST)
4.3. DNA Extraction and Sequencing
4.4. Bioinformatics Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NTS | Non-typhoidal Salmonella |
| WGS | Whole-genome sequencing |
| MDR | Multidrug resistance |
| ARG | Antimicrobial resistance genes |
| AST | Antimicrobial susceptibility testing |
| WT | Wild-type |
| NWT | Non-wild-type |
| BLs | Beta-lactamases |
| AMP | Ampicillin |
| CTZ | Ceftazidime |
| CTX | Cefotaxime |
| FEP | Cefepime |
| MEM | Meropenem |
| NAL | Nalidixic acid |
| CIP | Ciprofloxacin |
| TET | Tetracycline |
| CHL | Chloramphenicol |
| TMP/SMX | Trimethoprim/Sulfamethoxazole |
| CT | Colistin |
| GEN | Gentamycin |
| TOB | Tobramycin |
| AMI | Amikacin |
| AZM | Azithromycin |
References
- Nhung, N.T.; Phu, D.H.; Carrique-Mas, J.J.; Padungtod, P. A review and meta-analysis of non-typhoidal Salmonella in Vietnam: Challenges to the control and antimicrobial resistance traits of a neglected zoonotic pathogen. One Health 2024, 18, 100698. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, H.; Excler, J.-L.; Kim, J.H.; Lee, J.-S. The Economic Burden of Non-Typhoidal Salmonella and Invasive Non-Typhoidal Salmonella Infection: A Systematic Literature Review. Vaccines 2024, 12, 758. [Google Scholar] [CrossRef] [PubMed]
- Billah, M.M.; Rahman, M.S. Salmonella in the environment: A review on ecology, antimicrobial resistance, seafood contaminations, and human health implications. J. Hazard. Mater. Adv. 2024, 13, 100407. [Google Scholar] [CrossRef]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- Acosta, A.; Tirkaso, W.; Nicolli, F.; Van Boeckel, T.P.; Cinardi, G.; Song, J. The future of antibiotic use in livestock. NaNat. Commun. 2025, 16, 2469. [Google Scholar] [CrossRef]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Sima, C.M.; Buzilă, E.R.; Trofin, F.; Păduraru, D.; Luncă, C.; Duhaniuc, A.; Dorneanu, O.S.; Nastase, E.V. Emerging Strategies against Non-Typhoidal Salmonella: From Pathogenesis to Treatment. Curr. Issues Mol. Biol. 2024, 46, 7447–7472. [Google Scholar] [CrossRef]
- Kumar, G.; Kumar, S.; Jangid, H.; Dutta, J.; Shidiki, A. The rise of non-typhoidal Salmonella: An emerging global public health concern. Front. Microbiol. 2025, 16, 1524287. [Google Scholar] [CrossRef]
- Hengkrawit, K.; Tangjade, C. Prevalence and Trends in Antimicrobial Susceptibility Patterns of Multi-Drug-Resistance Non-Typhoidal Salmonella in Central Thailand, 2012–2019. Infect. Drug Resist. 2022, 15, 1305–1315. [Google Scholar] [CrossRef]
- Sahu, A.A.; Sephalika, S.; Mohakud, N.K.; Sahu, B.R. Prevalence and Multidrug Resistance in Non-Typhoidal Salmonella in India: A 20-Year Outlook. Acta Microbiol. Hell. 2025, 70, 6. [Google Scholar] [CrossRef]
- Tuat, C.V.; Hue, P.T.; Loan, N.T.P.; Thuy, N.T.; Hue, L.T.; Giang, V.N.; Erickson, V.I.; Padungtod, P. Antimicrobial Resistance Pilot Surveillance of Pigs and Chickens in Vietnam, 2017–2019. Front. Vet. Sci. 2021, 8, 618497. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, S.; Duong, V.T.; Tuyen, H.T.; Campbell, J.I.; Thomson, N.R.; Parkhill, J.; Le Phuc, H.; Chau, T.T.H.; Maskell, D.J.; Perron, G.G.; et al. Mobility of antimicrobial resistance across serovars and disease presentations in non-typhoidal Salmonella from animals and humans in Vietnam. Microb. Genom. 2022, 8, mgen000798. [Google Scholar] [CrossRef]
- Holohan, N.; Wallat, M.; Luu, T.H.Y.; Clark, E.; Truong, D.T.Q.; Xuan, S.D.; Vu, H.T.K.; Van Truong, D.; Huy, H.T.; Nguyen-Viet, H.; et al. Analysis of Antimicrobial Resistance in Non-typhoidal Salmonella Collected From Pork Retail Outlets and Slaughterhouses in Vietnam Using Whole Genome Sequencing. Front. Vet. Sci. 2022, 9, 816279. [Google Scholar] [CrossRef] [PubMed]
- Coyne, L.; Arief, R.; Benigno, C.; Giang, V.N.; Huong, L.Q.; Jeamsripong, S.; Kalpravidh, W.; McGrane, J.; Padungtod, P.; Patrick, I.; et al. Characterizing Antimicrobial Use in the Livestock Sector in Three South East Asian Countries (Indonesia, Thailand, and Vietnam). Antibiotics 2019, 8, 33. [Google Scholar] [CrossRef]
- Redondo-Salvo, S.; Fernández-López, R.; Ruiz, R.; Vielva, L.; DE Toro, M.; Rocha, E.P.C.; Garcillán-Barcia, M.P.; De La Cruz, F. Pathways for horizontal gene transfer in bacteria revealed by a global map of their plasmids. Nat. Commun. 2020, 11, 3602. [Google Scholar] [CrossRef]
- Commichaux, S.; Rand, H.; Javkar, K.; Molloy, E.K.; Pettengill, J.B.; Pightling, A.; Hoffmann, M.; Pop, M.; Jayeola, V.; Foley, S.; et al. Assessment of plasmids for relating the 2020 Salmonella enterica serovar Newport onion outbreak to farms implicated by the outbreak investigation. BMC Genom. 2023, 24, 165. [Google Scholar] [CrossRef]
- Yan, S.; Jiang, Z.; Zhang, W.; Liu, Z.; Dong, X.; Li, D.; Liu, Z.; Li, C.; Liu, X.; Zhu, L. Genomes-based MLST, cgMLST, wgMLST and SNP analysis of Salmonella Typhimurium from animals and humans. Comp. Immunol. Microbiol. Infect. Dis. 2023, 96, 101973. [Google Scholar] [CrossRef] [PubMed]
- Mather, A.E.; Phuong, T.L.T.; Gao, Y.; Clare, S.; Mukhopadhyay, S.; Goulding, D.A.; Hoang, N.T.D.; Tuyen, H.T.; Lan, N.P.H.; Thompson, C.N.; et al. New Variant of Multidrug-Resistant Salmonella enterica Serovar Typhimurium Associated with Invasive Disease in Immunocompromised Patients in Vietnam. mBio 2018, 9, e01056-18. [Google Scholar] [CrossRef]
- Soliani, L.; Rugna, G.; Prosperi, A.; Chiapponi, C.; Luppi, A. Salmonella Infection in Pigs: Disease, Prevalence, and a Link between Swine and Human Health. Pathogens 2023, 12, 1267. [Google Scholar] [CrossRef]
- Tu, L.T.P.; Hoang, N.V.M.; Cuong, N.V.; Campbell, J.; Bryant, J.E.; Hoa, N.T.; Kiet, B.T.; Thompson, C.; Duy, D.T.; Phat, V.V.; et al. High levels of contamination and antimicrobial-resistant non-typhoidal Salmonella serovars on pig and poultry farms in the Mekong Delta of Vietnam. Epidemiol. Infect. 2015, 143, 3074–3086. [Google Scholar] [CrossRef]
- Nguyen, D.T.A.; Kanki, M.; Nguyen, P.D.; Le, H.T.; Ngo, P.T.; Tran, D.N.M.; Le, N.H.; Van Dang, C.; Kawai, T.; Kawahara, R.; et al. Prevalence, antibiotic resistance, and extended-spectrum and AmpC beta-lactamase productivity of Salmonella isolates from raw meat and seafood samples in Ho Chi Minh City, Vietnam. Int. J. Food Microbiol. 2016, 236, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Ta, Y.T.; Nguyen, T.T.; To, P.B.; Pham, D.X.; Le, H.T.H.; Thi, G.N.; Alali, W.Q.; Walls, I.; Doyle, M.P. Quantification, serovars, and antibiotic resistance of salmonella isolated from retail raw chicken meat in Vietnam. J. Food Prot. 2014, 77, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Leinyuy, J.F.; Ali, I.M.; Ousenu, K.; Tume, C.B. Molecular characterization of antimicrobial resistance related genes in E. coli, Salmonella and Klebsiella isolates from broilers in the West Region of Cameroon. PLoS ONE 2023, 18, e0280150. [Google Scholar] [CrossRef] [PubMed]
- Nhung, N.T.; Van, N.T.B.; Van Cuong, N.; Duong, T.T.Q.; Nhat, T.T.; Hang, T.T.T.; Nhi, N.T.H.; Kiet, B.T.; Hien, V.B.; Ngoc, P.T.; et al. Antimicrobial residues and resistance against critically important antimicrobials in non-typhoidal Salmonella from meat sold at wet markets and supermarkets in Vietnam. Int. J. Food Microbiol. 2017, 266, 301–309. [Google Scholar] [CrossRef]
- Da Pham, X.; Hong, H.L.T.; Thanh, H.T.T.; Le, L.T.; Le, H.V.; Thi, N.H.; Le Tran, M.; Trung, N.T. Strains and virulence genes of salmonella with multidrug resistance isolated from chicken carcasses (Ha-noi, Vietnam). Health Risk Analysis 2023, 1, 115–123. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Le, H.V.; Hai, H.V.T.; Tuan, T.N.; Nguyen, H.M.; Xuan, D.P.; Thanh, H.T.T.; Le Thi, H.H. Whole-Genome Analysis of Antimicrobial-Resistant Salmonella enterica Isolated from Duck Carcasses in Hanoi, Vietnam. Curr. Issues Mol. Biol. 2023, 45, 2213–2229. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef]
- Song, Q.; Xu, Z.; Gao, H.; Zhang, D. Overview of the development of quinolone resistance in Salmonella species in China, 2005-2016. Infect. Drug Resist. 2018, 11, 267–274. [Google Scholar] [CrossRef]
- Pribul, B.R.; Festivo, M.L.; Rodrigues, M.S.; Costa, R.G.; Rodrigues, E.C.d.P.; de Souza, M.M.S.; Rodrigues, D.d.P. Characteristics of Quinolone Resistance in Salmonella spp. Isolates from the Food Chain in Brazil. Front. Microbiol. 2017, 8, 299. [Google Scholar] [CrossRef]
- Weng, R.; Gu, Y.; Zhang, W.; Hou, X.; Wang, H.; Tao, J.; Deng, M.; Zhou, M.; Zhao, Y. Corrigendum: Whole-genome sequencing provides insight into antimicrobial resistance and molecular characteristics of Salmonella from livestock meat and diarrhea patient in Hanzhong, China. Front. Microbiol. 2022, 13, 981414. [Google Scholar] [CrossRef]
- Kotb, D.N.; Mahdy, W.K.; Mahmoud, M.S.; Khairy, R.M.M. Impact of co-existence of PMQR genes and QRDR mutations on fluoroquinolones resistance in Enterobacteriaceae strains isolated from community and hospital acquired UTIs. BMC Infect. Dis. 2019, 19, 979. [Google Scholar] [CrossRef] [PubMed]
- Navickaite, I.; Holmes, H.; Dondi, L.; Randall, L.; Fearnley, C.; Taylor, E.; Fullick, E.; Horton, R.; Williamson, S.; AbuOun, M.; et al. Occurrence and characterization of rmtB-harbouring Salmonella and Escherichia coli isolates from a pig farm in the UK. J. Antimicrob. Chemother. 2024, 79, 1329–1336. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.-Y.; Wang, Y.; Sun, F.; Li, W.; Wu, H.; Shen, P.-C.; Pan, Z.-M.; Jiao, X. Emergence of 16S rRNA Methylase Gene rmtB in Salmonella Enterica Serovar London and Evolution of RmtB-Producing Plasmid Mediated by IS26. Front. Microbiol. 2021, 11, 604278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, J.; Yang, X.; Wu, Y.; Wang, Z.; Xu, Y.; Zhou, L.; Wang, J.; Jiao, X.; Sun, L. Emerging Mobile Colistin Resistance Gene Mcr-1 and Mcr-10 in Enterobacteriaceae Isolates From Urban Sewage in China. Infect. Drug Resist. 2025, 18, 1035–1048. [Google Scholar] [CrossRef]
- Fortini, D.; Owczarek, S.; Dionisi, A.M.; Lucarelli, C.; Arena, S.; Carattoli, A.; Enter-Net Italia Colistin Resistance Study Group; Villa, L.; García-Fernández, A. Colistin Resistance Mechanisms in Human Salmonella enterica Strains Isolated by the National Surveillance Enter-Net Italia (2016–2018). Antibiotics 2022, 11, 102. [Google Scholar] [CrossRef]
- Pavelquesi, S.L.S.; Ferreira, A.C.A.d.O.; Rodrigues, A.R.M.; Silva, C.M.d.S.; Orsi, D.C.; da Silva, I.C.R. Presence of Tetracycline and Sulfonamide Resistance Genes in Salmonella spp.: Literature Review. Antibiotics 2021, 10, 1314. [Google Scholar] [CrossRef] [PubMed]
- Bryan, A.; Shapir, N.; Sadowsky, M.J. Frequency and distribution of tetracycline resistance genes in genetically diverse, nonselected, and nonclinical Escherichia coli strains isolated from diverse human and animal sources. Appl. Environ. Microbiol. 2004, 70, 2503–2507. [Google Scholar] [CrossRef]
- Jurado-Rabadán, S.; de la Fuente, R.; A Ruiz-Santa-Quiteria, J.; Orden, J.A.; de Vries, L.E.; Agersø, Y. Detection and linkage to mobile genetic elements of tetracycline resistance gene tet(M) in Escherichia coli isolates from pigs. BMC Vet. Res. 2014, 10, 155. [Google Scholar] [CrossRef]
- Chia-Wei, L.; Cheng, J.-F.; Tung, K.-C.; Hong, Y.-K.; Lin, J.-H.; Lin, Y.-H.; Tsai, C.-A.; Lin, S.-P.; Chen, Y.-C.; Shi, Z.-Y.; et al. Evolution of trimethoprim/sulfamethoxazole resistance in Shewanella algae from the perspective of comparative genomics and global phylogenic analysis. J. Microbiol. Immunol. Infect. 2022, 55, 1195–1202. [Google Scholar] [CrossRef]
- ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of salmonella spp. International Organization for Standardization: Geneva, Switzerland, 2017.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serotype | ST | Number | Source | Place and Year of Isolation |
|---|---|---|---|---|
| S. Typhimurium monophasic (n = 12) | 34 | 6 | human | Ho Chi Minh city, 2012 (n = 1); Dong Nai, 2013 (n = 4), Ben Tre, 2018 (n = 1) |
| 6 | pig | Dong Nai, 2012 (n = 2), 2021 (n = 4) | ||
| S. Typhimurium (n = 5) | 34 | 2 | human | Dong Nai, 2012, 2013 |
| 36 | 1 | pig | Ho Chi Minh city, 2014 | |
| 19 | 1 | pork | Ho Chi Minh city, 2021 | |
| 1 | pig | Dong Nai, 2012 | ||
| S. Indiana (n = 5) | 17 | 5 | chicken | Dong Nai, 2012 |
| S. London (n = 4) | 155 | 2 | pig | Dong Nai, 2022 |
| 2 | pork | Ho Chi Minh city, 2014, 2021 | ||
| S. Infantis (n = 8) | 32 | 1 | human | An Giang, 2021 |
| 6 | broiler meat | Ho Chi Minh city, 2021 | ||
| 10 | 1 | broiler meat | Ho Chi Minh city, 2021 | |
| S. Rissen (n = 3) | 469 | 1 | pig | Ho Chi Minh city, 2014 |
| 1 | pork | Long An, 2013 | ||
| 1 | broiler meat | Ho Chi Minh city, 2021 | ||
| S. Newport (n = 2) | 4157 | 2 | broiler meat | Ho Chi Minh city, 2021 |
| S. Derby (n = 2) | 40 | 2 | pig | Dong Nai, 2021 |
| S. Give (n = 2) | 16 | 2 | pig | Dong Nai, 2021 Ho Chi Minh city, 2014 |
| S. Enteritidis (n = 1) | 11 | 1 | human | An Giang, 2020 |
| S. Meleagridis (n = 1) | 463 | 1 | pork | Ho Chi Minh city, 2014 |
| S. Kentucky (n = 1) | 198 | 1 | broiler meat | Ho Chi Minh city, 2021 |
| S. Panama (n = 1) | 48 | 1 | pig | Dong Nai, 2012 |
| S. Agona (n = 2) | 13 | 1 | human | An Giang, 2021 |
| 1 | pig | Ho Chi Minh city, 2014 |
| Antimicrobials | Resistant Isolates, n (%) | |||||
|---|---|---|---|---|---|---|
| Human n = 11 | Pig n = 18 | Chicken n = 5 | Broiler Meat n = 11 | Pork Meat n = 4 | Total n = 49 | |
| Ampicillin | 10 (90.9) | 17 (94.4) | 5 (100) | 10 (90.9) | 3 (75.0) | 45 (91.8) |
| Cefotaxime | 2 (18.2) | 0 | 0 | 9 (81.8) | 1 (25.0) | 12 (24.5) |
| Ceftazidime | 1 (9.1) | 0 | 0 | 2 (18.2) | 1 (25.0) | 4 (8.2) |
| Cefepime | 1 (9.1) | 0 | 0 | 2 (18.2) | 1 (25.0) | 4 (8.2) |
| Nalidixic acid | 10 (90.9) | 15 (83.3) | 5 (100) | 11 (100) | 3 (75.0) | 44 (89.8) |
| Ciprofloxacin | 10 (90.9) | 17 (94.4) | 5 (100) | 11 (100) | 3 (75) | 46 (93.9) |
| Tetracycline | 11 (100) | 15 (83.3) | 4 (80.0) | 11 (100) | 4 (100) | 45 (91.8) |
| Chloramphenicol | 10 (90.9) | 16 (88.9) | 5 (100) | 10 (90.9) | 3 (75.0) | 44 (89.8) |
| Trimethoprim/Sulfamethoxazole | 7 (63.6) | 13 (72.2) | 1 (20.0) | 8 (72.7) | 3 (75.0) | 32 (65.3) |
| Colistin | 3 (27.3) | 12 (66.7) | 0 | 0 | 1 (25.0) | 16 (32.7) |
| Gentamycin/Tobramycin | 8 (72.7) | 7 (38.9) | 5 (100) | 10 (90.9) | 2 (50.0) | 32 (65.3) |
| Amikacin | 0 | 0 | 1 (20.0) | 0 | 0 | 1 (2.0) |
| Azithromycin | 5 (45.5) | 3 (16.7) | 0 | 3 (27.3) | 0 | 11 (22.4) |
| Genotype | MIC. µg/mL | Source | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Human (n = 11) | Pig (n = 18) | Chicken (n = 5) | Broiler Meat (n = 11) | Pork Meat (n = 4) | ||||||
| AMP (R > 8) | CTX (R > 2) | CTZ (R > 4) | FEP (R > 4) | |||||||
| No genes | 1–8 | 0.047–0.125 | 0.047–0.125 | 0.016–0.19 | 1 | 1 | 1 | 1 | ||
| OXA-1 | >512 | 0.125–0.19 | 0.19–0.25 | 0.25–0.38 | 1 | 1 | ||||
| TEM-1B | >512 | 0.047–0.25 | 0.125–1 | 0.032–0.25 | 6 | 16 | 1 | 1 | ||
| TEM-1B + SHV-25 | >512 | 0.94 * | 0.38 | 0.064 | 1 | |||||
| TEM-1B + OXA-1 | >512 | 0.125–0.25 | 0.25–0.5 | 0.38–1 | 5 | |||||
| TEM-1B + LAP-2 | >512 | 0.25 | 0.38 | 0.19 | 1 | |||||
| CTX-M-65 | >512 | >32 | 1.5–2 | 2–4 | 1 | 6 | ||||
| CTX-M-65 + TEM-1B | >512 | >32 | 2 | 3 | 1 | |||||
| CTX-M-55 | >512 | >32 | 256 | 48 | 1 | |||||
| CTX-M-55 + TEM-1B | >512 | >32 | 24 | 24 | 1 | |||||
| CTX-M-55 + TEM-1B + LAP-2 | >512 | >32 | 16–256 | 16–128 | 1 | 1 | ||||
| GEN (R > 2) | TOB (R > 2) | AMI (R > 8) | ||||||||
| aac(6′)-Iaa + aph(6)-Id | 0.75 | 0.75 | 2 | 1 | ||||||
| aac(6′)-Iaa + aph(3″)-Ib + aph(6)-Id | 0.25–0.75 | 0.5–1.5 | 1.5–2 | 3 | 2 | 0 | ||||
| aac(6′)-Iaa + aadA | 0.5–1 | 0.5–1.5 | 1.5–3 | 7 | 1 | |||||
| aac(6′)-Iaa + aph(3″)-Ib + aph(6)-Id + aadA | 0.5–1 | 1 | 2 | 2 | ||||||
| aac(6′)-Iaa + aadA + aadB | 0.75 | 1.5 | 3 | 1 | ||||||
| aac(6′)-Iaa + aac3IId + aadA | 3–256 | 3–6 | 2 | 1 | 1 | |||||
| aac(6′)-Iaa + aac3IId + aph(3′)-Ia + aadA | 4–48 | 3–4 | 2 | 2 | ||||||
| aac(6′)-Iaa + aac3IId + aph(3″)-Ib + aph(6)-Id | 32–64 | 3–4 | 1.5–3 | 2 | 2 | |||||
| aac(6′)-Iaa + aac3IId + aph(3″)-Ib + aph(6)-Id + aadA | 257 | 16 | 2 | 1 | ||||||
| aac(6′)-Iaa + aac3IId + aph(3′)-Ia + aph(6)-Id + aadA | 24–32 | 3–6 | 1.5–2 | 1 | 1 | |||||
| aac(6′)-Iaa + aac3IId + aph(3′)-Ia + aph(3″)-Ib + aph(6)-Id + aadA | 6–64 | 4–48 | 1.5–2 | 3 | 1 | 1 | ||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aadA | 8–12 | 16–24 | 1.5–3 | 2 | ||||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aph(3′)-Ia + aadA | 4–8 | 12–24 | 1.5–3 | 1 | 5 | |||||
| aac(6′)-Iaa + aac(3)-IV + aac3IId + aph(4)-Ia + aph(3′)-Ia + aph(6)-Id + aadA | 48 | 32 | 1.5 | 1 | ||||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aph(3″)-Ib + aph(6)-Id + aac(6′)-lb-cr | 12–16 | 48–256 | 4 | 3 | ||||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aadA + aac(6′)-lb-cr | 16 | 64–256 | 3–4 | 1 | 1 | |||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aph(3′)-Ia + aadA + aac(6′)-lb-cr | 3.0 | 8 | 3 | 1 | ||||||
| aac(6′)-Iaa + aac(3)-IV + aph(4)-Ia + aph(3″)-Ib + aph(6)-Id + aac(6′)-lb-cr + rmtB | >256 | >256 | 256 | 1 | ||||||
| CIP (R > 0.06) | NAL (ECOFF 8.0) | |||||||||
| No genes | 0.03 | 8 | 1 | |||||||
| aac(6′)-Ib-cr | 0.06 | 6 | 1 | |||||||
| parC(T57S) + qnrA1 | 0.02 | 6 | 1 | |||||||
| qnrS1 | 0.13–1.5 | 8–256 | 7 | 6 | 2 | 1 | ||||
| parC(T57S) + qnrS1 | 0.09–0.75 | 8–96 | 1 | 8 | 1 | 2 | ||||
| qnrS1 + aac(6′)-Ib-cr | 1.5 | 48 | 1 | |||||||
| gyrA(D87Y) | 0.13 | >256 | 1 | |||||||
| gyrA(D87Y) + parC(T57S) | 0.19–0.25 | >256 | 1 | 6 | ||||||
| gyrA(S83Y) + parC(T57S) + qnrS1 | 0.38–0.5 | >256 | 2 | |||||||
| gyrA(D87Y) + parC(T57S) + qnrS1 | 0.13 | >256 | 1 | |||||||
| gyrA(S83F) + gyrA(D87N) + parC(T57S) + parC(S80I) | >32 | >256 | 1 | |||||||
| gyrA(S83F) + gyrA(D87G) + parC(T57S) + parC(S80R) + aac(6′)-Ib-cr | >32 | >256 | 4 | |||||||
| gyrA(S83F) + gyrA(D87G) + parC(T57S) + parC(S80R) + qnrS1 + aac(6′)-Ib-cr | >32 | >256 | 1 | |||||||
| CHL (ECOFF 16.0) | ||||||||||
| No genes | 2–16 | 1 | 2 | 1 | 1 | |||||
| cmlA1 | 64–192 | 4 | ||||||||
| cmlA1 + catB3 | 192 | 1 | ||||||||
| floR | >256 | 4 | 5 | 9 | 1 | |||||
| floR + catA2 | >256 | 4 | ||||||||
| floR + catA3 | >256 | 1 | ||||||||
| floR + cmlA1 | >256 | 1 | 6 | 1 | ||||||
| floR + cmlA1 + catA2 | >256 | 1 | ||||||||
| floR + cmlA1 + catA3 | >256 | 1 | ||||||||
| floR + cmlA1 + catB3 | >256 | 1 | ||||||||
| floR + catA1 + catB3 | >256 | 4 | ||||||||
| TMP/SMX (R > 4) | ||||||||||
| No genes | 0.05 | 1 | ||||||||
| sul1 | 0.06–0.25 | 2 | 1 | |||||||
| sul2 | 0.13–0.5 | 4 | 4 | |||||||
| sul1 + sul2 | 0.19–0.25 | 4 | ||||||||
| sul1 + sul2 + sul3 | 0.09 | 1 | ||||||||
| dfrA12 + sul2 | >32 | 3 | ||||||||
| dfrA12 + sul3 | >32 | 2 | ||||||||
| dfrA12 + sul2 + sul3 | >32 | 1 | 5 | 2 | ||||||
| dfrA12 + sul1 + sul2 + sul3 | >32 | 1 | 1 | |||||||
| dfrA14 + sul1 | >32 | 1 | 5 | |||||||
| dfrA14 + sul2 | >32 | 4 | ||||||||
| dfrA14 + sul1 + sul2 | >32 | 1 | ||||||||
| dfrA14 + + sul3 | >32 | 1 | 2 | 1 | ||||||
| dfrA12 + dfrA5 + sul1 + sul2 + sul3 | >32 | 2 | ||||||||
| AZM (ECOFF 16.0) | ||||||||||
| No genes | 4–16 | 6 | 15 | 5 | 8 | 4 | ||||
| mph(A) | 64–128 | 4 | 3 | 3 | ||||||
| mef(B) | 32 | 1 | ||||||||
| TET (ECOFF 8.0) | ||||||||||
| No genes | 1–4 | 0 | 1 | 1 | ||||||
| tetM | 4–6 | 2 | ||||||||
| tetA | >256 | 4 | 2 | 3 | 10 | 3 | ||||
| tetB | >256 | 2 | 3 | |||||||
| tetA + tetB | >256 | 5 | 2 | 1 | ||||||
| tetA + tetM | >256 | 8 | 1 | 1 | ||||||
| CT (R>2) | ||||||||||
| No genes | 1–2 | 8 | 6 | 5 | 11 | 3 | ||||
| No genes | 4–8 | 1 | 3 | |||||||
| mcr 1.1 | 8–16 | 6 | 1 | |||||||
| mcr 3.1 | 4–8 | 2 | 2 | |||||||
| mcr 3.5 | 8 | 1 | ||||||||
| MIC | Susceptible isolates | |||||||||
| MIC | Susceptible isolates (according clinical breakpoint or wild–type) | |||||||||
| MIC * | Non–wild type isolate. CTX ECOFF 0.5 mg/L | |||||||||
| MIC | Susceptible isolates. increased exposure | |||||||||
| MIC | Resistant isolates | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starkova, D.; Egorova, S.; Suzhaeva, L.; Nguyen, T.Q.; Kaftyreva, L.; Makarova, M.; Zhamborova, S.; Polev, D.; Saitova, A.; Nguyen, V.H.; et al. Antimicrobial Resistance and Phylogenetic Analysis of Multidrug-Resistant Non-Typhoidal Salmonella Isolates from Different Sources in Southern Vietnam. Antibiotics 2025, 14, 489. https://doi.org/10.3390/antibiotics14050489
Starkova D, Egorova S, Suzhaeva L, Nguyen TQ, Kaftyreva L, Makarova M, Zhamborova S, Polev D, Saitova A, Nguyen VH, et al. Antimicrobial Resistance and Phylogenetic Analysis of Multidrug-Resistant Non-Typhoidal Salmonella Isolates from Different Sources in Southern Vietnam. Antibiotics. 2025; 14(5):489. https://doi.org/10.3390/antibiotics14050489
Chicago/Turabian StyleStarkova, Daria, Svetlana Egorova, Ludmila Suzhaeva, Truong Quang Nguyen, Lidiia Kaftyreva, Maria Makarova, Samida Zhamborova, Dmitrii Polev, Alina Saitova, Vu Hoang Nguyen, and et al. 2025. "Antimicrobial Resistance and Phylogenetic Analysis of Multidrug-Resistant Non-Typhoidal Salmonella Isolates from Different Sources in Southern Vietnam" Antibiotics 14, no. 5: 489. https://doi.org/10.3390/antibiotics14050489
APA StyleStarkova, D., Egorova, S., Suzhaeva, L., Nguyen, T. Q., Kaftyreva, L., Makarova, M., Zhamborova, S., Polev, D., Saitova, A., Nguyen, V. H., Vo, T. K., & Nguyen, L. T. (2025). Antimicrobial Resistance and Phylogenetic Analysis of Multidrug-Resistant Non-Typhoidal Salmonella Isolates from Different Sources in Southern Vietnam. Antibiotics, 14(5), 489. https://doi.org/10.3390/antibiotics14050489