
Identification of Antimicrobial-Resistant Zoonotic Bacteria in Swine Production: Implications from the One Health Perspective

 ,
,  , , , , , , ,
, , , , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
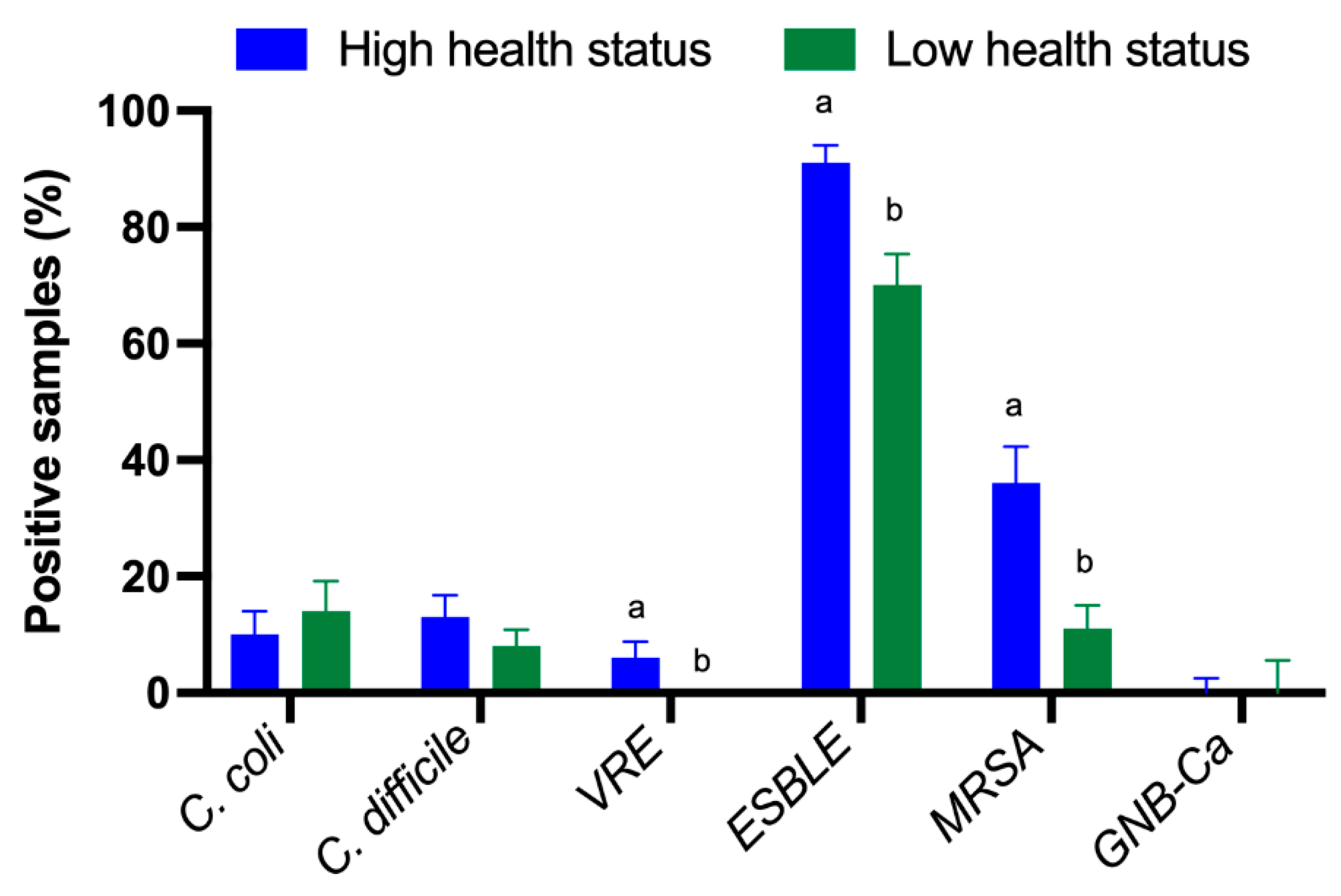
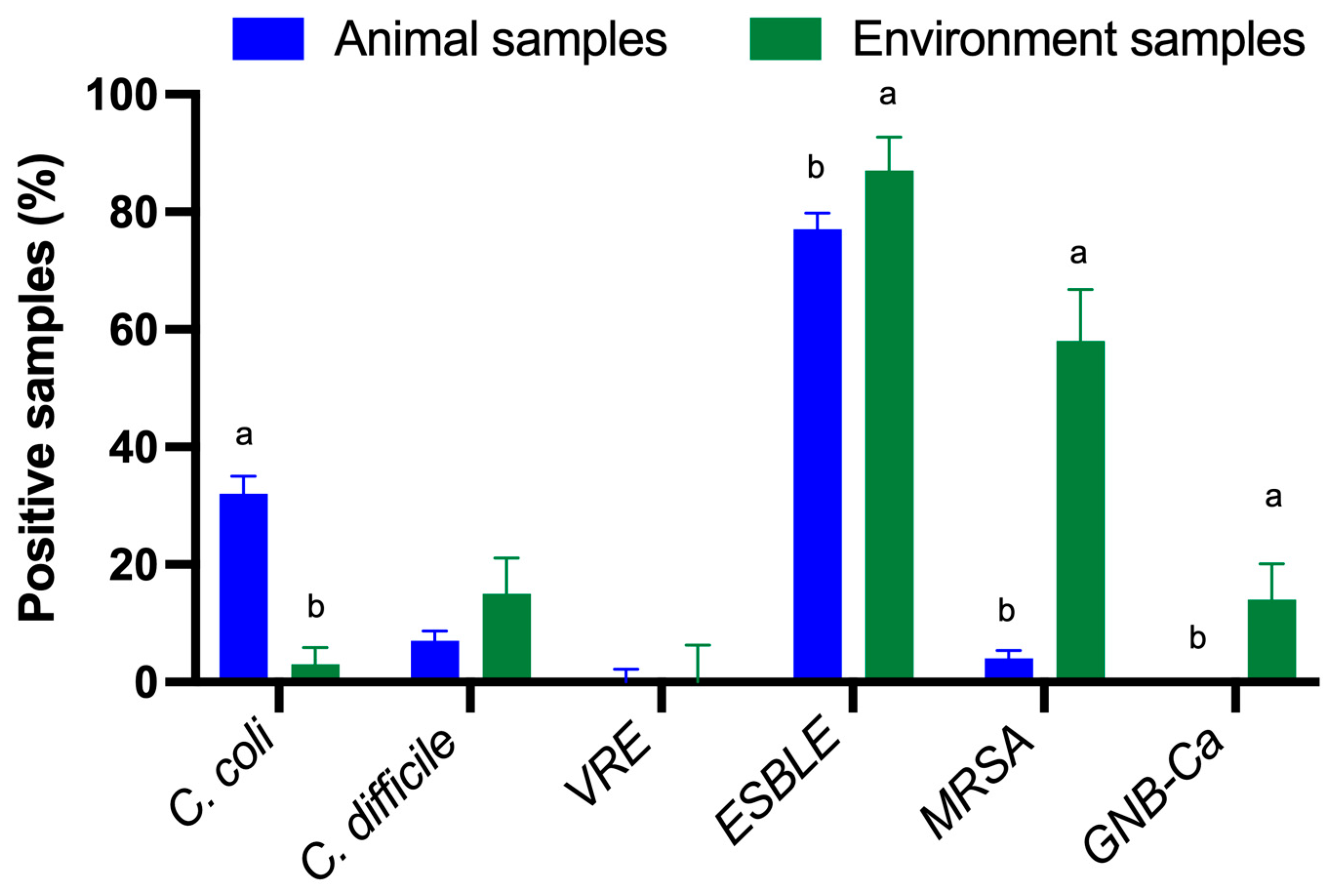
2. Results
Detection of Multidrug Resistant Bacteria and Enteric Pathogens
3. Discussion
4. Materials and Methods
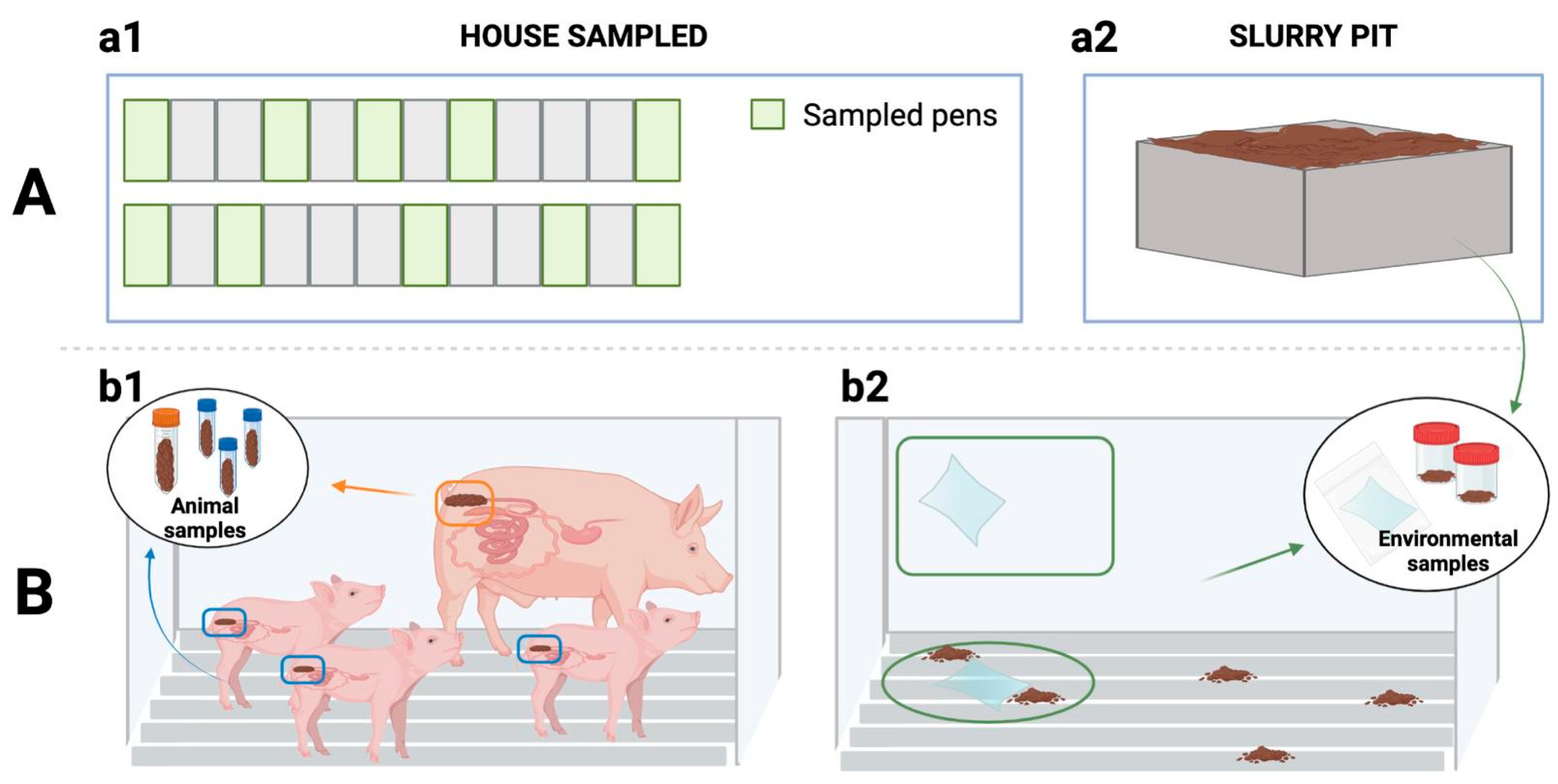
4.1. Study Sample
4.2. Selection of High and Low Sanitary Status Farms to Define “Healthy” and “Unhealthy” Microbiomes
4.3. Sampling
4.3.1. Environmental Sampling
4.3.2. Animal Sampling
4.4. Multidrug Resistant Bacteria and Enteric Pathogens Screening
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- OECD. Stemming the Superbug Tide; OECD: Paris, France, 2018. [Google Scholar]
- Gómez-García, M.; Sol, C.; De Nova, P.J.G.; Puyalto, M.; Mesas, L.; Puente, H.; Mencía-Ares, Ó.; Miranda, R.; Argüello, H.; Rubio, P.; et al. Antimicrobial Activity of a Selection of Organic Acids, Their Salts and Essential Oils against Swine Enteropathogenic Bacteria. Porc. Health Manag. 2019, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; CDC: Atlanta, GA, USA, 2013. [Google Scholar]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the Mechanisms and Drivers of Antimicrobial Resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Mencía-Ares, O.; Argüello, H.; Puente, H.; Gómez-García, M.; Manzanilla, E.G.; Álvarez-Ordóñez, A.; Carvajal, A.; Rubio, P. Antimicrobial Resistance in Commensal Escherichia coli and Enterococcus spp. Is Influenced by Production System, Antimicrobial Use, and Biosecurity Measures on Spanish Pig Farms. Porc. Health Manag. 2021, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Mencía-Ares, O.; Argüello, H.; Puente, H.; Gómez-García, M.; Álvarez-Ordóñez, A.; Manzanilla, E.G.; Carvajal, A.; Rubio, P. Effect of Antimicrobial Use and Production System on Campylobacter spp., Staphylococcus spp. and Salmonella spp. Resistance in Spanish Swine: A Cross-Sectional Study. Zoonoses Public Health 2021, 68, 54–66. [Google Scholar] [CrossRef]
- Plan Nacional Resistencia Antibióticos. Primer Informe Programa Reduce Colistina; PRAN: Madrid, Spain, 2017. [Google Scholar]
- Wu, Z. Approach Choices for Antimicrobial Use Reduction in European Food Animal Production. EuroChoices 2024, 23, 36–42. [Google Scholar] [CrossRef]
- De Paula Dias, C.; Pereira, A.R.; de Oliveira Paranhos, A.G.; Rodrigues, M.V.D.; de Lima, W.G.; de Aquino, S.F.; de Queiroz Silva, S. Spread of Multidrug-Resistant Bacteria and Antibiotic Resistance Genes in a Swine Wastewater Treatment Plant. Int. J. Environ. Res. 2024, 18, 2. [Google Scholar] [CrossRef]
- The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2021–2022. EFSA J. 2024, 22, e07867. [CrossRef]
- Castro, J.; Barros, M.M.; Araújo, D.; Campos, A.M.; Oliveira, R.; Silva, S.; Almeida, C. Swine Enteric Colibacillosis: Current Treatment Avenues and Future Directions. Front. Vet. Sci. 2022, 9, 981207. [Google Scholar] [CrossRef]
- Report from the Task Force on Zoonoses Data Collection Including Guidance for Harmonized Monitoring and Reporting of Antimicrobial Resistance in Commensal Escherichia coli and Enterococcus Spp. from Food Animals. EFSA J. 2008, 6, 141r. [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef]
- Anjum, M.F.; Marco-Jimenez, F.; Duncan, D.; Marín, C.; Smith, R.P.; Evans, S.J. Livestock-Associated Methicillin-Resistant Staphylococcus aureus From Animals and Animal Products in the UK. Front. Microbiol. 2019, 10, 442883. [Google Scholar] [CrossRef] [PubMed]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Programa REDUCE Porcino|PRAN. Available online: https://resistenciaantibioticos.es/es/lineas-de-accion/control/programas-de-reduccion-en-sanidad-animal/programa-reduce-porcino (accessed on 30 April 2024).
- Regulation—2019/6—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2019/6/oj (accessed on 30 April 2024).
- European Medicines Agency. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2022—Trends from 2010 to 2022—Thirteenth ESVAC Report; Publications Office of the European Union: Luxembourg, 2023. [Google Scholar] [CrossRef]
- Implementing Decision—2020/1729—EN—EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/en/TXT/?uri=CELEX%3A32020D1729 (accessed on 30 April 2024).
- Plan Nacional Resistencia Antibióticos. Plan Estratégico y de Acción para Reducir el Riesgo de Selección y Diseminación de la Resistencia a los Anti Bióti cos AEMPS; Agencia Española De Medicamentos Y Productos Sanitarios: Madrid, Spain, 2014. [Google Scholar]
- Cobo-Díaz, J.F.; Alvarez-Molina, A.; Alexa, E.A.; Walsh, C.J.; Mencía-Ares, O.; Puente-Gómez, P.; Likotrafiti, E.; Fernández-Gómez, P.; Prieto, B.; Crispie, F.; et al. Microbial Colonization and Resistome Dynamics in Food Processing Environments of a Newly Opened Pork Cutting Industry during 1.5 Years of Activity. Microbiome 2021, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Marin, C.; Cerdà-Cuéllar, M.; González-Bodi, S.; Lorenzo-Rebenaque, L.; Vega, S. Research Note: Persistent Salmonella Problems in Slaughterhouses Related to Clones Linked to Poultry Companies. Poult. Sci. 2022, 101, 101968. [Google Scholar] [CrossRef] [PubMed]
- Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef]
- GLASS|Global Antimicrobial Resistance Surveillance System (GLASS). Available online: https://www.who.int/initiatives/glass (accessed on 11 October 2019).
- O’Neill, L.; Manzanilla, E.G.; Ekhlas, D.; Leonard, F.C. Antimicrobial Resistance in Commensal Escherichia Coli of the Porcine Gastrointestinal Tract. Antibiotics 2023, 12, 1616. [Google Scholar] [CrossRef]
- Tseng, C.H.; Liu, C.W.; Liu, P.Y. Extended-Spectrum β-Lactamases (ESBL) Producing Bacteria in Animals. Antibiotics 2023, 12, 661. [Google Scholar] [CrossRef]
- Gelalcha, B.D.; Kerro Dego, O. Extended-Spectrum Beta-Lactamases Producing Enterobacteriaceae in the USA Dairy Cattle Farms and Implications for Public Health. Antibiotics 2022, 11, 1313. [Google Scholar] [CrossRef]
- Marin, C.; Lorenzo-Rebenaque, L.; Moreno-Moliner, J.; Sevilla-Navarro, S.; Montero, E.; Chinillac, M.C.; Jordá, J.; Vega, S. Multidrug-Resistant Campylobacer Jejuni on Swine Processing at a Slaughterhouse in Eastern Spain. Animals 2021, 11, 1339. [Google Scholar] [CrossRef]
- Marin, C.; Chinillac, M.C.; Cerdà-Cuéllar, M.; Montoro-Dasi, L.; Sevilla-Navarro, S.; Ayats, T.; Marco-Jimenez, F.; Vega, S. Contamination of Pig Carcass with Salmonella enterica Serovar Typhimurium Monophasic Variant 1,4[5],12:I:- Originates Mainly in Live Animals. Sci. Total Environ. 2020, 703, 134609. [Google Scholar] [CrossRef]
- Sevilla, E.; Marín, C.; Delgado-Blas, J.F.; González-Zorn, B.; Vega, S.; Kuijper, E.; Bolea, R.; Mainar-Jaime, R.C. Wild Griffon Vultures (Gyps fulvus) Fed at Supplementary Feeding Stations: Potential Carriers of Pig Pathogens and Pig-Derived Antimicrobial Resistance? Transbound. Emerg. Dis. 2020, 67, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lee, S.A.; Xue, J.; Riordan, S.M.; Zhang, L. Global Epidemiology of Campylobacteriosis and the Impact of COVID-19. Front. Cell. Infect. Microbiol. 2022, 12, 979055. [Google Scholar] [CrossRef] [PubMed]
- Spronk, T.; Green, A.L.; Vuolo, M.; Ruesch, L.; Edler, R.; Haley, C.; Scaria, J.; Hennings, J.; Dee, S.; Shivley, C.B. Antimicrobial Use and Antimicrobial Resistance Monitoring in Pig Production in the United States of America. OIE Rev. Sci. Tech. 2023, 42, 52–64. [Google Scholar] [CrossRef]
- Pokhrel, D.; Thames, H.T.; Zhang, L.; Dinh, T.T.N.; Schilling, W.; White, S.B.; Ramachandran, R.; Theradiyil Sukumaran, A. Roles of Aerotolerance, Biofilm Formation, and Viable but Non-Culturable State in the Survival of Campylobacter jejuni in Poultry Processing Environments. Microorganisms 2022, 10, 2165. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.S.; Hung, Y.P.; Lee, J.C.; Syue, L.S.; Hsueh, P.R.; Ko, W.C. Clostridioides difficile Infection: An Emerging Zoonosis? Expert Rev. Anti. Infect. Ther. 2021, 19, 1543–1552. [Google Scholar] [CrossRef]
- Uzal, F.A.; Navarro, M.A.; Asin, J.; Boix, O.; Ballarà-Rodriguez, I.; Gibert, X. Clostridial Diarrheas in Piglets: A Review. Vet. Microbiol. 2023, 280, 109691. [Google Scholar] [CrossRef]
- Redding, L.; Huang, E.; Ryave, J.; Webb, T.; Barnhart, D.; Baker, L.; Bender, J.; Kristula, M.; Kelly, D. Clostridioides difficile on Dairy Farms and Potential Risk to Dairy Farm Workers. Anaerobe 2021, 69, 102353. [Google Scholar] [CrossRef]
- Hain-Saunders, N.M.R.; Knight, D.R.; Bruce, M.; Riley, T.V. Clostridioides difficile Infection and One Health: An Equine Perspective. Environ. Microbiol. 2022, 24, 985–997. [Google Scholar] [CrossRef]
- Licciardi, C.; Primavilla, S.; Roila, R.; Lupattelli, A.; Farneti, S.; Blasi, G.; Petruzzelli, A.; Drigo, I.; Di Raimo Marrocchi, E. Prevalence, Molecular Characterization and Antimicrobial Susceptibility of Clostridioides Difficile Isolated from Pig Carcasses and Pork Products in Central Italy. Int. J. Environ. Res. Public Health 2021, 18, 11368. [Google Scholar] [CrossRef]
- Usui, M. One Health Approach to Clostridioides difficile in Japan. J. Infect. Chemother. 2020, 26, 643–650. [Google Scholar] [CrossRef]
- Alves, F.; Nunes, A.; Castro, R.; Sequeira, A.; Moreira, O.; Matias, R.; Rodrigues, J.C.; Silveira, L.; Gomes, J.P.; Oleastro, M. Assessment of the Transmission Dynamics of Clostridioides difficile in a Farm Environment Reveals the Presence of a New Toxigenic Strain Connected to Swine Production. Front. Microbiol. 2022, 13, 858310. [Google Scholar] [CrossRef]
- Spigaglia, P.; Barbanti, F.; Faccini, S.; Vescovi, M.; Criscuolo, E.M.; Ceruti, R.; Gaspano, C.; Rosignoli, C. Clostridioides difficile in Pigs and Dairy Cattle in Northern Italy: Prevalence, Characterization and Comparison between Animal and Human Strains. Microorganisms 2023, 11, 1738. [Google Scholar] [CrossRef] [PubMed]
- Hassoun-Kheir, N.; Guedes, M.; Ngo Nsoga, M.T.; Argante, L.; Arieti, F.; Gladstone, B.P.; Kingston, R.; Naylor, N.R.; Pezzani, M.D.; Pouwels, K.B.; et al. A Systematic Review on the Excess Health Risk of Antibiotic-Resistant Bloodstream Infections for Six Key Pathogens in Europe. Clin. Microbiol. Infect. 2023, 30, S14–S25. [Google Scholar] [CrossRef] [PubMed]
- Kasela, M.; Ossowski, M.; Dzikoń, E.; Ignatiuk, K.; Wlazło, Ł.; Malm, A. The Epidemiology of Animal-Associated Methicillin-Resistant Staphylococcus Aureus. Antibiotics 2023, 12, 1079. [Google Scholar] [CrossRef] [PubMed]
- Khairullah, A.R.; Kurniawan, S.C.; Effendi, M.H.; Sudjarwo, S.A.; Ramandinianto, S.C.; Widodo, A.; Riwu, K.H.P.; Silaen, O.S.M.; Rehman, S. A Review of New Emerging Livestock-Associated Methicillin-Resistant Staphylococcus aureus from Pig Farms. Vet. World 2023, 16, 46–58. [Google Scholar] [CrossRef]
- Silva, V.; Monteiro, A.; Pereira, J.E.; Maltez, L.; Igrejas, G.; Poeta, P. MRSA in Humans, Pets and Livestock in Portugal: Where We Came from and Where We Are Going. Pathogens 2022, 11, 1110. [Google Scholar] [CrossRef]
- Eichel, V.M.; Last, K.; Brühwasser, C.; von Baum, H.; Dettenkofer, M.; Götting, T.; Grundmann, H.; Güldenhöven, H.; Liese, J.; Martin, M.; et al. Epidemiology and Outcomes of Vancomycin-Resistant Enterococcus Infections: A Systematic Review and Meta-Analysis. J. Hosp. Infect. 2023, 141, 119–128. [Google Scholar] [CrossRef]
- Sugai, M.; Yuasa, A.; Miller, R.L.; Vasilopoulos, V.; Kurosu, H.; Taie, A.; Gordon, J.P.; Matsumoto, T. An Economic Evaluation Estimating the Clinical and Economic Burden of Increased Vancomycin-Resistant Enterococcus faecium Infection Incidence in Japan. Infect. Dis. Ther. 2023, 12, 1695–1713. [Google Scholar] [CrossRef]
- Liu, Z.L.; Chen, Y.J.; Meng, Q.L.; Zhang, X.; Wang, X.L. Progress in the Application of Enterococcus faecium in Animal Husbandry. Front. Cell. Infect. Microbiol. 2023, 13, 1168189. [Google Scholar] [CrossRef]
- Grudlewska-Buda, K.; Skowron, K.; Bauza-Kaszewska, J.; Budzyńska, A.; Wiktorczyk-Kapischke, N.; Wilk, M.; Wujak, M.; Paluszak, Z. Assessment of Antibiotic Resistance and Biofilm Formation of Enterococcus Species Isolated from Different Pig Farm Environments in Poland. BMC Microbiol. 2023, 23, 89. [Google Scholar] [CrossRef]
- Marques, J.M.; Coelho, M.; Santana, A.R.; Pinto, D.; Semedo-Lemsaddek, T. Dissemination of Enterococcal Genetic Lineages: A One Health Perspective. Antibiotics 2023, 12, 1140. [Google Scholar] [CrossRef] [PubMed]
- Tamai, S.; Suzuki, Y. Diversity of Fecal Indicator Enterococci among Different Hosts: Importance to Water Contamination Source Tracking. Microorganisms 2023, 11, 2981. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xiang, Q.; Ma, J.; Zhang, P.; Li, K.; Wu, K.; Su, M.; Li, R.; Hurley, D.; Bai, L.; et al. Characterization of Carbapenem-Resistant Enterobacteriaceae Cultured From Retail Meat Products, Patients, and Porcine Excrement in China. Front. Microbiol. 2021, 12, 743468. [Google Scholar] [CrossRef] [PubMed]
- Lysitsas, M.; Triantafillou, E.; Chatzipanagiotidou, I.; Antoniou, K.; Valiakos, G. Antimicrobial Susceptibility Profiles of Acinetobacter baumannii Strains, Isolated from Clinical Cases of Companion Animals in Greece. Vet. Sci. 2023, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Mairi, A.; Pantel, A.; Sotto, A.; Lavigne, J.P.; Touati, A. OXA-48-like Carbapenemases Producing Enterobacteriaceae in Different Niches. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 37, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, N.H.; Jang, K.M.; Jin, H.; Shin, K.; Jeong, B.C.; Kim, D.W.; Lee, S.H. Prioritization of Critical Factors for Surveillance of the Dissemination of Antibiotic Resistance in Pseudomonas aeruginosa: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 15209. [Google Scholar] [CrossRef]
- Tedersoo, T.; Roasto, M.; Mäesaar, M.; Fredriksson-Ahomaa, M.; Meremäe, K. Antimicrobial Resistance of Campylobacter coli Isolated from Caecal Samples of Fattening Pigs at Slaughter. Microorganisms 2023, 11, 1540. [Google Scholar] [CrossRef]
- Costa, M.M.; Cardo, M.; Ruano, Z.; Alho, A.M.; Dinis-Teixeira, J.; Aguiar, P.; Leite, A. Effectiveness of Antimicrobial Interventions Directed at Tackling Antimicrobial Resistance in Animal Production: A Systematic Review and Meta-Analysis. Prev. Vet. Med. 2023, 218, 106002. [Google Scholar] [CrossRef]
- Latifi, F.; Pakzad, R.; Asadollahi, P.; Hematian, A.; Pakzad, I. Worldwide Prevalence of Colistin Resistance among Enterobacteriaceae: A Systematic Review and Meta-Analysis. Clin. Lab. 2023, 69, 657–669. [Google Scholar] [CrossRef]
- Bastidas-Caldes, C.; de Waard, J.H.; Salgado, M.S.; Villacís, M.J.; Coral-Almeida, M.; Yamamoto, Y.; Calvopiña, M. Worldwide Prevalence of Mcr-Mediated Colistin-Resistance Escherichia Coli in Isolates of Clinical Samples, Healthy Humans, and Livestock—A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 659. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet. Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Madec, J.Y.; Laxminarayan, R. Colistin: From the Shadows to a One Health Approach for Addressing Antimicrobial Resistance. Int. J. Antimicrob. Agents 2023, 61, 106713. [Google Scholar] [CrossRef] [PubMed]
- Tozzo, P.; Delicati, A.; Caenazzo, L. Human Microbiome and Microbiota Identification for Preventing and Controlling Healthcare-Associated Infections: A Systematic Review. Front. Public Health 2022, 10, 989496. [Google Scholar] [CrossRef] [PubMed]
- Collineau, L.; Bourely, C.; Rousset, L.; Berger-Carbonne, A.; Ploy, M.C.; Pulcini, C.; Colomb-Cotinat, M. Towards One Health Surveillance of Antibiotic Resistance: Characterisation and Mapping of Existing Programmes in Humans, Animals, Food and the Environment in France, 2021. Eurosurveillance 2023, 28, 2200804. [Google Scholar] [CrossRef] [PubMed]
- Gehring, R.; Mochel, J.P.; Schmerold, I. Understanding the Background and Clinical Significance of the WHO, WOAH, and EMA Classifications of Antimicrobials to Mitigate Antimicrobial Resistance. Front. Vet. Sci. 2023, 10, 1153048. [Google Scholar] [CrossRef]
- De Koster, S.; Xavier, B.B.; Lammens, C.; Selva, N.P.; van Kleef-Van Koeveringe, S.; Coenen, S.; Glupczynski, Y.; Leroux-Roels, I.; Dhaeze, W.; Hoebe, C.J.P.A.; et al. One Health Surveillance of Colistin-Resistant Enterobacterales in Belgium and the Netherlands between 2017 and 2019. PLoS ONE 2024, 19, e0298096. [Google Scholar] [CrossRef]
- Abreu, R.; Rodríguez-álvarez, C.; Castro-Hernandez, B.; Lecuona-Fernández, M.; González, J.C.; Rodríguez-Novo, Y.; Arias Rodríguez, M. de los A. Prevalence and Characterisation of Multiresistant Bacterial Strains Isolated in Pigs from the Island of Tenerife. Vet. Sci. 2022, 9, 269. [Google Scholar] [CrossRef]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12, 487. [Google Scholar] [CrossRef]
- Li, T.; Wang, Z.; Guo, J.; de la Fuente-Nunez, C.; Wang, J.; Han, B.; Tao, H.; Liu, J.; Wang, X. Bacterial Resistance to Antibacterial Agents: Mechanisms, Control Strategies, and Implications for Global Health. Sci. Total Environ. 2023, 860, 160461. [Google Scholar] [CrossRef]
- Akram, F.; Imtiaz, M.; Haq, I. ul Emergent Crisis of Antibiotic Resistance: A Silent Pandemic Threat to 21st Century. Microb. Pathog. 2023, 174, 105923. [Google Scholar] [CrossRef]
- Chen, L.; Kumar, S.; Wu, H. A Review of Current Antibiotic Resistance and Promising Antibiotics with Novel Modes of Action to Combat Antibiotic Resistance. Arch. Microbiol. 2023, 205, 356. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventero, M.P.; Marin, C.; Migura-Garcia, L.; Tort-Miro, C.; Giler, N.; Gomez, I.; Escribano, I.; Marco-Fuertes, A.; Montoro-Dasi, L.; Lorenzo-Rebenaque, L.; et al. Identification of Antimicrobial-Resistant Zoonotic Bacteria in Swine Production: Implications from the One Health Perspective. Antibiotics 2024, 13, 883. https://doi.org/10.3390/antibiotics13090883
Ventero MP, Marin C, Migura-Garcia L, Tort-Miro C, Giler N, Gomez I, Escribano I, Marco-Fuertes A, Montoro-Dasi L, Lorenzo-Rebenaque L, et al. Identification of Antimicrobial-Resistant Zoonotic Bacteria in Swine Production: Implications from the One Health Perspective. Antibiotics. 2024; 13(9):883. https://doi.org/10.3390/antibiotics13090883
Chicago/Turabian StyleVentero, Maria Paz, Clara Marin, Lourdes Migura-Garcia, Carla Tort-Miro, Noemi Giler, Inmaculada Gomez, Isabel Escribano, Ana Marco-Fuertes, Laura Montoro-Dasi, Laura Lorenzo-Rebenaque, and et al. 2024. "Identification of Antimicrobial-Resistant Zoonotic Bacteria in Swine Production: Implications from the One Health Perspective" Antibiotics 13, no. 9: 883. https://doi.org/10.3390/antibiotics13090883
APA StyleVentero, M. P., Marin, C., Migura-Garcia, L., Tort-Miro, C., Giler, N., Gomez, I., Escribano, I., Marco-Fuertes, A., Montoro-Dasi, L., Lorenzo-Rebenaque, L., Vega, S., Pérez-Gracia, M. T., & Rodríguez, J. C. (2024). Identification of Antimicrobial-Resistant Zoonotic Bacteria in Swine Production: Implications from the One Health Perspective. Antibiotics, 13(9), 883. https://doi.org/10.3390/antibiotics13090883