
Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa

 ,
,
Abstract
1. Introduction
2. Results
2.1. Purification of Cineole and Selection of Homeopathic Medicines
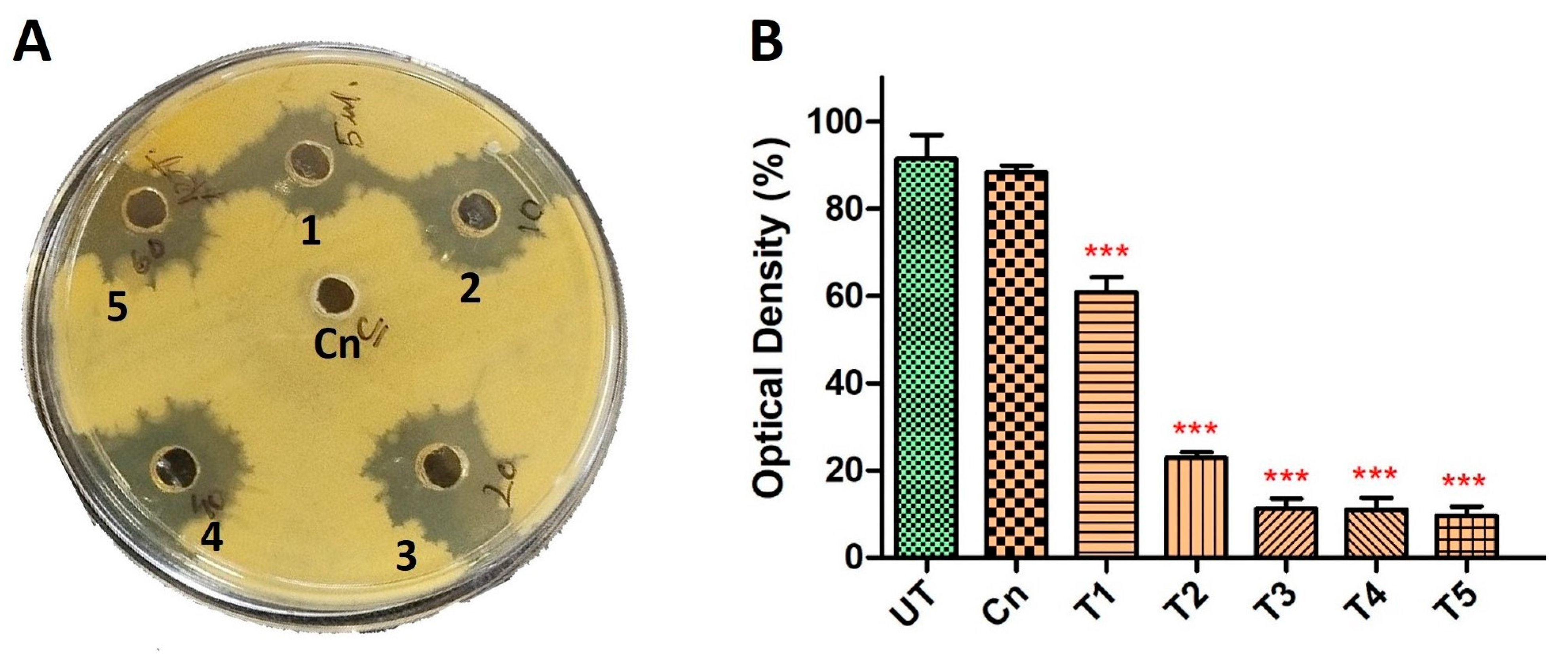
2.2. Screening of Homeopathic Medicines against Biofilm-Forming Drug-Resistant P. aeruginosa in Combination with Cineole
2.3. Hypericum Perforatum Showed Efficient Activity against Both Biofilm and Planktonic Cells of Drug-Resistant P. aeruginosa
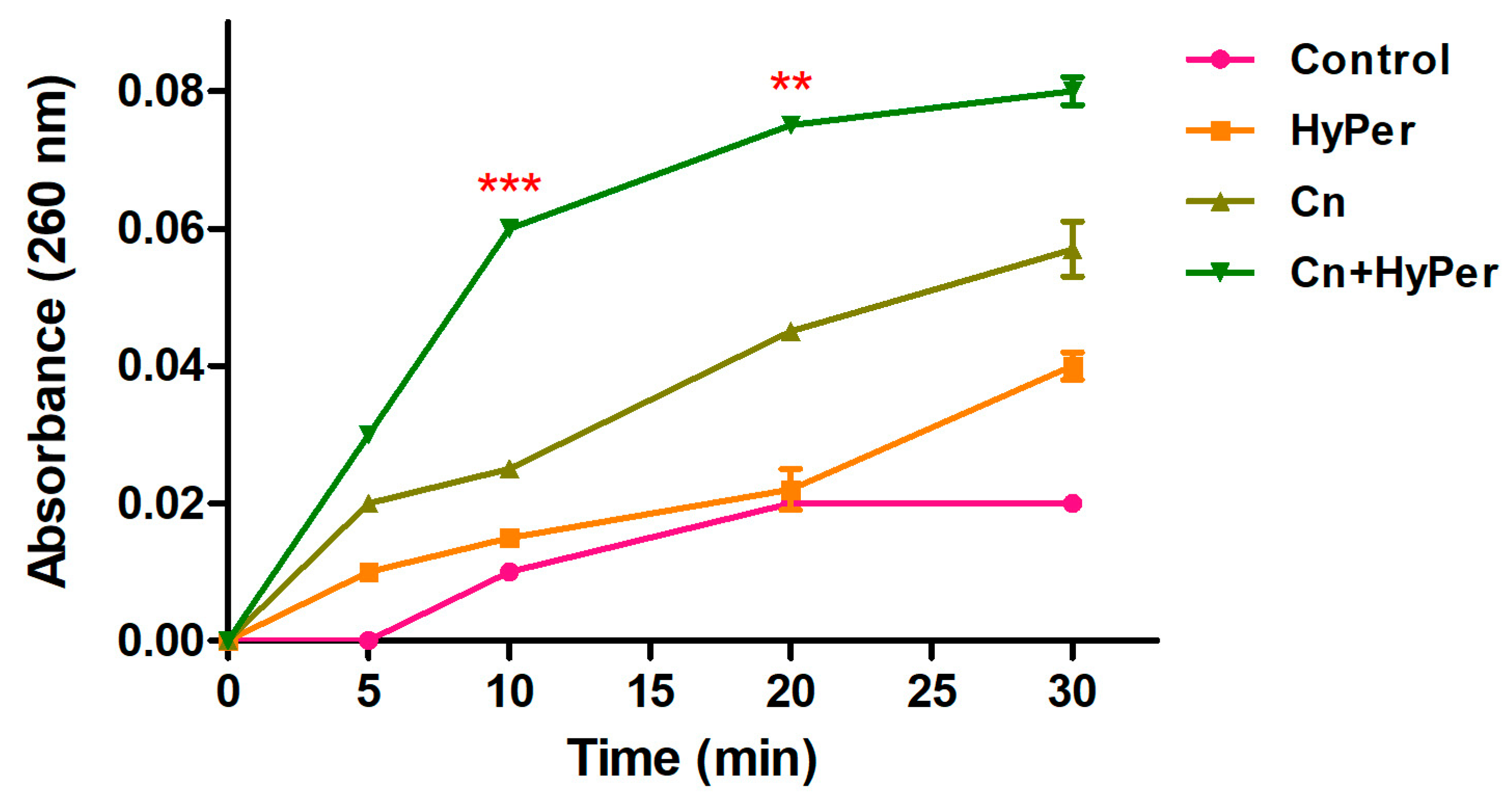
2.4. Synergistic Killing Mechanism of Hypericum Perforatum with Cineole against P. aeruginosa
2.4.1. Disruption of Membrane Zeta Potential
2.4.2. Disruption of Outer Membrane Permeability
2.4.3. DNA Leakage
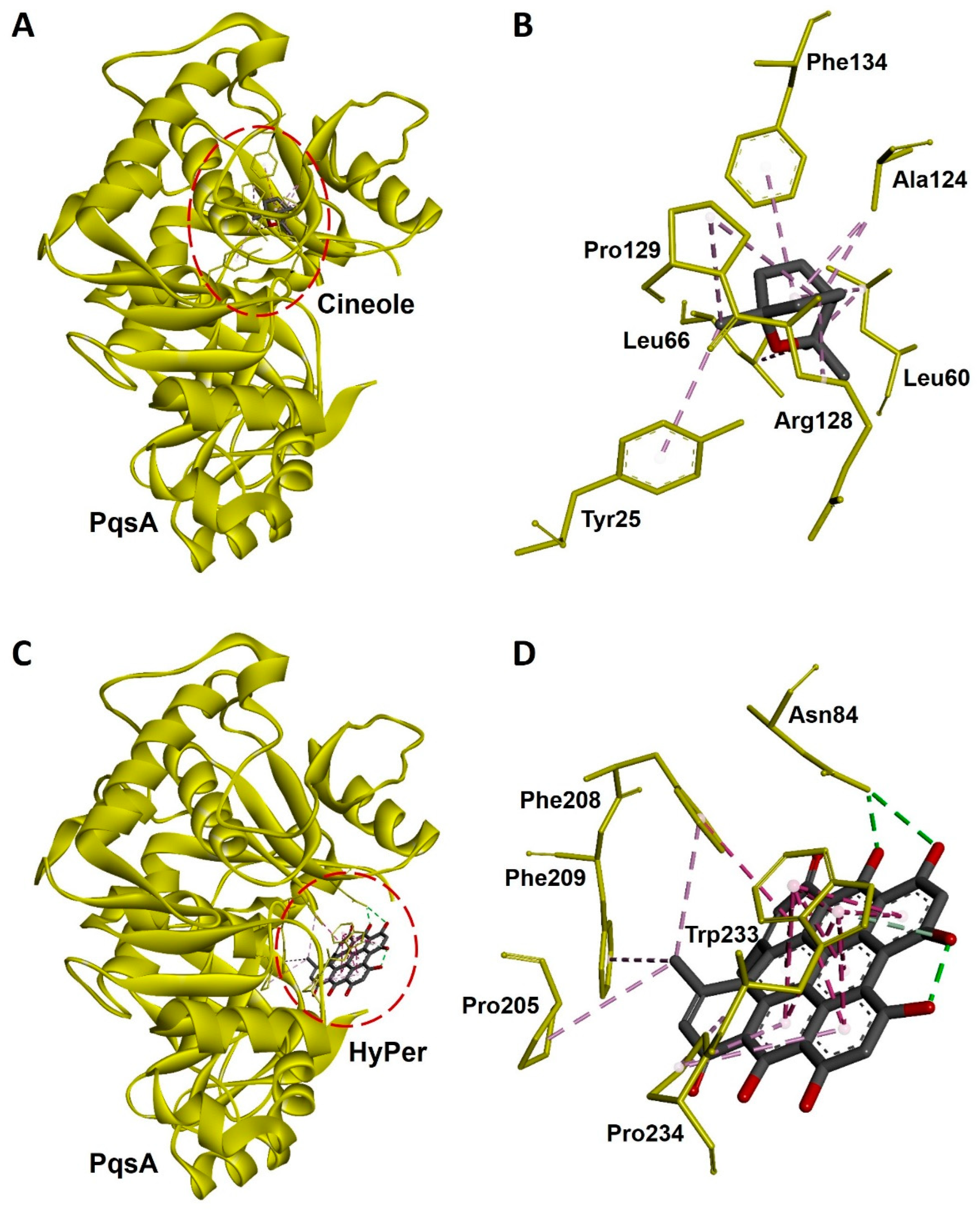
2.5. Molecular Docking Revealed a Strong Interaction of Hypericin with PqsA of P. aeruginosa
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Plant Materials
5.2. Homeopathic Medicines and Cineole
5.3. Bacterial Strain and Media
5.4. Thin Layer Chromatography
5.5. High-Performance Liquid Chromatography
5.6. Gas Chromatography–Mass Spectrometry Analysis
5.7. Agar Well Diffusion Assay
5.8. MIC Determination
5.9. Biofilm Eradication Assay
5.10. Microscopy
5.11. Determination of Zeta Potential
5.12. Outer Membrane Permeability Assay
5.13. DNA Leakage Assay
5.14. Protein Retrieval and Preparation
5.15. Ligand Preparation
5.16. Molecular Docking Analysis
5.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Trouillet, J.L.; Chastre, J.; Vuagnat, A.; Joly-Guillou, M.L.; Combaux, D.; Dombret, M.C.; Gibert, C. Ventilator-associated pneumonia caused by potentially drug-resistant bacteria. Am. J. Respir. Crit. Care Med. 1998, 157, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Guitor, A.K.; Wright, G.D. Antimicrobial Resistance and Respiratory Infections. Chest 2018, 154, 1202–1212. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Kunz Coyne, A.J.; El Ghali, A.; Holger, D.; Rebold, N.; Rybak, M.J. Therapeutic Strategies for Emerging Multidrug-Resistant Pseudomonas aeruginosa. Infect. Dis. Ther. 2022, 11, 661–682. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Mandal, S.M. Gut-Antimicrobial Peptides: Synergistic Co-Evolution with Antibiotics to Combat Multi-Antibiotic Resistance. Antibiotics 2023, 12, 1732. [Google Scholar] [CrossRef]
- Mandal, S.M.; Dias, R.O.; Franco, O.L. Phenolic Compounds in Antimicrobial Therapy. J. Med. Food 2017, 20, 1031–1038. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Khameneh, B.; Eskin, N.A.M.; Iranshahy, M.; Fazly Bazzaz, B.S. Phytochemicals: A promising weapon in the arsenal against antibiotic-resistant bacteria. Antibiotics 2021, 10, 1044. [Google Scholar] [CrossRef]
- Hawke, K.; van Driel, M.L.; Buffington, B.J.; McGuire, T.M.; King, D. Homeopathic medicinal products for preventing and treating acute respiratory tract infections in children. Cochrane Database Syst. Rev. 2018, 9, CD005974. [Google Scholar]
- Steinsbekk, A.; Bentzen, N.; Fønnebø, V.; Lewith, G. Self treatment with one of three self selected, ultramolecular homeopathic medicines for the prevention of upper respiratory tract infections in children. A double-blind randomized placebo controlled trial. Br. J. Clin. Pharmacol. 2005, 59, 447–455. [Google Scholar] [CrossRef]
- Zanasi, A.; Mazzolini, M.; Tursi, F.; Morselli-Labate, A.M.; Paccapelo, A.; Lecchi, M. Homeopathic medicine for acute cough in upper respiratory tract infections and acute bronchitis: A randomized, double-blind, placebo-controlled trial. Pulm. Pharmacol. Ther. 2014, 27, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Moo, C.L.; Osman, M.A.; Yang, S.K.; Yap, W.S.; Ismail, S.; Lim, S.H.E.; Chong, C.M.; Lai, K.S. Antimicrobial activity and mode of action of 1,8-cineol against carbapenemase-producing Klebsiella pneumoniae. Sci. Rep. 2021, 11, 20824. [Google Scholar] [CrossRef] [PubMed]
- Hoch, C.C.; Petry, J.; Griesbaum, L.; Weiser, T.; Werner, K.; Ploch, M.; Verschoor, A.; Multhoff, G.; Bashiri Dezfouli, A.; Wollenberg, B. 1,8-cineole (eucalyptol): A versatile phytochemical with therapeutic applications across multiple diseases. Biomed. Pharmacother. 2023, 167, 115467. [Google Scholar] [CrossRef] [PubMed]
- Bononi, I.; Balatti, V.; Gaeta, S.; Tognon, M. Gram-negative bacterial lipopolysaccharide retention by a positively charged new-generation filter. Appl. Environ. Microbiol. 2008, 74, 6470–6472. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, L.; Wang, X.; Li, X.; Zhang, Y.; Yu, J.; Jiang, J.; You, X.; Xiong, Y.Q. Hypericin enhances β-lactam antibiotics activity by inhibiting sarA expression in methicillin-resistant Staphylococcus aureus. Acta Pharm. Sin. B 2019, 9, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Shaker, B.; Ahmad, S.; Thai, T.D.; Eyun, S.-I.; Na, D. Rational Drug Design for Pseudomonas aeruginosa PqsA Enzyme: An in silico Guided Study to Block Biofilm Formation. Front. Mol. Biosci. 2020, 7, 577316. [Google Scholar] [CrossRef] [PubMed]
- Maurice, N.M.; Bedi, B.; Sadikot, R.T. Pseudomonas aeruginosa biofilms: Host response and clinical implications in lung infections. Am. J. Respir. Cell Mol. Biol. 2018, 58, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef]
- Dale, J.L.; Nilson, J.L.; Barnes, A.M.T.; Dunny, G.M. Restructuring of Enterococcus faecalis biofilm architecture in response to antibiotic-induced stress. npj Biofilms Microbiomes 2017, 3, 15. [Google Scholar] [CrossRef]
- Almas, I.; Innocent, E.; Machumi, F.; Kisinza, W. Chemical composition of essential oils from Eucalyptus globulus and Eucalyptus maculata grown in Tanzania. Sci. Afr. 2021, 12, e00758. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and resistance of Pseudomonas aeruginosa biofilms to antimicrobial agents—How P. aeruginosa Can escape antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Fixsen, A. Homeopathy in the Age of Antimicrobial Resistance: Is It a Viable Treatment for Upper Respiratory Tract Infections? Homeopathy 2018, 107, 99–114. [Google Scholar] [CrossRef]
- Kurd, R.; Freed, Y.; Jarjoui, A.; Izbicki, G.; Levin, P.; Helvitz, Y.; Sherr, J.; Oberbaum, M. Homeopathic Treatment for COVID-19-Related Symptoms: A Case Series. Complement. Med. Res. 2022, 29, 83–88. [Google Scholar] [CrossRef]
- To, K.L.A.; Fok, Y.Y.Y. Homeopathic Clinical Features of 18 Patients in COVID-19 Outbreaks in Hong Kong. Homeopathy 2020, 109, 146–162. [Google Scholar] [CrossRef]
- Abbas, A.; Anwar, F.; Alqahtani, S.M.; Ahmad, N.; Al-Mijalli, S.H.; Shahid, M.; Iqbal, M. Hydro-Distilled and Supercritical Fluid Extraction of Eucalyptus camaldulensis Essential Oil: Characterization of Bioactives Along With Antioxidant, Antimicrobial and Antibiofilm Activities. Dose-Response 2022, 20, 15593258221125477. [Google Scholar] [CrossRef] [PubMed]
- Abd al-Raouf Ammar Semysim, E. Antibacterial Activity of Essential oils Isolated from Eucalyptus globulus Labill and Eugnia caryophyllata Thunbery (Family Myrtaceae): A comparative study. Al-Kufa Univ. J. Biol. 2018, 10, 32–39. [Google Scholar] [CrossRef]
- Chaudhry, V.; Baindara, P.; Pal, V.K.; Chawla, N.; Patil, P.B.; Korpole, S. Methylobacterium indicum sp. nov., a facultative methylotrophic bacterium isolated from rice seed. Syst. Appl. Microbiol. 2016, 39, 25–32. [Google Scholar] [CrossRef]
- Baindara, P.; Singh, N.; Ranjan, M.; Nallabelli, N.; Chaudhry, V.; Pathania, G.L.; Sharma, N.; Kumar, A.; Patil, P.B.; Korpole, S. Laterosporulin10: A novel defensin like class iid bacteriocin from Brevibacillus sp. strain SKDU10 with inhibitory activity against microbial pathogens. Microbiology 2016, 162, 1286–1299. [Google Scholar] [CrossRef]
- Chakraborty, S.; Baindara, P.; Mondal, S.K.; Roy, D.; Mandal, S.M. Synthesis of a tetralone derivative of ampicillin to control ampicillin-resistant Staphylococcus aureus. Biochem. Biophys. Res. Commun. 2024, 714, 149974. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G.; Petrelli, R.; Cappellacci, L.; Lupidi, G.; Sut, S.; Dall’Acqua, S.; Maggi, F. Exploring the Insecticidal Potential of Boldo (Peumus boldus) Essential Oil: Toxicity to Pests and Vectors and Non-target Impact on the Microcrustacean Daphnia magna. Molecules 2019, 24, 879. [Google Scholar] [CrossRef]
- Haney, E.F.; Trimble, M.J.; Cheng, J.T.; Vallé, Q.; Hancock, R.E.W. Critical assessment of methods to quantify biofilm growth and evaluate antibiofilm activity of host defence peptides. Biomolecules 2018, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Mai, C.W.; Lim, W.M.; Yap, W.S.; Lim, S.H.E.; Lai, K.S. Additivity vs. synergism: Investigation of the additive interaction of cinnamon bark oil and meropenem in combinatory therapy. Molecules 2017, 22, 1733. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yusoff, K.; Ajat, M.; Yap, W.S.; Lim, S.H.E.; Lai, K.S. Antimicrobial activity and mode of action of terpene linalyl anthranilate against carbapenemase-producing Klebsiella pneumoniae. J. Pharm. Anal. 2021, 11, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.S.X.; Krishnan, T.; Yiap, B.C.; Hu, C.P.; Chan, K.G.; Lim, S.H.E. Membrane disruption and anti-quorum sensing effects of synergistic interaction between Lavandula angustifolia (lavender oil) in combination with antibiotic against plasmid-conferred multi-drug-resistant Escherichia coli. J. Appl. Microbiol. 2014, 116, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Li, X.; Bi, W.; Jiang, C. Relationship between membrane damage, leakage of intracellular compounds, and inactivation of Escherichia coli treated by pressurized CO2. J. Basic Microbiol. 2014, 54, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Schmidtke, P.; Le Guilloux, V.; Maupetit, J.; Tufféry, P. fpocket: Online tools for protein ensemble pocket detection and tracking. Nucleic Acids Res. 2010, 38, W582-9. [Google Scholar] [CrossRef] [PubMed]
- Dinata, R.; Baindara, P. Laterosporulin25: A probiotically produced, novel defensin-like bacteriocin and its immunogenic properties. Int. Immunopharmacol. 2023, 121, 110500. [Google Scholar] [CrossRef]
- Yan, Y.; Tao, H.; He, J.; Huang, S.Y. The HDOCK server for integrated protein–protein docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Binding Energy (kcal/mol) | ||
|---|---|---|
| Cineole | Hypericin | |
| PqsA | −70.21 | −176.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, S.; Baindara, P.; Sharma, P.; Jose T, A.; V, K.; Manoharan, R.; Mandal, S.M. Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa. Antibiotics 2024, 13, 689. https://doi.org/10.3390/antibiotics13080689
Chakraborty S, Baindara P, Sharma P, Jose T A, V K, Manoharan R, Mandal SM. Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa. Antibiotics. 2024; 13(8):689. https://doi.org/10.3390/antibiotics13080689
Chicago/Turabian StyleChakraborty, Sourav, Piyush Baindara, Pralay Sharma, Austin Jose T, Kumaravel V, Raja Manoharan, and Santi M. Mandal. 2024. "Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa" Antibiotics 13, no. 8: 689. https://doi.org/10.3390/antibiotics13080689
APA StyleChakraborty, S., Baindara, P., Sharma, P., Jose T, A., V, K., Manoharan, R., & Mandal, S. M. (2024). Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa. Antibiotics, 13(8), 689. https://doi.org/10.3390/antibiotics13080689