Abstract
Gastric cancer (GC) still represents one of the leading causes of cancer-related mortality and is a major public health issue worldwide. Understanding the etiopathogenetic mechanisms behind GC development holds immense potential to revolutionize patients’ treatment and prognosis. Within the complex web of genetic predispositions and environmental factors, the connection between Helicobacter pylori (H. pylori) and gastric microbiota emerges as a focus of intense research investigation. According to the most recent hypotheses, H. pylori triggers inflammatory responses and molecular alterations in gastric mucosa, while non-Helicobacter microbiota modulates disease progression. In this review, we analyze the current state of the literature on the relationship between H. pylori and non-Helicobacter gastric microbiota in gastric carcinogenesis, highlighting the mechanisms by which microecological dysbiosis can contribute to the malignant transformation of the mucosa.
1. Introduction
Gastric cancer (GC) ranks as the fifth most common cancer and the third leading cause of cancer-related death [1]. Helicobacter pylori (H. pylori) is the most well-established risk factor for GC, being associated with an up to six-fold increased risk of non-cardia adenocarcinoma [2,3]. However, only 1–3% of the infected individuals progress to GC [4,5,6]. The genetic characteristics of both the bacterium [7,8,9,10] and the host [11,12,13], as well as environmental factors, such as diet and lifestyle, can impact H. pylori-associated disease outcomes [14,15]. However, none of these factors fully account for GC risk, and it is not yet possible to predict who will develop cancer.
Increasing evidence suggests a potential role of non-Helicobacter gastric microbiota as a significant contributor to GC development [16]. This hypothesis stems from the observation that, at a certain point, gastric carcinogenesis appears to occur independently of the presence of H. pylori. Indeed, H. pylori colonization decreases over time in individuals with precancerous gastric lesions and eventually disappears in the adenocarcinoma stage [17]. The hypo/achlorhydria of the gastric environment induced by the chronic H. pylori infection shifts the gastric microbiota composition toward microbes typically found in the oral cavity and lower bowel, which would not normally thrive in acidic environments. The altered microbiota may promote the malignant transformation of gastric mucosa by sustaining inflammation, increasing the production of carcinogenic N-nitroso compounds and numerous other pro-carcinogenic mechanisms [18,19].
From this point of view, H. pylori might function through a hit-and-run mechanism, predisposing the gastric mucosa to subsequent oncogenic alterations, which other microorganisms could facilitate [20].
In this review, we synthesize the most recent evidence on the microecological dysbiosis in gastric carcinogenesis, highlighting the mechanisms by which H. pylori and the non-Helicobacter gastric microbiota cooperate in the malignant transformation of the mucosa.
2. The Gastric Microbiota in Healthy Individuals
The discovery of Campylobacter pyloridis in 1982 demonstrated that bacteria could survive and thrive in the stomach’s acidic environment, definitively shifting the paradigm of the stomach as a sterile niche for bacteria [21]. Later, advancements in high-throughput and next-generation sequencing techniques confirmed that the stomach harbors a complex microbial community typical of the organ and different from that found in other parts of the digestive tract.
The gastric microbiota of healthy individuals comprises 57 bacterial genera across eight phyla, including Proteobacteria, Firmicutes, Bacteroidetes, Actinobacteria, Fusobacteria, Spirochetes, Tenericutes, and Saccharibacteria. The most prevalent genera for each phylum are Streptococcus and Veillonella (Firmicutes), Neisseria and Haemophilus (Proteobacteria), Fusobacterium (Fusobacteria), Prevotella and Porphyromonas (Bacteroidetes), and Rothia (Actinobacteria) [22,23,24]. Interestingly, more than 65% of the bacterial phylotypes in the stomach also inhabit the human mouth [25,26].
The richness and diversity of gastric microbiota significantly vary between mucosa and gastric juice and between antrum and corpus. In particular, the microbiota inhabiting gastric juice, primarily composed of Firmicutes, Bacteroidetes, and Actinobacteria phyla, does not accurately represent the diversity of stomach bacteria, as transient bacteria fail to colonize the gastric mucosa [27]. Furthermore, species richness in the corpus is notably higher than in the antral mucosa, albeit with no significant difference [28].
Several factors can influence the composition of gastric microbiota [29], such as dietary patterns [30], geographical location [31], prolonged use of proton pump inhibitors [32] or probiotics [33], surgical interventions [34], and, above all, H. pylori infection [35].
In Mongolian gerbils, H. pylori infection was associated with the occurrence of Enterococcus spp. and Staphylococcus aureus in both the stomach and duodenum. Additionally, infected animals exhibited a notable increase in Bacteroides spp. in the stomach, while Lactobacillus spp. had a decreasing trend. Moreover, the levels and distributions of Bifidobacterium spp., Bacteroides spp., and total aerobes changed. Intriguingly, gerbils positive for H. pylori colonization had a higher histopathologic score for gastritis and a similar score for duodenitis [36].
In the healthy human gastric mucosa, Streptococcus and Prevotella stand out as the dominant genera [22,23,24]. However, upon infection, H. pylori becomes the predominant gastric microbe, prevailing in up to 40–99% of the total microbial community. This explains up to 28% of individuals’ overall variation in gastric microbiota [37,38].
H. pylori infection negatively correlates with the alpha diversity of gastric microbiota and effectively clusters groups of infected and uninfected individuals based on the beta diversity [39]. In particular, H. pylori is accompanied by a lower abundance of Actinobacteria, Bacteroidetes, and Firmicutes and an increase in Proteobacteria [40,41,42]. Furthermore, there exists a negative association between H. pylori and some gastric members, such as Acidovorax, Aeromonas, Bacillus, Bradyrhizobium, Halomonas, Cloacibacterium, Meiothermus, Methylobacterium, and Ralstonia, while the interactions of the non-H. pylori members are positively correlated [43].
3. H. pylori and Cancer
H. pylori infection significantly increases the risk of non-cardia adenocarcinomas with an overall odds ratio (OR) of 21.0 [44,45,46]. Persistent colonization of the stomach by H. pylori induces a stepwise process that can eventually progress into GC. According to the gastric precancerous cascade, H. pylori primarily triggers the transition from normal mucosa to non-atrophic gastritis (NAG). Subsequently, it induces a series of progressive changes, starting with the loss of stomach-specific cell types (atrophic gastritis, AG), then advancing to intestinal metaplasia–dysplasia, and finally culminating in gastric adenocarcinoma. [4,5]. Several factors may contribute to H. pylori-induced gastric carcinogenesis, such as strain variation, genetic polymorphisms of hosts, and environmental influences.
Different allotypes of bacterial virulence factors, especially cytotoxin-associated gene A (CagA) and particular alleles of oligomeric vacuolating cytotoxin A (VacA), are closely associated with GC development. The CagA gene characterizes more virulent strains of H. pylori, increasing the risk of developing GC by 1.64-fold [47]. It encodes the CagA protein, which promotes tumorigenesis by activating inflammatory signaling and oncogenic pathways, stimulating cell proliferation, losing contact inhibition, and suppressing apoptosis [7,8,9,47]. VacA is a pore-forming cytotoxin secreted by H pylori, causing cell vacuolization and other cytotoxic features, such as alterations in plasma membrane permeability [10], paracellular leakage in epithelial monolayers [48], and mitochondria dysfunction [49]. VacA protein destroys the tight junctions of epithelial cells, induces apoptosis by interfering with mitochondrial function [4], and causes alteration in cell signaling [50,51]. The ability of VacA to downregulate autophagy and lysosomal degradation contributes to the accumulation of CagA in gastric epithelial cells [52]. Polymorphisms and epigenetic alterations in genes linked to both the adaptive and innate immune systems, such as interleukins (IL1β and IL8), transcription factors (CDX2, RUNX3, and TLR1), and DNA repair enzymes, can influence the individual’s susceptibility to GC development [11,12,13].
A Western dietary pattern, characterized by high red meat and salt intake and a low intake of antioxidant foods (fruit and vegetables), is associated with increased GC risk [14,15]. Habitual high salt intake increases susceptibility to H. pylori infection and can synergistically promote GC through glandular atrophy, DNA damage, and cell proliferation. Similarly, a poor-quality diet with consumption of salt-preserved smoked foods, rich in carcinogenic N-nitroso compounds, promotes cancer development [52]. A high-fat diet may induce gastric dysbiosis (characterized by an increase in Lactobacillus abundance), intestinal metaplasia; overexpression of leptin, phosphorylated leptin receptor, and STAT3; and intracellular b-catenin accumulation in GC [27]. Finally, other established factors associated with an increased GC risk include older age, low socioeconomic status, obesity, industrial and chemical pollutants, cigarette smoking, and heavy alcohol drinking [2].
4. Non-H. pylori Gastric Microbiota and Cancer
Robust evidence supporting the potential role of non-H. pylori microbiota in gastric carcinogenesis emerged from studies on animal models.
In C57BL/6N mice infected with the same strain of H. pylori but raised in different environments (i.e., Charles River Lab and Taconic farms), the incidence of severe inflammation, gastritis, and metaplasia was higher in those from Charles River Lab than in those from Taconic farm. The colonization rate by Lactobacillus strains differed significantly between the two groups, thus suggesting that mice grown in different environments harbor distinct stomach microbiota [53].
In the insulin–gastrin (INS-GAS) transgenic mouse model, mice harboring a gastric-specific pathogen-free microbiota developed more severe gastric precancerous lesions than those with H. pylori mono-infection [54]. The colonization with altered Schaedler’s Flora (a microbiota restricted to three species, including ASF356 Clostridium species, ASF361 Lactobacillus murinus, and ASF519 Bacteroides spp.) was sufficient to promote gland atrophy or dysplasia in mice. However, H. pylori coinfection led to more severe abnormalities, and 69% of mice with gastric dysplasia had intraepithelial neoplasia. In particular, the overgrowth of Lactobacillus murinus ASF361 correlated with a robust expression of gastric inflammation and cancer molecules, including TNF-α, Ptger4, and TGF-β [55]. On the other hand, antibiotic treatment significantly prevented the progression of gastric neoplasia in H. pylori-free and specific pathogen-free INS-GAS mice [56].
The transgenic K19-Wnt1/C2mE mice (GAN mice) express Wnt1, COX-2, and PGE2 in the gastric mucosa and develop gastric tumors with histological characteristics closely resembling those observed in human intestinal-type GC. Oshima et al. observed that GAN mice raised under germ-free conditions develop significantly smaller tumors than specific pathogen-free (SPF)-GAN mice. Likewise, antibiotic treatment of SPF-GAN mice suppressed gastric tumor development. In addition, the reconstitution of commensal bacteria in germ-free (GF)-GAN mice and the infection with Helicobacter felis led to the development of gastric tumors [57,58]. Finally, gastric microbiota tissue and fluids transplanted from human patients with intestinal metaplasia or GC induce precancerous features in GF- salivarius or Staphylococcus epidermidis causes increased gastric damage or reduced proinflammatory cytokine responses, respectively, compared to mice infected solely with H. pylori [59,60].
Based on this evidence, it seems plausible that non-H. pylori bacteria could contribute to developing gastric diseases through synergistic or competitive interactions with H. pylori or independent of it. However, the nature of the interaction between H. pylori and non-Helicobacter microbiota in gastric carcinogenesis still remains unclear.
One hypothesis proposes that chronic H. pylori infection, leading to gastric atrophy and achlorhydria, favors the overgrowth of non-Helicobacter species. These bacteria potentially fuel malignant transformation by perpetuating inflammation and generating carcinogenic N-nitroso compounds, including N-nitrosamines and nitric oxide, which induce DNA damage and reactive oxygen species (ROS) [61,62]. For example, Veillonella parvula and Haemophilus parainfluenzae could enhance nitrite accumulation in gastric juice due to their faster nitrate-reducing activity, causing DNA damage and methylation in epithelial cells. Similarly, Escherichia coli, Streptococcus, Clostridium, Haemophilus, Veillonella, Staphylococcus, Neisseria, Nitrospirae, and Lactobacillus could contribute to GC through the production of carcinogenic N-nitroso compounds [63]. Finally, lactic acid bacteria produce ROS, which induce DNA damage, reduce nitrate to nitrite, drive the oncogenes’ activation, enhance angiogenesis, and inhibit apoptosis [16].
Although data from animal studies provide biological plausibility for the hypothesis that the gastric microbiota can impact the type and timing of gastric lesions, human studies have not yielded conclusive results. The profile of the gastric microbiota at the time of diagnosis cannot provide insight into the causal role of bacteria in human GC. On the other hand, due to the long natural history of gastric diseases, conducting longitudinal studies to correlate specific bacterial signatures with various disease outcomes is not feasible.
In Colombia, where GC reaches one of the highest incidence and prevalence rates globally, infection by H. pylori is nearly universal [64]. However, among inhabitants of Tumaco on the Pacific Coast, GC’s incidence and mortality rates are significantly lower than those observed in the Túquerres Mountain region. The residents of mountainous areas and coastal regions exhibit significant differences in terms of population ancestry, lifestyle, diet, and prevalence of intestinal parasite infestations. Nevertheless, none of these factors, together with the phylogeographical group of H. pylori or the presence of cag pathogenicity island, provide a complete rationale for the so-called “Colombian Enigma” [65]. Interestingly, several operational taxonomic units (OTUs) were detected exclusively in either the Tumaco or Tùquerres population. Specifically, some taxa, such as Leptotrichia wadei and Veillonella spp. were more abundant, and Staphylococcus spp. were less abundant in gastric biopsies of individuals from Túquerres than those from Tumaco, highlighting the potential role of non-Helicobacter gastric microbiota in the carcinogenic process [31].
When GC develops, the composition of the resident microbiota significantly differs between cancerous and non-cancerous individuals.
In one of the first studies employing terminal restriction fragment length polymorphisms alongside 16S rRNA gene cloning and sequencing, the characterization of gastric microbiota in 10 GC patients and 5 dyspeptic controls revealed a relatively low abundance of H. pylori. Conversely, various species from the genera Streptococcus, Lactobacillus, Veillonella, and Prevotella were predominant in the GC microbiota [66]. Castaño-Rodríguez et al. found a marked increase in the relative abundances of Lactococcus, Fusobacterium, Veillonella, Haemophilus, and Leptochichia in GC patients compared to control samples. Of interest, predicted pathways related to bacterial carbohydrate metabolism, such as short-chain fatty acid production, were enriched in GC [67]. A more extensive study including 103 patients with GC compared to 212 with chronic gastritis (CG) found a markedly increased bacterial load in GC, albeit with no significant diversity at the phylum level. However, five genera with potential cancer-promoting activities, namely Lactobacillus, Escherichia–Shigella, Nitrospirae, Burkholderia fungorum, and Lachnospiraceae uncultured, were found to be enriched in GC samples. Interestingly, Nitrospirae was found in all GC patients and was absent in CG cases [68].
The microbial profiling of the gastric mucosa of 54 patients with GC compared to 81 patients with CG revealed reduced bacterial diversity and Helicobacter abundance in GC cases. Conversely, commensal genera such as Achromobacter, Citrobacter, Clostridium, Lactobacillus, and Rhodococcus were over-represented. The combination of ten relevant bacteria with a different abundance in GC and CG established a microbial dysbiosis index that demonstrated a strong performance in effectively distinguishing CG and GC cases. Interestingly, the microbial communities identified in GC had nitrosation functions, indicative of increased genotoxic potential [17]. Coker et al. obtained similar results by identifying five non-H. pylori bacterial taxa of oral origin (Peptostreptococcus stomatis, Streptococcus anginosus, Parvimonas micra, Slackia exigua, and Dialister pneumosintes), which exhibited significant centrality in the GC microbial ecological network and distinguished GC from superficial gastritis with optimal diagnostic performance (area under the receiver operating curve [AUC] = 0.82) [69]. Finally, in a population of 268 early GC cases, the relative abundance of H. pylori, Propionibacterium acnes, and Prevotella copri in the stomach was higher than that of non-cancer-bearing people, thus identifying these taxa as strong risk factors for GC development. In contrast, Lactobacillus lactis was protective against GC [70].
Despite the importance of the findings, all previous studies are affected by inherent bias stemming from the comparison between different patient cohorts, specifically those with GC and cancer-free individuals. In such cases, variations in environmental factors and genetic backgrounds can significantly influence the composition of the microbiota. Therefore, some researchers chose a paired design approach, wherein the gastric microbiota was analyzed comparatively in the same patient’s tumor and non-malignant tissues.
Once again, the bacterial community differed considerably from non-tumoral and tumoral tissue in GC patients [71,72,73,74].
In cancer tissue, H. pylori, Propionibacterium spp., Staphylococcus spp., and Corynebacterium spp. were significantly reduced compared to normal gastric mucosa, while Clostridium spp. and Prevotella spp. were significantly increased [72]. Oral bacteria such as Peptostreptococcus, Streptococcus, and Fusobacterium were predominant among the enriched taxa in GC samples. At the same time, there were elevated levels of lactic acid-producing bacteria in the adjacent non-tumor tissues, including Lactococcus lactis and Lactobacillus brevis [73]. In a retrospective cohort study including 276 patients with GC, Liu et al. found that the bacterial richness was low in both the peritumoral and tumoral microhabitat. However, H. pylori, Prevotella copri, and Bacteroides uniformis significantly decreased, and Prevotella melaninogenica, Streptococcus anginosus, and Propionibacterium acnes significantly increased in the tumoral microhabitat. The specific stomach microenvironment, i.e., distinct pH, oxygen, nutrients, ions, and chemicals of tumor tissues and adjacent tumor-free tissues, were the main determinant of the composition and diversity of the gastric microbiota, providing a setting for symbiotic interactions among the various microbes within that ecosystem and the host [74].
Finally, the gastric microbiota profile differs significantly between early GC and advanced GC. Wang et al. demonstrated that Burkholderia, Tsukamurella, Uruburuella, and Salinivibrio varied in their relative abundance between the early and advanced stages of GC. Functional analyses showed that urease and bacterial flagella synthesis were impaired in early GC, whereas fructose glycolysis and glycoside hydrolysis were enhanced. Interestingly, the microbial signature could accurately distinguish early GC from chronic gastritis or advanced GC in an independent cohort of patients [75].
5. Microbiota Changes during the Multistep Process of Gastric Carcinogenesis
Changes in the gastric microenvironment, alterations in host immune responses, and the progression of GC can account for the different microecology of the stomach. In the early GC, the microbiota composition might reflect the initial stages of tumor development, with specific bacterial species potentially promoting or inhibiting tumor growth. As cancer progresses to advanced stages, the tumor microenvironment undergoes further changes, which can influence the composition of the gastric microbiota. Additionally, treatments such as chemotherapy and radiation therapy can impact microbiota composition. Overall, the dynamic interplay between the host, the tumor, and the microbiota likely contributes to the observed differences between early and advanced GC.
Notably, the gastric microbiota undergoes diversity and composition alterations also through the different stages of the gastric precancerous cascade [76], suggesting that dysbiosis worsens as the carcinogenesis process advances.
Aviles-Jimenez et al. first demonstrated a gradual shift in microbiota composition from NAG to intestinal metaplasia and GC. In particular, bacterial diversity steadily decreased as patients progressed from NAG to intestinal metaplasia and GC. At the genus level, Porphyromonas, Neisseria, TM7, and Streptococcus sinensis had a decreasing trend, while Lactobacillus coleohominis and Lachnospiraceae increased, suggesting that these alterations might favor gastric tumorigenesis. Interestingly, the microbiota segregated between NAG and GC, with 44 taxa showing significant changes in relative abundance [77].
In another study, GC patients had an increase in the Bacilli class and Streptococcaceae family and a reduced Epsilonproteobacteria class and Helicobacteraceae family compared to those with chronic gastritis and intestinal metaplasia [78].
To what extent changes in gastric microbiota are influenced by histological alterations of the mucosa or the presence of H. pylori is not entirely understood.
Li et al. characterized the microbiota alterations through different histological stages of gastric carcinogenesis and after H. pylori eradication. Apart from dominant H. pylori, Proteobacteria, including Haemophilus, Serratia, Neisseria, and Stenotrophomonas, were the major components of the human gastric microbiota. Interestingly, there was a strong negative correlation between the relative abundance of H. pylori and bacterial diversity. However, in GC patients, there was a more pronounced decreasing trend in bacterial diversity compared to other precancerous lesions with comparable levels of H. pylori. Interestingly, eradicating H. pylori infection restored the gastric microenvironment to that of H. pylori-negative subjects, including a similar phyla composition and increased bacterial diversity index [79]. In H. pylori-negative cases, across precancerous stages from atrophic gastritis to dysplasia, Burkholderiaceae abundance constantly increased, while the abundance of Streptococcaceae and Prevotellaceae had a continuous decreasing trend. Patients with intestinal metaplasia and those with dysplasia had similar gastric mucosa microbiota profiles, with Ralstonia and Rhodococcus as the predominant genera. In contrast, GC had the lowest bacterial richness, with Streptococcaceae and Lactobacillaceae being the associated bacteria. Gastric juice had a higher microbial diversity than the mucosa, with Proteobacteria prevalent in the gastric mucosa and Firmicutes in gastric juice [27].
Comprehensive profiling of gastric mucosa and gastric juice showed a notable shift in microbial composition in both samples and a convergent dysbiosis during the progression from superficial gastritis to GC. This suggests that the colonization of carcinogenic microbes in the mucosa might originate from gastric juice and underscores the potential of gastric juice as a promising source of biomarkers [80,81]. Interestingly, a panel of GC classifiers, including Gemella, Haemophilus, Peptostreptococcus, Streptococcus, and Veillonella, exhibited considerable changes and performed well in an independent cohort, proving their effectiveness in identifying GC [80].
Relevant studies addressing the difference in gastric microbiota composition across the histological stages of gastric carcinogenesis are summarized in Table 1.

Table 1.
Changes in gastric microbiota in carcinogenesis.
However, differences in analytical methods, bioinformatic techniques, sampling strategies, participant demographics, target regions of the 16S rRNA gene, and sequencing platforms contribute to significant variability in the results, preventing the generalizability of the conclusions [85,86].
A meta-analysis explored the global microbial signature associated with gastric carcinogenesis by pooling data from six independent studies, including 825 gastric biopsy samples. Five phyla, Proteobacteria, Firmicutes, Bacteroidetes, Actinobacteria, and Fusobacteria, dominated the gastric microbiota in descending order of relative abundance. At the genus level, ten genera, namely Helicobacter, Halomonas, Pseudomonas, Streptococcus, Lactobacillus, Shewanella, Prevotella, Acinetobacter, Cryocola, and Staphylococcus, dominated the gastric mucosal microbiota. The alpha diversity decreased along the disease stages, with GC showing the lowest alpha diversity compared to superficial gastritis, atrophic gastritis, and intestinal metaplasia. Interestingly, the microbial composition of the four disease stages significantly clustered according to beta-diversity indexes. Opportunistic pathogens such as Fusobacterium, Parvimonas, Veillonella, Prevotella, and Peptostreptococcus were enriched in GC. At the same time, the commensals Bifidobacterium, Bacillus, and Blautia were depleted in GC compared to superficial gastritis. Overall, six GC-enriched (Veillonella, Dialister, Granulicatella, Herbaspirillum, Comamonas, and Chryseobacterium) and two GC-depleted genera (Shewanella and Helicobacter) were identified as potential biomarkers for discriminating GC from superficial gastritis, yielding an outstanding diagnostic performance (AUC = 0.91 for the training set and 0.85 for the test set, respectively). Except for Veillonella, Herbaspirillum, and Shewanella, all the evaluated biomarkers significantly differed between GC and superficial gastritis in Asian and European populations. Moreover, H. pylori-positive samples exhibited reduced microbial diversity, altered microbiota community, and weaker interactions among gastric microbes [87].
A more recent meta-analysis of nine public datasets of human gastric microbiota confirmed that the gastric microbiome composition significantly shifted during the progression of gastric carcinogenesis, accompanied by a gradual decline in microbial diversity. Using the random effect model, the authors identified a set of universal microbial biomarkers that differentiate GC and gastritis and have the potential for GC diagnosis. Key microbial signatures included Fusobacterium; Leptotrichia; and several lactic acid bacteria, such as Bifidobacterium, Lactobacillus, and Streptococcus anginosus [20].
In summary, the microbiota profiles in patients with H. pylori-induced superficial gastritis or glandular atrophy are characterized by the dominance of Helicobacter and, to a much lesser extent, Streptococcus, Prevotella, and Neisseria with a decreased phylotype richness, diversity, and evenness when compared with patients with normal gastric mucosa. The loss of specialized glandular tissue and reduced acid secretion in GC tissue results in H. pylori loss and the enrichment of intestinal commensals, including Lactobacillus, Enterococci, Carnobacterium, Parvimonas, Citrobacter, Clostridium, Achromobacter, and Rhodococcus, as well as the oral species Fusobacterium nucleatum, Veillonella, Leptotrichia, Haemophilus, and Campylobacter [63].
As expected, functional alterations accompany the progressive dysbiosis through gastric disease stages. The samples from GC patients were significantly enriched in many metabolic pathways, including those related to purine metabolism [73], carbohydrate metabolism (such as short-chain fatty acid production) [63,67] or glycolysis and glycoside hydrolysis [75], lipopolysaccharide (LPS) and L-arginine biosynthesis [27], amino acid and nitrate metabolism [17,63], and peptidoglycan biosynthesis [20]. These findings align with the increased energy metabolism and concentration of nitrogen-containing compounds in the tumor microenvironment [73]. In contrast, pathways involved in the fermentation of short-chain fatty acids and branched amino acid metabolism were more abundant in superficial gastritis [27].
Overall, the data that emerged from both animal and human studies do not definitively clarify whether the microbial changes observed in GC are causative in the development of the disease or if they result from the histologic progression through the precancerous cascade. Although a unique profile of gastric microbiota associated with GC development remains elusive, numerous taxa, including members of the oral and intestinal microbial communities, are reported to be enriched in the gastric microbial communities of cancer patients. This suggests a complex evolution of gastric carcinogenesis potentially coordinated by H. pylori in cooperation with other microbial species contributing to the process (Figure 1).
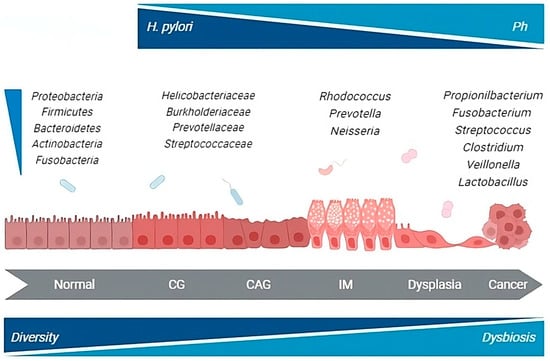
Figure 1.
H. pylori and gastric microbiota alterations throughout the carcinogenic process. CG, chronic gastritis; CAG, chronic atrophic gastritis; IM, intestinal metaplasia. Gastric microbiota changes through the different steps of the carcinogenic cascade. GC-associated microbiota is enriched in species from the oral and intestinal microbial communities that may possess nitrosation functions.
6. Effects of H. pylori Eradication on the Gastric Microbiome and Patient’s Outcome
The data on the effect of H. pylori eradication on gastric microbiome are somewhat conflicting.
A recent meta-analysis including nine studies and 546 participants showed that H. pylori eradication was associated with increased alpha diversity and restored the composition of commensals commonly dominant in the healthy stomach, independently of the therapeutic schedule used and length of follow-up [88]. Since the reduction of commensal microbial communities is a hallmark of GC risk, it is conceivable to hypothesize that restoring the abundance of Firmicutes, Bacteroides, and Actinobacteria might be one of the mechanisms by which H. pylori eradication reduces GC risk.
On the other hand, in patients with endoscopic follow-up for >1 year, eradicating H. pylori infection led to the predominance of proinflammatory Acinetobacter in gastric corpus mucosa with a decrease in microbial diversity in about half of the cases [89]. Moreover, after H. pylori eradication, the abundance of some bacteria predominantly of oral origin, such as Actinomyces, Granulicatella or Peptostreptococcus, was associated with precancerous gastric lesions, thus significantly increasing the risk of GC [90].
Based on the available evidence, a careful assessment of the individual benefit–risk ratio should replace the unconditional “test-and-treat” strategy in eradicating H. pylori infection.
7. Conclusions and Perspectives
The stomach hosts a complex core microbiota, and H. pylori and non-Helicobacter gastric microbiota might be interactive contributors rather than acting autonomously in the malignant alteration of gastric tissues. Similar to the driver–passenger hypothesis proposed in colorectal cancer and gut microbiota interactions [91], H. pylori could trigger the gastric carcinogenic process. At the same time, other bacteria may damage gastric mucosa, thus promoting the progression of gastric precancerous lesions to GC.
Although progress has been made in characterizing the human gastric microbiota in recent years, definitive conclusions are hindered by the inherent biases of the current literature. Evidence mainly derives from case-control studies comparing gastric microbiota signatures from GC patients and healthy individuals. Additionally, most studies are limited by small sample sizes, heterogeneous enrolled populations, and cancer diagnoses that do not differentiate between histological types or previous treatments. Finally, research on the gastric virome and mycome is less extensive than bacterial studies, and only in recent years has it gained interest.
At the current state of our knowledge, the most critical question remains whether the alterations in the composition of the gastric microbiota are causal or consequential to the development of gastric precancerous lesions and GC so that we cannot infer a causal relationship between dysbiosis and the development and progression of GC. In addition, definitive conclusions cannot be drawn on the beneficial or harmful effects of H. pylori eradication on gastric microbiota.
Through in-depth research, considering large-scale, multicenter, cohort prospective studies with long-term follow-up could significantly contribute to elucidating the dynamic changes in the microbiota along the gastric carcinogenic process and possibly identify specific microorganisms that are potentially useful as biomarkers for GC diagnosis and prognosis. Furthermore, a complex biological process such as GC should be studied holistically, using an integrative approach that combines multi-omics data to highlight the interrelationships of the involved biomolecules and their functions.
An integrative approach could clarify the mechanisms by which microbes impact cancer development and prognosis. Finally, a better understanding of the impact of eradicating H. pylori infection on gastric microecology could help to personalize indication to therapy. As our knowledge advances, new avenues for the early detection and treatment of GC will become available, offering guidance for determining personalized treatment strategies and improving patient outcomes.
Author Contributions
C.S., A.R., D.C. and K.P. reviewed the current literature on the topic; C.S., A.R. and D.C. wrote the original draft; A.R., G.N. and M.R. reviewed and supervised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The figure was created with BioRender.com (accessed on 18 April 2024).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.; Lordick, F. Gastric Cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Fock, K.M.; Graham, D.Y.; Malfertheiner, P. Helicobacter Pylori Research: Historical Insights and Future Directions. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Correa, P. A Human Model of Gastric Carcinogenesis. Cancer Res. 1986, 48, 3554–3560. [Google Scholar]
- Yang, H.; Wei, B.; Hu, B. Chronic Inflammation and Long-Lasting Changes in the Gastric Mucosa after Helicobacter Pylori Infection Involved in Gastric Cancer. Inflamm. Res. 2021, 70, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N. The Magnitude of Association Between Helicobacter Pylori Infection and the Development of Gastric Cancer. Scand. J. Gastroenterol. 2002, 37, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Brandt, S.; Kwok, T.; Hartig, R.; König, W.; Backert, S. NF-κB Activation and Potentiation of Proinflammatory Responses by the Helicobacter Pylori CagA Protein. Proc. Natl. Acad. Sci. USA 2005, 102, 9300–9305. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Ooki, T.; Murata-Kamiya, N.; Komura, D.; Tahmina, K.; Wu, W.; Takahashi-Kanemitsu, A.; Knight, C.T.; Kunita, A.; Suzuki, N.; et al. Helicobacter Pylori CagA Elicits BRCAness to Induce Genome Instability That May Underlie Bacterial Gastric Carcinogenesis. Cell Host Microbe 2021, 29, 941–958.e10. [Google Scholar] [CrossRef] [PubMed]
- Yong, X.; Tang, B.; Li, B.-S.; Xie, R.; Hu, C.-J.; Luo, G.; Qin, Y.; Dong, H.; Yang, S.-M. Helicobacter Pylori Virulence Factor CagA Promotes Tumorigenesis of Gastric Cancer via Multiple Signaling Pathways. Cell Commun. Signal. 2015, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I. Formation of Anion-Selective Channels in the Cell Plasma Membrane by the Toxin VacA of Helicobacter Pylori Is Required for Its Biological Activity. EMBO J. 1999, 18, 5517–5527. [Google Scholar] [CrossRef]
- El-Omar, E.M.; Carrington, M.; Chow, W.-H.; McColl, K.E.L.; Bream, J.H.; Young, H.A.; Herrera, J.; Lissowska, J.; Yuan, C.-C.; Rothman, N.; et al. Interleukin-1 Polymorphisms Associated with Increased Risk of Gastric Cancer. Nature 2000, 404, 398–402. [Google Scholar] [CrossRef]
- Hou, L.; El-Omar, E.M.; Chen, J.; Grillo, P.; Rabkin, C.S.; Baccarelli, A.; Yeager, M.; Chanock, S.J.; Zatonski, W.; Sobin, L.H.; et al. Polymorphisms in Th1-Type Cell-Mediated Response Genes and Risk of Gastric Cancer. Carcinogenesis 2007, 28, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Companioni, O.; Bonet, C.; Muñoz, X.; Weiderpass, E.; Panico, S.; Tumino, R.; Palli, D.; Agnoli, C.; Vineis, P.; Boutron-Ruault, M.C.; et al. Polymorphisms of Helicobacter Pylori Signaling Pathway Genes and Gastric Cancer Risk in the European Prospective Investigation into Cancer-Eurgast Cohort. Int. J. Cancer 2014, 134, 92–101. [Google Scholar] [CrossRef]
- Gonázlez, C.A.; López-Carrillo, L. Helicobacter Pylori, Nutrition and Smoking Interactions: Their Impact in Gastric Carcinogenesis. Scand. J. Gastroenterol. 2010, 45, 6–14. [Google Scholar] [CrossRef]
- Venneman, K.; Huybrechts, I.; Gunter, M.J.; Vandendaele, L.; Herrero, R.; Van Herck, K. The Epidemiology of Helicobacter Pylori Infection in Europe and the Impact of Lifestyle on Its Natural Evolution toward Stomach Cancer after Infection: A Systematic Review. Helicobacter 2018, 23, e12483. [Google Scholar] [CrossRef]
- Wang, L.L.; Yu, X.J.; Zhan, S.H.; Jia, S.J.; Tian, Z.B.; Dong, Q.J. Participation of Microbiota in the Development of Gastric Cancer. World J. Gastroenterol. 2014, 20, 4948–4952. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; MacHado, J.C.; Figueiredo, C. Gastric Microbial Community Profiling Reveals a Dysbiotic Cancer-Associated Microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Mowat, C.; Williams, C.; Gillen, D.; Hossack, M.; Gilmour, D.; Carswell, A.; Wirz, A.; Preston, T.; McColl, K.E.L. Omeprazole, Helicobacter Pylori Status, and Alterations in the Intragastric Milieu Facilitating Bacterial N-Nitrosation. Gastroenterology 2000, 119, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Keszei, A.P.; Goldbohm, R.A.; Schouten, L.J.; Jakszyn, P.; Van Den Brandt, P.A. Dietary N-Nitroso Compounds, Endogenous Nitrosation, and the Risk of Esophageal and Gastric Cancer Subtypes in the Netherlands Cohort Study. Am. J. Clin. Nutr. 2013, 97, 135–146. [Google Scholar] [CrossRef]
- Li, Y.; Hu, Y.; Zhan, X.; Song, Y.; Xu, M.; Wang, S.; Huang, X.; Xu, Z.Z. Meta-Analysis Reveals Helicobacter Pylori Mutual Exclusivity and Reproducible Gastric Microbiome Alterations during Gastric Carcinoma Progression. Gut Microbes 2023, 15, 1024–1032. [Google Scholar] [CrossRef]
- Robin Warren, J.; Marshall, B. Unidentified Curved Bacilli on Gastric Epithelium in Active Chronic Gastritis. Lancet 1983, 321, 1273–1275. [Google Scholar] [CrossRef]
- Bik, E.M.; Eckburg, P.B.; Gill, S.R.; Nelson, K.E.; Purdom, E.A.; Francois, F.; Perez-Perez, G.; Blaser, M.J.; Relman, D.A. Molecular Analysis of the Bacterial Microbiota in the Human Stomach. Proc. Natl. Acad. Sci. USA 2006, 103, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Wong, G.L.H.; To, K.F.; Wong, V.W.S.; Lai, L.H.; Chow, D.K.L.; Lau, J.Y.W.; Sung, J.J.Y.; Ding, C. Bacterial Microbiota Profiling in Gastritis without Helicobacter Pylori Infection or Non-Steroidal Anti-Inflammatory Drug Use. PLoS ONE 2009, 4, e7985. [Google Scholar] [CrossRef] [PubMed]
- Delgado, S.; Cabrera-Rubio, R.; Mira, A.; Suárez, A.; Mayo, B. Microbiological Survey of the Human Gastric Ecosystem Using Culturing and Pyrosequencing Methods. Microb. Ecol. 2013, 65, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Nardone, G.; Compare, D.; Rocco, A. A Microbiota-Centric View of Diseases of the Upper Gastrointestinal Tract. Lancet Gastroenterol. Hepatol. 2017, 2, 298–312. [Google Scholar] [CrossRef]
- Bassis, C.M.; Erb-Downward, J.R.; Dickson, R.P.; Freeman, C.M.; Schmidt, T.M.; Young, V.B.; Beck, J.M.; Curtis, J.L.; Huffnagle, G.B. Analysis of the Upper Respiratory Tract Microbiotas as the Source of the Lung and Gastric Microbiotas in Healthy Individuals. MBio 2015, 6, e00037-15. [Google Scholar] [CrossRef]
- Sun, Q.-H.; Zhang, J.; Shi, Y.-Y.; Zhang, J.; Fu, W.-W.; Ding, S.-G. Microbiome Changes in the Gastric Mucosa and Gastric Juice in Different Histological Stages of Helicobacter Pylori-Negative Gastric Cancers. World J. Gastroenterol. 2022, 28, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.-H.; Kim, N.; Jo, H.J.; Kim, J.; Park, J.H.; Nam, R.H.; Seok, Y.-J.; Kim, Y.-R.; Lee, D.H. Analysis of Gastric Body Microbiota by Pyrosequencing: Possible Role of Bacteria Other Than Helicobacter Pylori in the Gastric Carcinogenesis. J. Cancer Prev. 2017, 22, 115–125. [Google Scholar] [CrossRef]
- Nardone, G.; Compare, D. The Human Gastric Microbiota: Is It Time to Rethink the Pathogenesis of Stomach Diseases? United Eur. Gastroenterol. J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Gunathilake, M.; Lee, J.H.; Choi, I.J.; Kim, Y.I.; Kim, J.S. Effect of the Interaction between Dietary Patterns and the Gastric Microbiome on the Risk of Gastric Cancer. Nutrients 2021, 13, 2692. [Google Scholar] [CrossRef]
- Yang, I.; Woltemate, S.; Piazuelo, M.B.; Bravo, L.E.; Yepez, M.C.; Romero-Gallo, J.; Delgado, A.G.; Wilson, K.T.; Peek, R.M.; Correa, P.; et al. Different Gastric Microbiota Compositions in Two Human Populations with High and Low Gastric Cancer Risk in Colombia. Sci. Rep. 2016, 6, 18594. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, A.; Suda, W.; Morita, H.; Takanashi, K.; Takagi, A.; Koga, Y.; Hattori, M. Influence of Proton-Pump Inhibitors on the Luminal Microbiota in the Gastrointestinal Tract. Clin. Transl. Gastroenterol. 2015, 6, e89. [Google Scholar] [CrossRef] [PubMed]
- Nakae, H.; Tsuda, A.; Matsuoka, T.; Mine, T.; Koga, Y. Gastric Microbiota in the Functional Dyspepsia Patients Treated with Probiotic Yogurt. BMJ Open Gastroenterol. 2016, 3, e000109. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Lin, J.T.; Ho, H.J.; Lai, Z.L.; Wang, C.B.; Tang, S.L.; Wu, C.Y. Gastric Microbiota and Predicted Gene Functions Are Altered after Subtotal Gastrectomy in Patients with Gastric Cancer. Sci. Rep. 2016, 6, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, T.; Llorca, L.; Perez-Perez, G. Impact of the Microbiota and Gastric Disease Development by Helicobacter Pylori. Curr. Top. Microbiol. Immunol. 2017, 400, 253–275. [Google Scholar] [PubMed]
- Yin, Y.N.; Wang, C.L.; Liu, X.W.; Cui, Y.; Xie, N.; Yu, Q.F.; Li, F.J.; Lu, F.G. Gastric and Duodenum Microflora Analysis after Long-Term Helicobacter Pylori Infection in Mongolian Gerbils. Helicobacter 2011, 16, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Schütte, K.; Koch, N.; Vilchez-Vargas, R.; Wos-Oxley, M.L.; Oxley, A.P.A.; Vital, M.; Malfertheiner, P.; Pieper, D.H. The Active Bacterial Assemblages of the Upper GI Tract in Individuals with and without Helicobacter Infection. Gut 2018, 67, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Contreras, A.; Goldfarb, K.C.; Godoy-Vitorino, F.; Karaoz, U.; Contreras, M.; Blaser, M.J.; Brodie, E.L.; Dominguez-Bello, M.G. Structure of the Human Gastric Bacterial Community in Relation to Helicobacter Pylori Status. ISME J. 2011, 5, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Llorca, L.; Pérez-Pérez, G.; Urruzuno, P.; Martinez, M.J.; Iizumi, T.; Gao, Z.; Sohn, J.; Chung, J.; Cox, L.; Simón-Soro, A.; et al. Characterisation of the Gastric Microbiota in a Pediatric Population According to Helicobacter Pylori Status. Pediatr. Infect. Dis. J. 2017, 36, 173–178. [Google Scholar] [CrossRef]
- Miao, R.; Wan, C.; Wang, Z. The Relationship of Gastric Microbiota and Helicobacter Pylori Infection in Pediatrics Population. Helicobacter 2020, 25, e12676. [Google Scholar] [CrossRef]
- Rajilic-Stojanovic, M.; Figueiredo, C.; Smet, A.; Hansen, R.; Kupcinskas, J.; Rokkas, T.; Andersen, L.; Machado, J.C.; Ianiro, G.; Gasbarrini, A.; et al. Systematic Review: Gastric Microbiota in Health and Disease. Aliment. Pharmacol. Ther. 2020, 51, 582–602. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, M.; Tohumcu, E.; Del Vecchio, L.E.; Porcari, S.; Cammarota, G.; Gasbarrini, A.; Ianiro, G. The Influence of Helicobacter Pylori on Human Gastric and Gut Microbiota. Antibiotics 2023, 12, 765. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Pereira, V.; Saxena, S.; Ghosh, T.S.; Anbumani, D.; Bag, S.; Das, B.; Nair, G.B.; Abraham, P.; Mande, S.S. Gastric Microbiome of Indian Patients with Helicobacter Pylori Infection, and Their Interaction Networks. Sci. Rep. 2017, 7, 15438. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.E.; Engstrand, L.; Nyrén, O.; Evans, D.J.; Lindgren, A.; Bergström, R.; Andersson, B.; Athlin, L.; Bendtsen, O.; Tracz, P. Helicobacter Pylori Infection: Independent Risk Indicator of Gastric Adenocarcinoma. Gastroenterology 1993, 105, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Ekström, A.M.; Held, M.; Hansson, L.E.; Engstrand, L.; Nyrén, O. Helicobacter Pylori in Gastric Cancer Established by CagA Immunoblot as a Marker of Past Infection. Gastroenterology 2001, 121, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Okamoto, S.; Yamamoto, S.H. Pylori Infection and the Development of Gastric Cancer. Keio J. Med. 2002, 51, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Casellas, F.; Aparici, A.; Casaus, M.; Rodríguez, P.; Malagelada, J.R. Subjective Perception of Lactose Intolerance Does Not Always Indicate Lactose Malabsorption. Clin. Gastroenterol. Hepatol. 2010, 8, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Papini, E.; Satin, B.; Norais, N.; De Bernard, M.; Telford, J.L.; Rappuoli, R.; Montecucco, C. Selective Increase of the Permeability of Polarized Epithelial Cell Monolayers by Helicobacter Pylori Vacuolating Toxin. J. Clin. Investig. 1998, 102, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Papini, E.; de Bernard, M.; Milia, E.; Bugnoli, M.; Zerial, M.; Rappuoli, R.; Montecucco, C. Cellular Vacuoles Induced by Helicobacter Pylori Originate from Late Endosomal Compartments. Proc. Natl. Acad. Sci. USA 1994, 91, 9720–9724. [Google Scholar] [CrossRef]
- Burkitt, M.D.; Duckworth, C.A.; Williams, J.M.; Pritchard, D.M. Helicobacter Pylori-Induced Gastric Pathology: Insights from in Vivo and Ex Vivo Models. Dis. Model. Mech. 2017, 10, 89–104. [Google Scholar] [CrossRef]
- Raju, D.; Hussey, S.; Ang, M.; Terebiznik, M.R.; Sibony, M.; Galindo-Mata, E.; Gupta, V.; Blanke, S.R.; Delgado, A.; Romero-Gallo, J.; et al. Vacuolating Cytotoxin and Variants in Atg16L1 That Disrupt Autophagy Promote Helicobacter Pylori Infection in Humans. Gastroenterology 2012, 142, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Greenfield, L.K.; Bronte-Tinkew, D.; Capurro, M.I.; Rizzuti, D.; Jones, N.L. VacA Promotes CagA Accumulation in Gastric Epithelial Cells during Helicobacter Pylori Infection. Sci. Rep. 2019, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Rolig, A.S.; Cech, C.; Ahler, E.; Carter, J.E.; Ottemann, K.M. The Degree of Helicobacter Pylori-Triggered Inflammation Is Manipulated by Preinfection Host Microbiota. Infect. Immun. 2013, 81, 1382–1389. [Google Scholar] [CrossRef]
- Lofgren, J.L.; Whary, M.T.; Ge, Z.; Muthupalani, S.; Taylor, N.S.; Mobley, M.; Potter, A.; Varro, A.; Eibach, D.; Suerbaum, S.; et al. Lack of Commensal Flora in Helicobacter pylori–Infected INS-GAS Mice Reduces Gastritis and Delays Intraepithelial Neoplasia. Gastroenterology 2011, 140, 210–220.e4. [Google Scholar] [CrossRef] [PubMed]
- Lertpiriyapong, K.; Whary, M.T.; Muthupalani, S.; Lofgren, J.L.; Gamazon, E.R.; Feng, Y.; Ge, Z.; Wang, T.C.; Fox, J.G. Gastric Colonisation with a Restricted Commensal Microbiota Replicates the Promotion of Neoplastic Lesions by Diverse Intestinal Microbiota in the Helicobacter Pylori INS-GAS Mouse Model of Gastric Carcinogenesis. Gut 2014, 63, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Rickman, B.; Rogers, A.B.; Muthupalani, S.; Takaishi, S.; Peiying, Y.; Wang, T.C.; Fox, J.G. Combination of Sulindac and Antimicrobial Eradication of Helicobacter Pylori Prevents Progression of Gastric Cancer in Hypergastrinemic INS-GAS Mice. Cancer Res. 2009, 69, 8166–8174. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Matsunaga, A.; Fujimura, T.; Tsukamoto, T.; Taketo, M.M.; Oshima, M. Carcinogenesis in Mouse Stomach by Simultaneous Activation of the Wnt Signaling and Prostaglandin E2 Pathway. Gastroenterology 2006, 131, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Hioki, K.; Popivanova, B.K.; Oguma, K.; Van Rooijen, N.; Ishikawa, T.; Oshima, M. Prostaglandin E2 Signaling and Bacterial Infection Recruit Tumor-Promoting Macrophages to Mouse Gastric Tumors. Gastroenterology 2011, 140, 596–607.e7. [Google Scholar] [CrossRef]
- Kwon, S.-K.; Park, J.C.; Kim, K.H.; Yoon, J.; Cho, Y.; Lee, B.; Lee, J.-J.; Jeong, H.; Oh, Y.; Kim, S.-H.; et al. Human Gastric Microbiota Transplantation Recapitulates Premalignant Lesions in Germ-Free Mice. Gut 2022, 71, 1266–1276. [Google Scholar] [CrossRef]
- Shen, Z.; Dzink-Fox, J.; Feng, Y.; Muthupalani, S.; Mannion, A.J.; Sheh, A.; Whary, M.T.; Holcombe, H.R.; Piazuelo, B.M.; Bravo, L.E.; et al. Gastric Non-Helicobacter Pylori Urease-Positive Staphylococcus Epidermidis and Streptococcus Salivarius Isolated from Humans Have Contrasting Effects on H. Pylori-Associated Gastric Pathology and Host Immune Responses in a Murine Model of Gastric Cancer. mSphere 2022, 7, e0077221. [Google Scholar] [CrossRef]
- Hecht, S.S. DNA Adduct Formation from Tobacco-Specific N-Nitrosamines. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 1999, 424, 127–142. [Google Scholar] [CrossRef]
- Tsujiuchi, T.; Masaoka, T.; Sugata, E.; Onishi, M.; Fujii, H.; Shimizu, K.; Honoki, K. Hypermethylation of the Dal-1 Gene in Lung Adenocarcinomas Induced by N-Nitrosobis (2-Hydroxypropyl)amine in Rats. Mol. Carcinog. 2007, 46, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Smet, A.; Kupcinskas, J.; Link, A.; Hold, G.L.; Bornschein, J. The Role of Microbiota in Gastrointestinal Cancer and Cancer Treatment: Chance or Curse? Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 857–874. [Google Scholar] [CrossRef] [PubMed]
- Parra-Lara, L.G.; Falla-Martínez, J.C.; Isaza-Pierotti, D.F.; Mendoza-Urbano, D.M.; Tangua-Arias, A.R.; Bravo, J.C.; Bravo, L.E.; Zambrano, Á.R. Gastric Adenocarcinoma Burden, Trends and Survival in Cali, Colombia: A Retrospective Cohort Study. Front. Oncol. 2023, 13, 1069369. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L. Virulence-Associated Genotypes of Helicobacter Pylori: Do They Explain the African Enigma? Am. J. Gastroenterol. 2002, 97, 2839–2842. [Google Scholar] [CrossRef] [PubMed]
- Dicksved, J.; Lindberg, M.; Rosenquist, M.; Enroth, H.; Jansson, J.K.; Engstrand, L. Molecular Characterization of the Stomach Microbiota in Patients with Gastric Cancer and in Controls. J. Med. Microbiol. 2009, 58, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Rodríguez, N.; Goh, K.L.; Fock, K.M.; Mitchell, H.M.; Kaakoush, N.O. Dysbiosis of the Microbiome in Gastric Carcinogenesis. Sci. Rep. 2017, 7, 15957. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, J.; Xin, Y.; Geng, C.; Tian, Z.; Yu, X.; Dong, Q. Bacterial Overgrowth and Diversification of Microbiota in Gastric Cancer. Eur. J. Gastroenterol. Hepatol. 2016, 28, 261–266. [Google Scholar] [CrossRef]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal Microbiome Dysbiosis in Gastric Carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef]
- Gunathilake, M.N.; Lee, J.; Choi, I.J.; Kim, Y.I.; Ahn, Y.; Park, C.; Kim, J. Association between the Relative Abundance of Gastric Microbiota and the Risk of Gastric Cancer: A Case-Control Study. Sci. Rep. 2019, 9, 13589. [Google Scholar] [CrossRef]
- Yu, G.; Torres, J.; Hu, N.; Medrano-Guzman, R.; Herrera-Goepfert, R.; Humphrys, M.S.; Wang, L.; Wang, C.; Ding, T.; Ravel, J.; et al. Molecular Characterisation of the Human Stomach Microbiota in Gastric Cancer Patients. Front. Cell. Infect. Microbiol. 2017, 7, 302. [Google Scholar] [CrossRef] [PubMed]
- Seo, I.; Jha, B.K.; Suh, S.-I.; Suh, M.-H.; Baek, W.-K. Microbial Profile of the Stomach: Comparison between Normal Mucosa and Cancer Tissue in the Same Patient. J. Bacteriol. Virol. 2014, 44, 162. [Google Scholar] [CrossRef][Green Version]
- Chen, X.-H.; Wang, A.; Chu, A.-N.; Gong, Y.-H.; Yuan, Y. Mucosa-Associated Microbiota in Gastric Cancer Tissues Compared With Non-Cancer Tissues. Front. Microbiol. 2019, 10, 1261. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of Gastric Mucosal Microbiota across Different Stomach Microhabitats in a Cohort of 276 Patients with Gastric Cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xin, Y.; Zhou, J.; Tian, Z.; Liu, C.; Yu, X.; Meng, X.; Jiang, W.; Zhao, S.; Dong, Q. Gastric Mucosa-Associated Microbial Signatures of Early Gastric Cancer. Front. Microbiol. 2020, 11, 1548. [Google Scholar] [CrossRef] [PubMed]
- Correa, P.; Piazuelo, M.B. The Gastric Precancerous Cascade. J. Dig. Dis. 2012, 13, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Aviles-Jimenez, F.; Vazquez-Jimenez, F.; Medrano-Guzman, R.; Mantilla, A.; Torres, J. Stomach Microbiota Composition Varies between Patients with Non-Atrophic Gastritis and Patients with Intestinal Type of Gastric Cancer. Sci. Rep. 2014, 4, 4202. [Google Scholar] [CrossRef] [PubMed]
- Eun, C.S.; Kim, B.K.; Han, D.S.; Kim, S.Y.; Kim, K.M.; Choi, B.Y.; Song, K.S.; Kim, Y.S.; Kim, J.F. Differences in Gastric Mucosal Microbiota Profiling in Patients with Chronic Gastritis, Intestinal Metaplasia, and Gastric Cancer Using Pyrosequencing Methods. Helicobacter 2014, 19, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Li, T.H.; Qin, Y.; Sham, P.C.; Lau, K.S.; Chu, K.-M.; Leung, W.K. Alterations in Gastric Microbiota After H. Pylori Eradication and in Different Histological Stages of Gastric Carcinogenesis. Sci. Rep. 2017, 7, 44935. [Google Scholar] [CrossRef]
- He, C.; Peng, C.; Shu, X.; Wang, H.; Zhu, Z.; Ouyang, Y.; Yang, X.; Xie, C.; Hu, Y.; Li, N.; et al. Convergent Dysbiosis of Gastric Mucosa and Fluid Microbiome during Stomach Carcinogenesis. Gastric Cancer 2022, 25, 837–849. [Google Scholar] [CrossRef]
- Lopes, C.; Chaves, J.; Ortigão, R.; Dinis-Ribeiro, M.; Pereira, C. Gastric Cancer Detection by Non-blood-based Liquid Biopsies: A Systematic Review Looking into the Last Decade of Research. United Eur. Gastroenterol. J. 2023, 11, 114–130. [Google Scholar] [CrossRef]
- Hu, Y.L.; Pang, W.; Huang, Y.; Zhang, Y.; Zhang, C.J. The Gastric Microbiome Is Perturbed in Advanced Gastric Adenocarcinoma Identified Through Shotgun Metagenomics. Front. Cell. Infect. Microbiol. 2018, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.Y.; Tung, S.Y.; Pan, H.Y.; Yen, C.W.; Xu, H.W.; Lin, Y.J.; Deng, Y.F.; Hsu, W.T.; Wu, C.S.; Li, C. Increased Abundance of Clostridium and Fusobacterium in Gastric Microbiota of Patients with Gastric Cancer in Taiwan. Sci. Rep. 2018, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Seo, H.; Kang, C.S.; Shin, T.S.; Kim, J.W.; Park, J.M.; Kim, J.G.; Kim, Y.K. Dysbiotic Change in Gastric Microbiome and Its Functional Implication in Gastric Carcinogenesis. Sci. Rep. 2022, 12, 4285. [Google Scholar] [CrossRef] [PubMed]
- Nearing, J.T.; Douglas, G.M.; Hayes, M.G.; MacDonald, J.; Desai, D.K.; Allward, N.; Jones, C.M.A.; Wright, R.J.; Dhanani, A.S.; Comeau, A.M.; et al. Microbiome Differential Abundance Methods Produce Different Results across 38 Datasets. Nat. Commun. 2022, 13, 342. [Google Scholar] [CrossRef] [PubMed]
- Kameoka, S.; Motooka, D.; Watanabe, S.; Kubo, R.; Jung, N.; Midorikawa, Y.; Shinozaki, N.O.; Sawai, Y.; Takeda, A.K.; Nakamura, S. Benchmark of 16S rRNA Gene Amplicon Sequencing Using Japanese Gut Microbiome Data from the V1–V2 and V3–V4 Primer Sets. BMC Genom. 2021, 22, 527. [Google Scholar] [CrossRef]
- Liu, C.; Ng, S.K.; Ding, Y.; Lin, Y.; Liu, W.; Wong, S.H.; Sung, J.J.Y.; Yu, J. Meta-Analysis of Mucosal Microbiota Reveals Universal Microbial Signatures and Dysbiosis in Gastric Carcinogenesis. Oncogene 2022, 41, 3599–3610. [Google Scholar] [CrossRef]
- Guo, Y.; Cao, X.S.; Guo, G.Y.; Zhou, M.G.; Yu, B. Effect of Helicobacter Pylori Eradication on Human Gastric Microbiota: A Systematic Review and Meta-Analysis. Front. Cell. Infect. Microbiol. 2022, 12, 899248. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.M.; Kim, N.; Park, J.H.; Lee, D.H. Changes in Gastric Corpus Microbiota With Age and After Helicobacter Pylori Eradication: A Long-Term Follow-Up Study. Front. Microbiol. 2021, 11, 621879. [Google Scholar] [CrossRef]
- Sung, J.J.Y.; Coker, O.O.; Chu, E.; Szeto, C.H.; Luk, S.T.Y.; Lau, H.C.H.; Yu, J. Gastric Microbes Associated with Gastric Inflammation, Atrophy and Intestinal Metaplasia 1 Year after Helicobacter Pylori Eradication. Gut 2020, 69, 1572–1580. [Google Scholar] [CrossRef]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A Bacterial Driver–passenger Model for Colorectal Cancer: Beyond the Usual Suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).