
Environmental and Nutritional Parameters Modulating Genetic Expression for Virulence Factors of Clostridioides difficile

 and
and 
Abstract
1. Introduction
2. Overview of Genes Associated with Virulence of C. difficile
2.1. Production of Toxin A and Toxin B: tcdA and tcdB
2.2. Regulatory Genes for Toxin A and Toxin B: tcdR, tcdC and tcdE
2.3. Promoter Genes for Toxin A and Toxin B Production: ccpA, codY
2.4. Quorum Sensing Genes: agr1, agr2, luxS
2.5. Binary Toxin Genes: cdtA and cdtB
2.6. Adhesion Genes: cwp13, cwp66, cwp84, Etc
2.7. Biofilm Genes: spoOA
2.8. Motility Genes: fliC, fliE, filG
2.9. Genes Involved in Spore Germination: cspA, cspC, spoOA
3. Modifiable Parameters of C. difficile Gene Expression
3.1. Antibiotics
3.1.1. Antibiotics and Toxin A and Toxin B Production
3.1.2. Antibiotics and Adherence
3.1.3. Antibiotics and Biofilm Formation
3.1.4. Antibiotics and Spore Germination
3.2. Nutritional Factors
3.2.1. Amino Acids
3.2.2. Carbohydrates
3.2.3. Green Tea
3.2.4. Bile Acids
3.3. Environmental Stimuli
3.3.1. Stress
3.3.2. Temperature
3.3.3. pH
3.4. Viral Factors: Prophages
3.5. Microbial Factors: Probiotics
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Centers for Communicable Diseases (CDC). National and State Healthcare-Associated Infections Progress Report. 2020. Available online: https://www.cdc.gov/hai/data/portal/progress-report.html (accessed on 2 February 2023).
- Wiuff, C.; Banks, A.L.; Fitzpatrick, F.; Cottom, L. The need for European surveillance of CDI. In Updates on Clostridium difficile in Europe. Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2018; Volume 1050, pp. 13–25. [Google Scholar]
- Feuerstadt, P.; Boules, M.; Strong, L.; Dahdal, D.N.; Sacks, N.C.; Lang, K.; Nelson, W.W. Clinical complications in patients with primary and recurrent Clostridioides difficile infection: A real-world data analysis. SAGE Open Med. 2021, 9, 2050312120986733. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Goldstein, E.J.C.; Kullar, R. Microbiome-related and infection control approaches to primary and secondary prevention of Clostridioides difficile infections. Microorganisms 2023, 11, 1534. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Monaghan, T.; Yadegar, A.; Louie, T.; Kao, D. Insights into the evolving epidemiology of Clostridioides difficile infection and treatment: A global perspective. Antibiotics 2023, 12, 1141. [Google Scholar] [CrossRef] [PubMed]
- Buddle, J.E.; Fagan, R.P. Pathogenicity and virulence of Clostridioides difficile. Virulence 2023, 14, 2150452. [Google Scholar] [CrossRef] [PubMed]
- Aldeyab, M.A.; Devine, M.J.; Flanagan, P.; Mannion, M.; Craig, A.; Scott, M.G.; Harbarth, S.; Vernaz, N.; Davies, E.; Brazier, J.S.; et al. Multihospital outbreak of Clostridium difficile ribotype 027 infection: Epidemiology and analysis of control measures. Infect. Cont. Hosp. Epidemiol. 2011, 32, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Paquette, I.M.; Stewart, D.B. Clostridium difficile Infection. In The ASCRS Textbook of Colon and Rectal Surgery; Springer: Berlin/Heidelberg, Germany, 2022; pp. 879–891. [Google Scholar]
- Majumdar, A.; Govind, R. Regulation of Clostridioides difficile toxin production. Curr. Opin. Microbiol. 2022, 65, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Fatima, R.; Aziz, M. The hypervirulent strain of Clostridium difficile: NAP1/B1/027—A brief overview. Cureus 2019, 11, e3977. [Google Scholar] [CrossRef] [PubMed]
- Vaishnavi, C. Virulence factors associated with Clostridiodes difficile: An overview. J. Gastro Infect. 2021, 11, 25. [Google Scholar] [CrossRef]
- Saund, K.; Pirani, A.; Lacy, B.; Hanna, P.C.; Snitkin, E.S. Strain variation in Clostridioides difficile toxin activity associated with genomic variation at both PaLoc and non-PaLoc loci. bioRxiv 2021. [Google Scholar] [CrossRef]
- Shen, J.; Mehrotra, D.V.; Dorr, M.B.; Zeng, Z.; Li, J.; Xu, X.; Nickle, D.; Holzinger, E.R.; Chhibber, A.; Wilcox, M.H. Genetic association reveals protection against recurrence of Clostridium difficile infection with bezlotoxumab treatment. Msphere 2020, 5, e00232-20. [Google Scholar] [CrossRef]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; Di Masi, A. Clostridium difficile toxins A and B: Insights into pathogenic properties and extraintestinal effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Gunaratnam, S.; Millette, M.; McFarland, L.V.; DuPont, H.L.; Lacroix, M. Potential role of probiotics in reducing Clostridioides difficile virulence: Interference with quorum sensing systems. Microb. Pathog. 2021, 153, 104798. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, R.; Lacy, D.B. The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 2017, 41, 723–750. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.R.; Pike, C.M.; Parsons, R.J.; Rivera, A.J.; Foley, M.H.; McLaren, M.R.; Montgomery, S.A.; Theriot, C.M. Clostridioides difficile exploits toxin-mediated inflammation to alter the host nutritional landscape and exclude competitors from the gut microbiota. Nat. Commun. 2021, 12, 462. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Li, W.G.; Zhang, W.Z.; Wu, Y.; Lu, J.X. Molecular characterization of Clostridium difficile isolates in China from 2010 to 2015. Front. Microbiol. 2018, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Neumann-Schaal, M.; Jahn, D.; Schmidt-Hohagen, K. Metabolism the difficile way: The key to the success of the pathogen Clostridioides difficile. Front. Microbiol. 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Daou, N.; Wang, Y.; Levdikov, V.M.; Nandakumar, M.; Livny, J.; Bouillaut, L.; Blagova, E.; Zhang, K.; Belitsky, B.R.; Rhee, K. Impact of CodY protein on metabolism, sporulation and virulence in Clostridioides difficile ribotype 027. PLoS ONE 2019, 14, e0206896. [Google Scholar] [CrossRef] [PubMed]
- Dineen, S.S.; McBride, S.M.; Sonenshein, A.L. Integration of metabolism and virulence by Clostridium difficile CodY. J. Bacteriol. 2010, 192, 5350–5362. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, U.K.B.; Ballard, J.D. Autoinducing peptide-based quorum signaling systems in Clostridioides difficile. Curr. Opin. Microbiol. 2022, 65, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Boo, A.; Amaro, R.L.; Stan, G.B. Quorum sensing in synthetic biology: A review. Curr. Opin. Syst. Biol. 2021, 28, 100378. [Google Scholar] [CrossRef]
- Nag, M.; Lahiri, D.; Ghosh, A.; Das, D.; Ray, R.R. Quorum sensing. In Biofilm-Mediated Diseases: Causes and Controls; Springer: Singapore, 2021; pp. 21–45. [Google Scholar] [CrossRef]
- Darkoh, C.; Odo, C.; DuPont, H.L. Accessory gene regulator-1 locus is essential for virulence and pathogenesis of Clostridium difficile. Mbio 2016, 7, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Mawer, D.; Wilcox, M.H. Clostridium difficile: Biological therapies. Curr. Opin. Infect. Dis. 2013, 26, 454–460. [Google Scholar] [CrossRef]
- Young, M.; Leslie, J.L.; Madden, G.R.; Lyerly, D.M.; Carman, R.J.; Lyerly, M.W.; Stewart, D.B.; Abhyankar, M.M.; Petri, W.A., Jr. Binary toxin expression by Clostridioides difficile is associated with worse disease. Open For. Infect. Dis. 2022, 9, ofac001. [Google Scholar] [CrossRef] [PubMed]
- Monot, M.; Eckert, C.; Lemire, A.; Hamiot, A.; Dubois, T.; Tessier, C.; Dumoulard, B.; Hamel, B.; Petit, A.; Lalande, V. Clostridium difficile: New insights into the evolution of the pathogenicity locus. Sci. Rep. 2015, 5, 15023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Rao, F.; Chen, Z.; Cheng, Y.; Zhang, Q.; Zhang, J.; Guan, Z.; He, Y.; Yu, W.; Cui, G. The cwp66 gene affects cell adhesion, stress tolerance, and antibiotic resistance in Clostridioides difficile. Microbiol. Spect. 2022, 10, e02704-21. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, Y.D.; Dupuy, B. The blueprint for building a biofilm the Clostridioides difficile way. Curr. Opin. Microbiol. 2022, 66, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Taggart, M.G.; Snelling, W.J.; Naughton, P.J.; La Ragione, R.M.; Dooley, J.S.; Ternan, N.G. Biofilm regulation in Clostridioides difficile: Novel systems linked to hypervirulence. PLoS Path 2021, 17, e1009817. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.B.; Emerson, J.E.; De la Riva, L.; Fagan, R.P.; Fairweather, N.F. The Clostridium difficile cell wall protein CwpV is antigenically variable between strains, but exhibits conserved aggregation-promoting function. PLoS Path 2011, 7, e1002024. [Google Scholar] [CrossRef] [PubMed]
- Vuotto, C.; Donelli, G.; Buckley, A.; Chilton, C. Clostridium difficile biofilm. In Updates on Clostridium difficile in Europe: Advances in Microbiology, Infectious Diseases and Public Health; Springer: Berlin/Heidelberg, Germany, 2018; Volume 8, pp. 97–115. [Google Scholar]
- Frost, L.R.; Cheng, J.K.; Unnikrishnan, M. Clostridioides difficile biofilms: A mechanism of persistence in the gut? PLoS Path 2021, 17, e1009348. [Google Scholar] [CrossRef] [PubMed]
- Arora, G.; Sajid, A.; Virmani, R.; Singhal, A.; Kumar, C.S.; Dhasmana, N.; Khanna, T.; Maji, A.; Misra, R.; Molle, V. Ser/Thr protein kinase PrkC-mediated regulation of GroEL is critical for biofilm formation in Bacillus anthracis. NPJ Biofilms Microbiomes 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Dapa, T.; Unnikrishnan, M. Biofilm formation by Clostridium difficile. Gut Microbiol. 2013, 4, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, O.A. Characterisation of Reduced Susceptibility to Metronidazole in Epidemic Clostridium difficile Clinical Isolates. Ph.D. Thesis, University of Hertfordshire, Hatfield, UK, 2018. [Google Scholar] [CrossRef]
- Slater, R.T.; Frost, L.R.; Jossi, S.E.; Millard, A.D.; Unnikrishnan, M. Clostridioides difficile LuxS mediates inter-bacterial interactions within biofilms. Sci. Rep. 2019, 9, 9903. [Google Scholar] [CrossRef] [PubMed]
- Aubry, A.; Hussack, G.; Chen, W.; KuoLee, R.; Twine, S.M.; Fulton, K.M.; Foote, S.; Carrillo, C.D.; Tanha, J.; Logan, S.M. Modulation of toxin production by the flagellar regulon in Clostridium difficile. Infect. Immun. 2012, 80, 3521–3532. [Google Scholar] [CrossRef] [PubMed]
- Batah, J.; Kansau, I. Intestinal epithelial cell response to Clostridium difficile flagella. In Clostridium difficile: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2016; pp. 103–116. [Google Scholar]
- Dingle, T.; Mulvey, G.L.; Armstrong, G.D. Mutagenic analysis of the Clostridium difficile flagellar proteins, FliC and FliD, and their contribution to virulence in hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.N.; Willams, C.L.; Pareek, N.; McBride, S.M.; Tamayo, R. c-di-GMP inhibits early sporulation in Clostridioides difficile. Msphere 2021, 6, e00919–e00921. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Ozen, M.; Dinleyici, E.C.; Goh, S. Comparison of pediatric and adult antibiotic-associated diarrhea and Clostridium difficile infections. World J. Gastroenterol. 2016, 22, 3078–3104. [Google Scholar] [CrossRef] [PubMed]
- Gerber, M.; Walch, C.; Löffler, B.; Tischendorf, K.; Reischl, U.; Ackermann, G. Effect of sub-MIC concentrations of metronidazole, vancomycin, clindamycin and linezolid on toxin gene transcription and production in Clostridium difficile. J. Med. Microbiol. 2008, 57, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Mansouri, S.; Nakhaee, N.; Sarafzadeh, F.; Zarandi, E.R. Toxin A and B genes expression of Clostridium difficile in the sub-minimum inhibitory concentration of clindamycin, vancomycin and in combination with ceftazidime. Iran. J. Microbiol. 2020, 12, 18. [Google Scholar] [CrossRef]
- Aldape, M.J.; Packham, A.E.; Nute, D.W.; Bryant, A.E.; Stevens, D.L. Effects of ciprofloxacin on the expression and production of exotoxins by Clostridium difficile. J. Med. Microbiol. 2013, 62 Pt 5, 741. [Google Scholar] [CrossRef] [PubMed]
- Aldape, M.J.; Heeney, D.D.; Bryant, A.E.; Stevens, D.L. Tigecycline suppresses toxin A and B production and sporulation in Clostridium difficile. J. Antimicrob. Chemother. 2015, 70, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Babakhani, F.; Bouillaut, L.; Sears, P.; Sims, C.; Gomez, A.; Sonenshein, A.L. Fidaxomicin inhibits toxin production in Clostridium difficile. J. Antimicrob. Chemother. 2013, 68, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Deneve, C.; Delomenie, C.; Barc, M.C.; Collignon, A.; Janoir, C. Antibiotics involved in Clostridium difficile-associated disease increase colonization factor gene expression. J. Med. Microbiol. 2008, 57, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Denève, C.; Bouttier, S.; Dupuy, B.; Barbut, F.; Collignon, A.; Janoir, C. Effects of subinhibitory concentrations of antibiotics on colonization factor expression by moxifloxacin-susceptible and moxifloxacin-resistant Clostridium difficile strains. Antimicrob. Agents Chemother. 2009, 53, 5155–5162. [Google Scholar] [CrossRef] [PubMed]
- Mikamo, H.; Tateda, K.; Yanagihara, K.; Kusachi, S.; Takesue, Y.; Miki, T.; Oizumi, Y.; Gamo, K.; Hashimoto, A.; Toyoshima, J. Efficacy and safety of fidaxomicin for the treatment of Clostridioides (Clostridium) difficile infection in a randomized, double-blind, comparative Phase III study in Japan. J. Infect. Chemother. 2018, 24, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Yamaguchi, T.; Ishii, Y.; Chono, K.; Tateda, K. Inhibitory effect of fidaxomicin on biofilm formation in Clostridioides difficile. J. Infect. Chemother. 2020, 26, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.D.; Otto, A.; Berges, M.; Biedendieck, R.; Michel, A.M.; Becher, D.; Jahn, D.; Neumann-Schaal, M. Metabolic reprogramming of Clostridioides difficile during the stationary phase with the induction of toxin production. Front. Microbiol. 2018, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- Dubois, T.; Dancer-Thibonnier, M.; Monot, M.; Hamiot, A.; Bouillaut, L.; Soutourina, O.; Martin-Verstraete, I.; Dupuy, B. Control of Clostridium difficile physiopathology in response to cysteine availability. Infect. Immun. 2016, 84, 2389–2405. [Google Scholar] [CrossRef] [PubMed]
- Sommermeyer, H.; Piątek, J. Pathophysiology of C. difficile. In Clostridioides difficile: Infections, Risk Factors, Prevention and Treatment; Springer International Publisher: New York, NY, USA, 2021; pp. 19–33. [Google Scholar]
- Gu, H.; Shi, K.; Liao, Z.; Qi, H.; Chen, S.; Wang, H.; Li, S.; Ma, Y.; Wang, J. Time-resolved transcriptome analysis of Clostridium difficile R20291 response to cysteine. Microbiol. Res. 2018, 215, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, K.L.; Wetzel, D.; Jones, J.B.; Woods, E.C.; McBride, S.M. Ethanolamine is a valuable nutrient source that impacts Clostridium difficile pathogenesis. Environ. Microbiol. 2018, 20, 1419–1435. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Martin-Verstraete, I.; Dupuy, B. CcpA-mediated repression of Clostridium difficile toxin gene expression. Mol. Microbiol. 2011, 79, 882–899. [Google Scholar] [CrossRef]
- Antunes, A.; Camiade, E.; Monot, M.; Courtois, E.; Barbut, F.; Sernova, N.V.; Rodionov, D.A.; Martin-Verstraete, I.; Dupuy, B. Global transcriptional control by glucose and carbon regulator CcpA in Clostridium difficile. Nucl. Acids Res. 2012, 40, 10701–10718. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Oh, S.; Song, M.; Hong, Y.S.; Park, S.; Park, D.J.; Griffiths, M.W.; Oh, S. Inhibitory effect of epigallocatechin gallate on the virulence of Clostridium difficile PCR ribotype 027. J. Food Sci. 2015, 80, M2925–M2931. [Google Scholar] [CrossRef] [PubMed]
- Budi, N.; Safdar, N.; Rose, W.E. Treatment issues in recurrent Clostridioides difficile and the possible role of germinants. FEMS Microbes 2020, 1, xtaa001. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.J.; Brunner, A.; Lewis, J.; Ford, C.D.; Lopansri, B.K. Repurposing an old drug for a new epidemic: Ursodeoxycholic acid to prevent recurrent Clostridioides difficile infection. Clin. Infect. Dis. 2019, 68, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Rahal, A.; Sohal, J.S.; Gupta, V.K. Bacterial stress response: Understanding the molecular mechanics to identify possible therapeutic targets. Expert Rev. Anti-Infect. Ther. 2021, 19, 121–127. [Google Scholar] [CrossRef] [PubMed]
- De Bruijn, F.J. Introduction. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; De Bruijn, F.J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; Volume 1, pp. 1–2. [Google Scholar]
- Willing, S.E.; Richards, E.J.; Sempere, L.; Dale, A.G.; Cutting, S.M.; Fairweather, N.F. Increased toxin expression in a Clostridium difficile mfd mutant. BMC Microbiol. 2015, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Tilly, K. Temperature-regulated expression of bacterial virulence genes. Microb. Infect. 2000, 2, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Dupuy, B.; Mukherjee, K.; Norin, E.; Burman, L.G.; Akerlund, T. Expression of Clostridium difficile toxins A and B and their sigma factor TcdD is controlled by temperature. Infect. Immun. 2003, 71, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.; Hegarty, J.P. Correlation between virulence gene expression and proton pump inhibitors and ambient pH in Clostridium difficile: Results of an in vitro study. J. Med. Microbiol. 2013, 62, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, S.A.; Merenstein, D.; Fraser, C.M.; Marco, M.L. Molecular mechanisms of probiotic prevention of antibiotic-associated diarrhea. Curr. Opin. Biotechnol. 2020, 61, 226–234. [Google Scholar] [CrossRef]
- Schroven, K.; Aertsen, A.; Lavigne, R. Bacteriophages as drivers of bacterial virulence and their potential for biotechnological exploitation. FEMS Microbiol. Rev. 2021, 45, fuaa041. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C. Bacteriophages contribute to shaping Clostridioides (Clostridium) difficile species. Front. Microbiol. 2018, 9, 2033. [Google Scholar] [CrossRef] [PubMed]
- Govind, R.; Vediyappan, G.; Rolfe, R.D.; Dupuy, B.; Fralick, J.A. Bacteriophage-mediated toxin gene regulation in Clostridium difficile. J. Virol. 2009, 83, 12037–12045. [Google Scholar] [CrossRef]
- Riedel, T.; Wittmann, J.; Bunk, B.; Schober, I.; Spröer, C.; Gronow, S.; Overmann, J. A Clostridioides difficile bacteriophage genome encodes functional binary toxin-associated genes. J. Biotechnol. 2017, 250, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Kropinski, A.M.; Clokie, M.R. What does the talking?: Quorum sensing signalling genes discovered in a bacteriophage genome. PLoS ONE 2014, 9, e85131. [Google Scholar] [CrossRef] [PubMed]
- Su, G.L.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA clinical practice guidelines on the role of probiotics in the management of gastrointestinal disorders. Gastroenterology 2020, 159, 697–705. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Ship, N.; Auclair, J.; Millette, M. Primary prevention of Clostridium difficile infections with a specific probiotic combining Lactobacillus acidophilus, L. casei and L. rhamnosus strains: Assessing the evidence. J. Hosp. Infect. 2018, 99, 443–452. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Strain-specificity and disease-specificity of probiotic efficacy: A systematic review and meta-analysis. Front. Med. 2018, 5, 124. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Mora, D.; Gschwender, M.; Popp, K. Randomised clinical trial: Bifidobacterium bifidum MIMBb75 significantly alleviates irritable bowel syndrome and improves quality of life—A double-blind, placebo-controlled study. Aliment. Pharmacol. Ther. 2011, 33, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Sniffen, J.C.; McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Choosing an appropriate probiotic product for your patient: An evidence-based practical guide. PLoS ONE 2018, 13, e0209205. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Oh, S.; Griffiths, M. Lactobacillus acidophilus modulates the virulence of Clostridium difficile. J. Dairy Sci. 2014, 97, 4745–4758. [Google Scholar] [CrossRef] [PubMed]
- Najarian, A.; Sharif, S.; Griffiths, M. Evaluation of protective effect of Lactobacillus acidophilus La-5 on toxicity and colonization of Clostridium difficile in human epithelial cells in vitro. Anaerobe 2019, 55, 142–151. [Google Scholar] [CrossRef]
- Yong, C.; Lim, J.; Kim, B.K.; Park, D.J.; Oh, S. Suppressive effect of Lactobacillus fermentum Lim2 on Clostridioides difficile 027 toxin production. Lett. Appl. Microbiol. 2019, 68, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Masset, Z.; Gunaratnam, S.; Millette, M.; McFarland, L.V.; Lacroix, M. Transcriptome analysis of the Clostridioides difficile response to a specific lactobacilli probiotic formulation: Explanations for its mechanisms of action. J. Appl. Microbiol. 2023, 134, lxad047. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, H. Antibacterial activity of Bifidobacterium breve against Clostridioides difficile. Front. Cell Infect. Microbiol. 2019, 9, 288. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Schmidt, D.; Liu, W.; Li, S.; Shi, L.; Sheng, J.; Chen, K.; Yu, H.; Tremblay, J.M.; Chen, X. A novel multivalent, single-domain antibody targeting TcdA and TcdB prevents fulminant Clostridium difficile infection in mice. J. Infect. Dis. 2014, 210, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, H. Evaluation of the therapeutic effect and dose–effect of Bifidobacterium breve on the primary Clostridioides difficile infected mice. Appl. Microbiol. Biotechnol. 2021, 105, 9243–9260. [Google Scholar] [CrossRef] [PubMed]
- Lamm-Schmidt, V.; Fuchs, M.; Sulzer, J.; Gerovac, M.; Hör, J.; Dersch, P.; Vogel, J.; Faber, F. Grad-seq identifies KhpB as a global RNA-binding protein in Clostridioides difficile that regulates toxin production. Microlife 2021, 2, uqab004. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M.; Lamm-Schmidt, V.; Sulzer, J.; Ponath, F.; Jenniches, L.; Kirk, J.A.; Fagan, R.P.; Barquist, L.; Vogel, J.; Faber, F. An RNA-centric global view of Clostridioides difficile reveals broad activity of Hfq in a clinically important gram-positive bacterium. Proc. Nat. Acad. Sci. USA 2021, 118, e2103579118. [Google Scholar] [CrossRef] [PubMed]
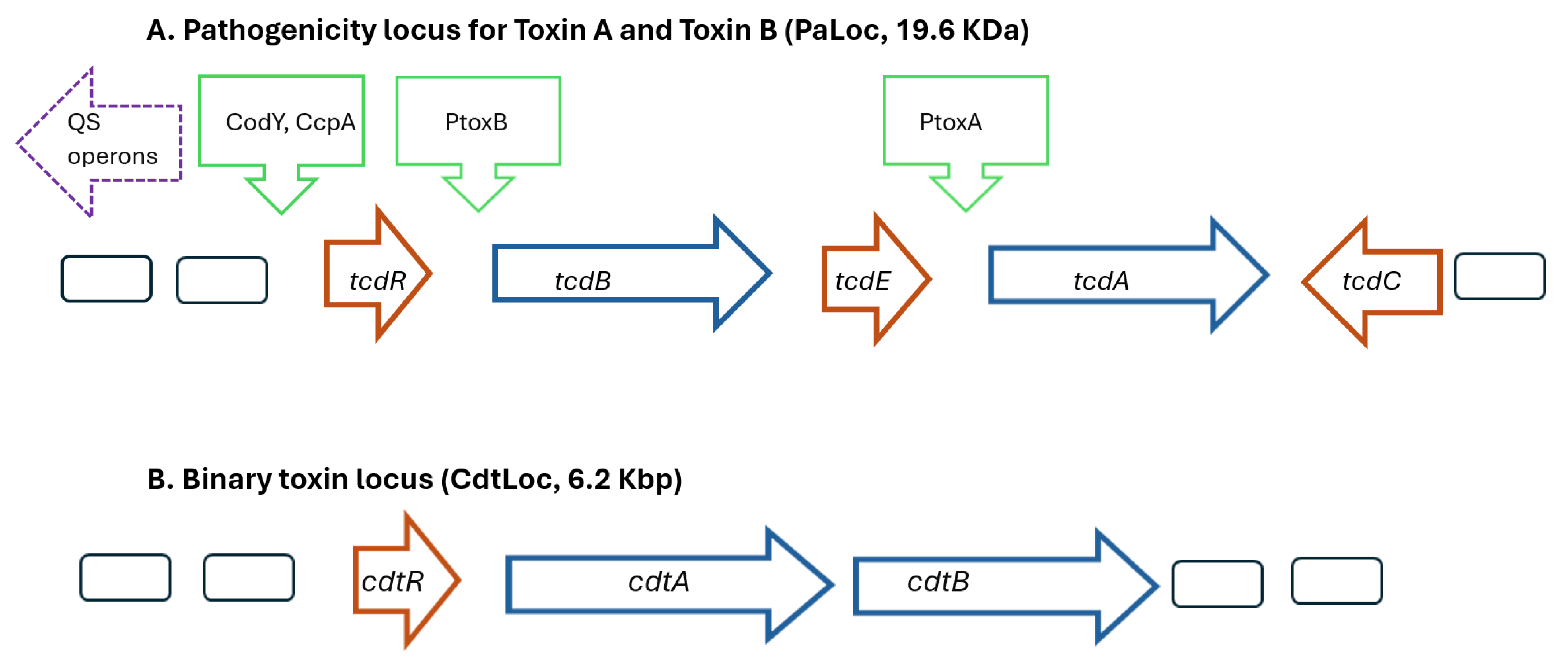

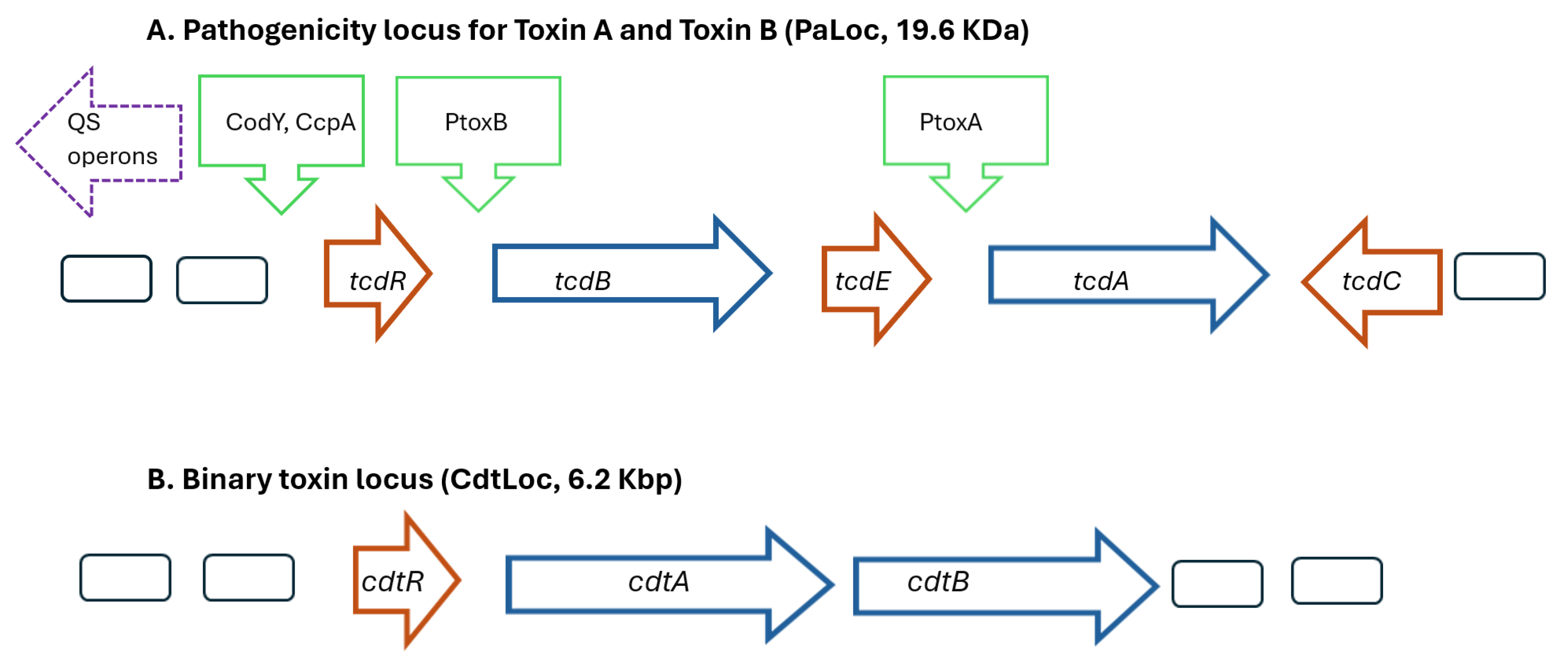{kind=link}
| Genes | Codes for | Roles | Factors Impacting Genetic Expression |
|---|---|---|---|
| tcdA tcdB | Toxin A (308 KDa) Toxin B (270 KDa) | enterotoxin and cytotoxins, inactivates Rho GTPases, resulting in actin filament disorganization, loss of cytoskeleton results in enterocyte collapse, opening tight junctions, with fluid secretion and diarrhea; also promotes inflammatory response | antibiotics, amino acids (cysteine), carbohydrates, green tea components, stress, temperature, prophage, probiotics |
| tcdR | RNA polymerase sigma factor (555 bp) | positive regulator for Toxins A and B | |
| tcdE | holin-like protein (501 bp) | positive regulator that facilitates extracellular release of toxins A and B via lytic action | |
| tcdC | anti-sigma factor (699 bp) | negative regulator for Toxin A and B production by disruption of TcdR | |
| codY | promoter for transcription (1113 bp) | binds to tcdR | nutritional (butyrate) |
| ccpA | promoter for transcription (831 bp) | binds to tcdR | nutritional (glucose) |
| agr1, agr2 | intraspecies accessory gene regulators (1500 bp, 500 bp) | QS pathway | nutritional (green tea), probiotics |
| luxS | interspecies gene regulatory (552 bp) | QS pathway | probiotics |
| cdtA cdtB | binary toxins (CDT), CdtA is an ADP-ribosyltransferase (1785 bp), CdtB is a polypeptide (1956 bp) | CdtA depolymerizes actin cytoskeleton, CdtB binds to host cells and transports CdtA into host cells | prophages, probiotics |
| cdtR | response regulator for CDT (747 bp) | positive regulator cdtA/cdtB | |
| cwp13, cwp66, cwp84, cwpV, etc. | cell wall proteins (2982 bp, 3168 bp, 2040 bp, etc.) | adhesion on S-layer of C. difficile cell wall | antibiotics, stress, probiotics |
| slpA | major S-layer constituent (885 bp) | cellular structure | antibiotics, stress, probiotics |
| spoOA | protein (1500 bp) | biofilm formation | antibiotics, carbohydrates |
| fliC, fliE, fliG | flagellar proteins (1155 bp, 1053 bp, 1317 bp) | motility and toxin production | antibiotics, probiotics |
| cspA, cspC, spoOA | regulatory genes (216 bp, 168 bp, 1509 bp) | spore germination, bile acid receptor | antibiotics, bile acids, environmental pH, probiotics |
| sigL | sigma factor regulator (426 bp) | downregulates Toxin A and B production | nutrition (cysteine) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masset, Z.; Gunaratnam, S.; Millette, M.; McFarland, L.V.; Lacroix, M. Environmental and Nutritional Parameters Modulating Genetic Expression for Virulence Factors of Clostridioides difficile. Antibiotics 2024, 13, 365. https://doi.org/10.3390/antibiotics13040365
Masset Z, Gunaratnam S, Millette M, McFarland LV, Lacroix M. Environmental and Nutritional Parameters Modulating Genetic Expression for Virulence Factors of Clostridioides difficile. Antibiotics. 2024; 13(4):365. https://doi.org/10.3390/antibiotics13040365
Chicago/Turabian StyleMasset, Zoe, Sathursha Gunaratnam, Mathieu Millette, Lynne V. McFarland, and Monique Lacroix. 2024. "Environmental and Nutritional Parameters Modulating Genetic Expression for Virulence Factors of Clostridioides difficile" Antibiotics 13, no. 4: 365. https://doi.org/10.3390/antibiotics13040365
APA StyleMasset, Z., Gunaratnam, S., Millette, M., McFarland, L. V., & Lacroix, M. (2024). Environmental and Nutritional Parameters Modulating Genetic Expression for Virulence Factors of Clostridioides difficile. Antibiotics, 13(4), 365. https://doi.org/10.3390/antibiotics13040365