
Assessment of Colistin Heteroresistance among Multidrug-Resistant Klebsiella pneumoniae Isolated from Intensive Care Patients in Europe

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample and Isolate Collection
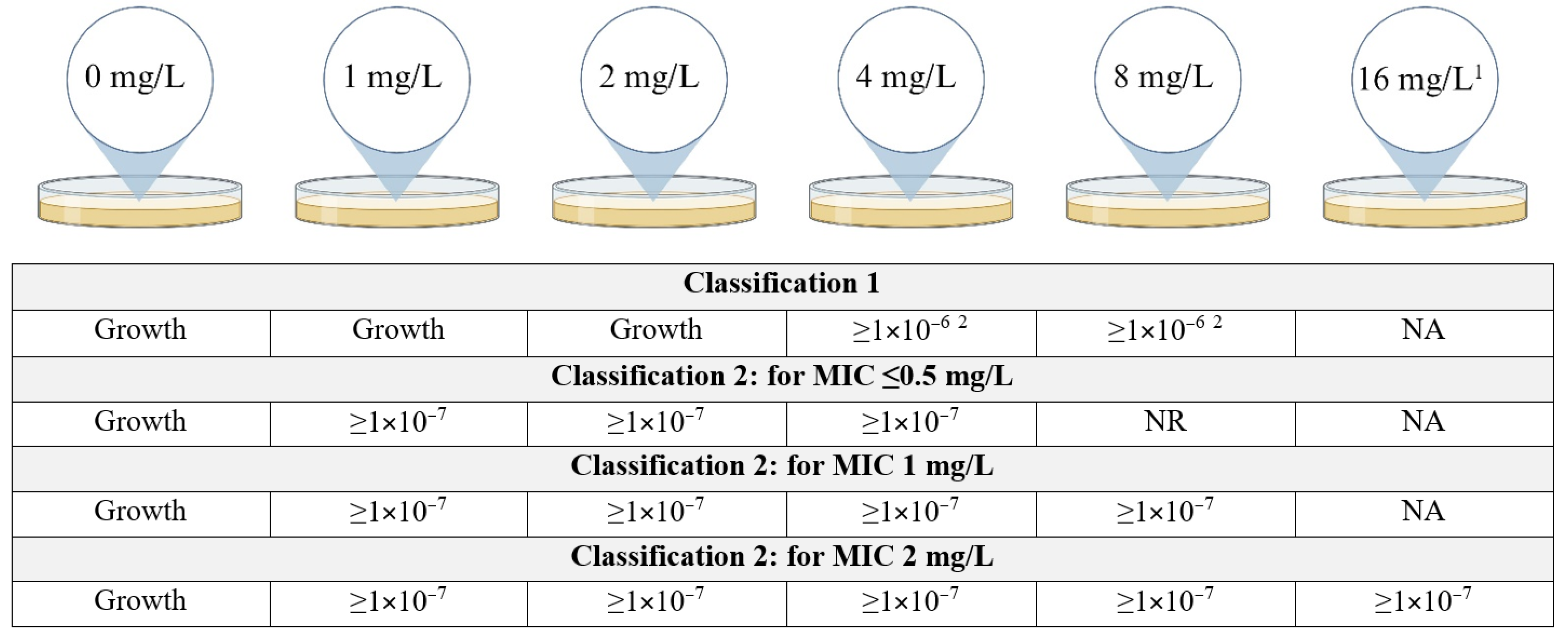
2.2. MIC Determination
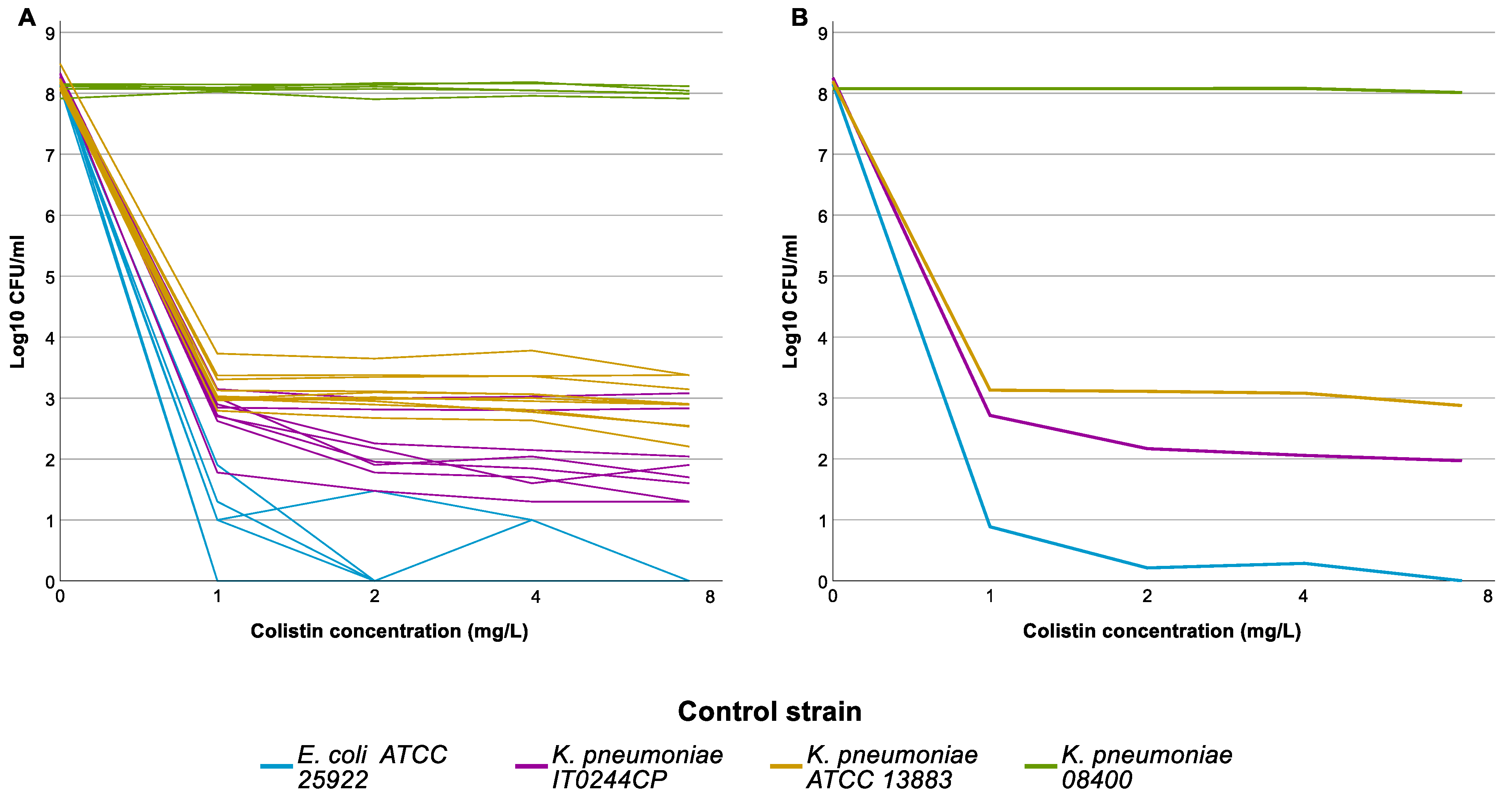
2.3. Population Analysis Profiling (PAP) Assay
2.4. Whole-Genome Sequencing
2.5. Statistical Analysis
3. Results
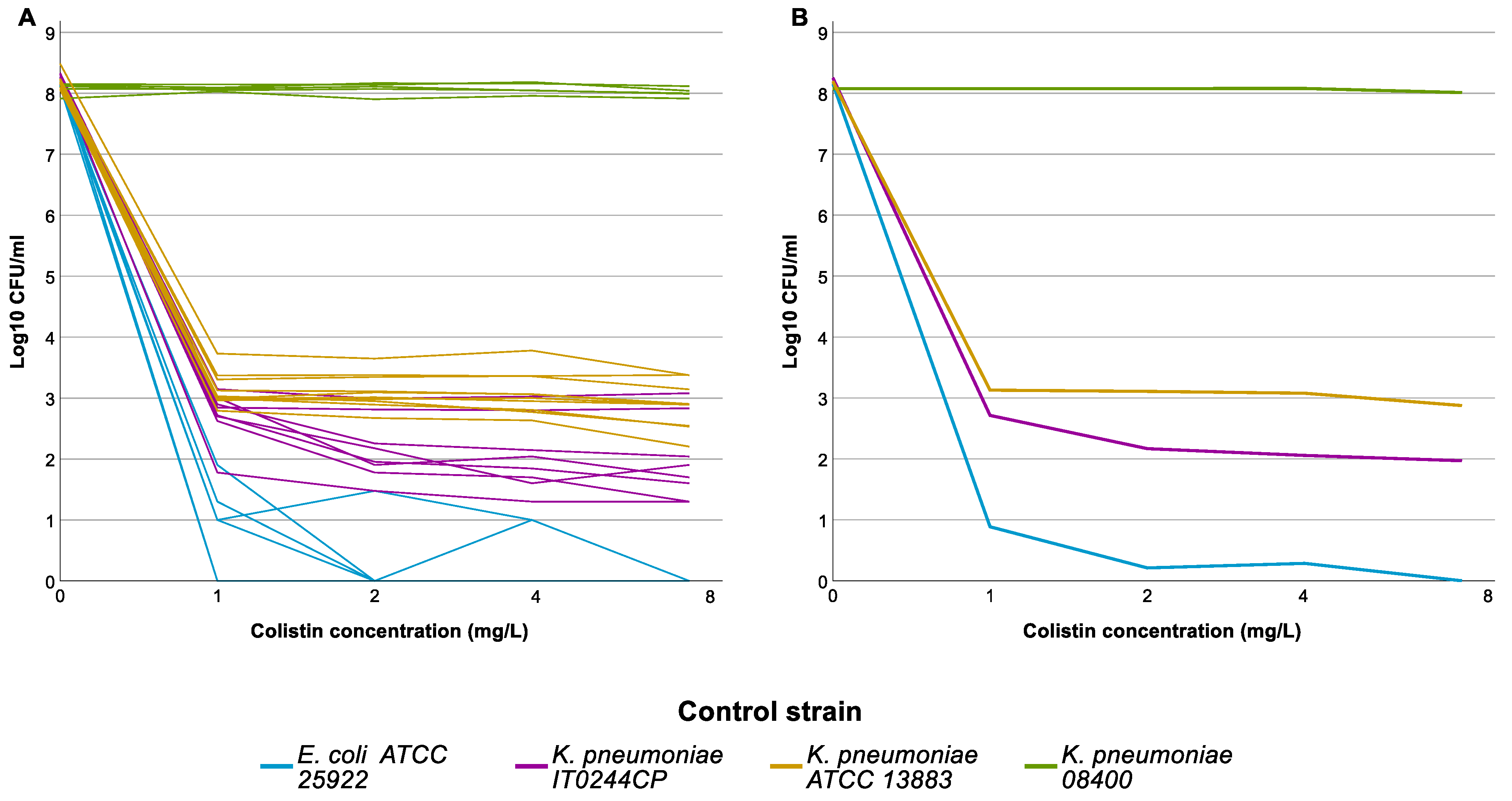
3.1. Population Analysis Profiling (PAP)
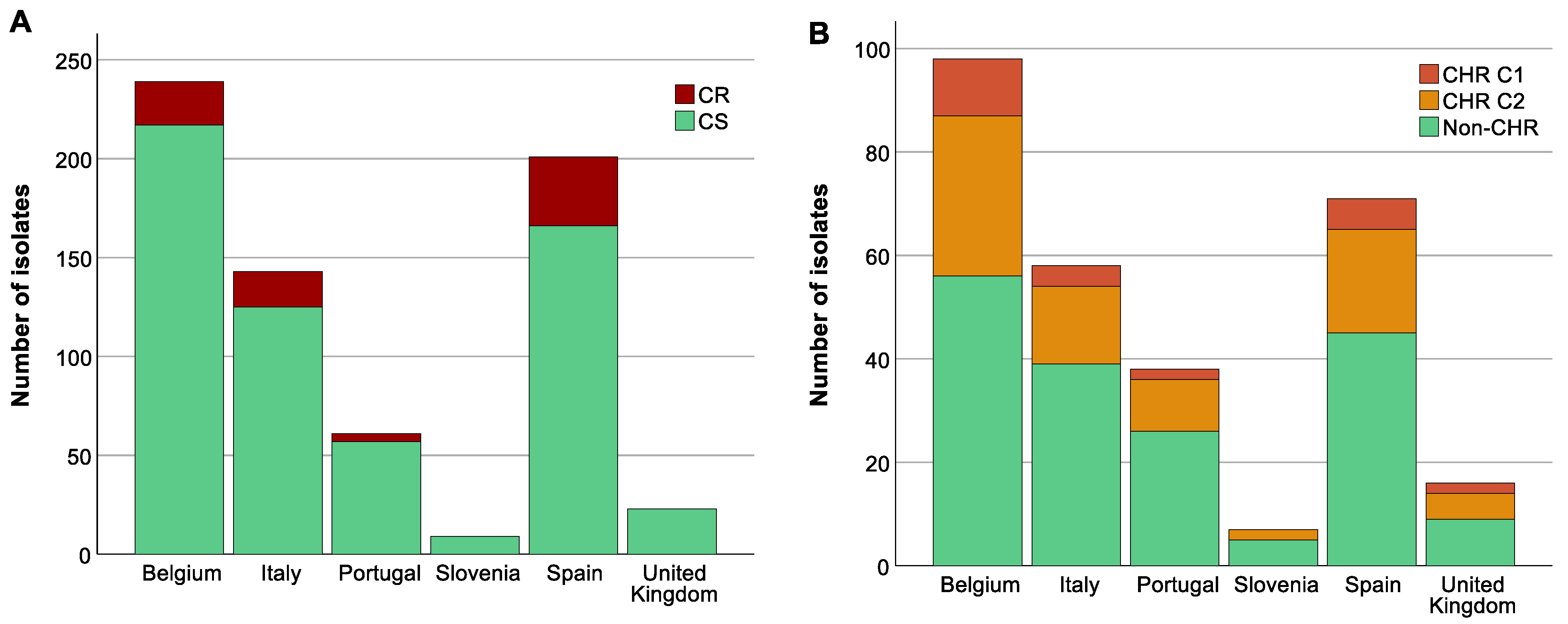
3.2. High Prevalence of Multi-Drug-Resistant and Carbapenemase-Producing Isolates
3.3. Analysis of Association between Colistin-Resistant K. pneumoniae, ST, O-Antigen Type and K-Antigen Type
3.4. A Higher Proportion of CR among Specific STs Was Not Reflected in CHR Proportions
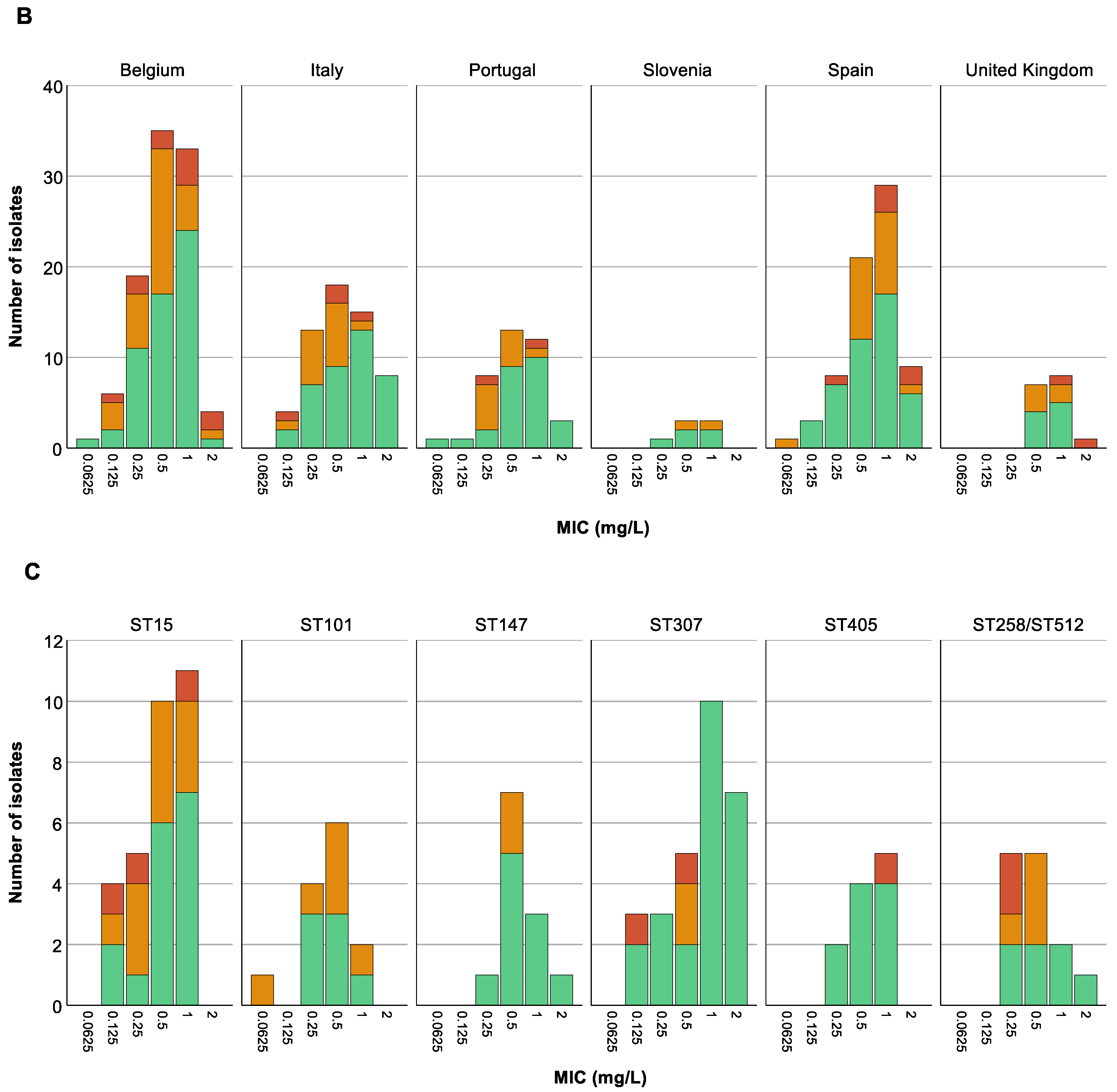
3.5. Analysis of Association between Colistin (Hetero)Resistant K. pneumoniae, Country, Intervention Strategy, and MIC-Value
3.6. Colistin Resistance Mechanisms in Resistant Subpopulations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roch, M.; Sierra, R.; Andrey, D.O. Antibiotic heteroresistance in ESKAPE pathogens, from bench to bedside. Clin. Microbiol. Infect. 2022, 29, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Della Rocca, M.T.; Foglia, F.; Crudele, V.; Greco, G.; De Filippis, A.; Franci, G.; Finamore, E.; Galdiero, M. Antimicrobial resistance changing trends of Klebsiella pneumoniae isolated over the last 5 years. New Microbiol. 2022, 45, 338–343. [Google Scholar] [PubMed]
- Mohd Asri, N.A.; Ahmad, S.; Mohamud, R.; Mohd Hanafi, N.; Mohd Zaidi, N.F.; Irekeola, A.A.; Shueb, R.H.; Yee, L.C.; Mohd Noor, N.; Mustafa, F.H.; et al. Global Prevalence of Nosocomial Multidrug-Resistant Klebsiella pneumoniae: A Systematic Review and Meta-Analysis. Antibiotics 2021, 10, 1508. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.G.; Earley, M.; Chen, L.; Hanson, B.M.; Yu, Y.S.; Liu, Z.Y.; Salcedo, S.; Cober, E.; Li, L.J.; Kanj, S.S.; et al. Clinical outcomes and bacterial characteristics of carbapenem-resistant complex among patients from different global regions (CRACKLE-2): A prospective, multicentre, cohort study. Lancet Infect. Dis. 2022, 22, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Ah, Y.M.; Kim, A.J.; Lee, J.Y. Colistin resistance in Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2014, 44, 8–15. [Google Scholar] [CrossRef]
- Ahmed, M.A.G.E.S.; Zhong, L.L.; Shen, C.; Yang, Y.Q.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Band, V.I.; Satola, S.W.; Burd, E.M.; Farley, M.M.; Jacob, J.T.; Weiss, D.S. Carbapenem-Resistant Klebsiella pneumoniae Exhibiting Clinically Undetected Colistin Heteroresistance Leads to Treatment Failure in a Murine Model of Infection. mBio 2018, 9, e02448-17. [Google Scholar] [CrossRef]
- Moosavian, M.; Shoja, S.; Nashibi, R.; Ebrahimi, N.; Tabatabaiefar, M.A.; Rostami, S.; Peymani, A. Post Neurosurgical Meningitis due to Colistin Heteroresistant Acinetobacter baumannii. Jundishapur J. Microbiol. 2014, 7, e12287. [Google Scholar] [CrossRef]
- Andersson, D.I.; Nicoloff, H.; Hjort, K. Mechanisms and clinical relevance of bacterial heteroresistance. Nat. Rev. Microbiol. 2019, 17, 479–496. [Google Scholar] [CrossRef]
- Stojowska-Swedrzynska, K.; Lupkowska, A.; Kuczynska-Wisnik, D.; Laskowska, E. Antibiotic Heteroresistance in Klebsiella pneumoniae. Int. J. Mol. Sci. 2021, 23, 449. [Google Scholar] [CrossRef] [PubMed]
- Dewachter, L.; Fauvart, M.; Michiels, J. Bacterial Heterogeneity and Antibiotic Survival: Understanding and Combatting Persistence and Heteroresistance. Mol. Cell 2019, 76, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Wittekamp, B.H.; Plantinga, N.L.; Cooper, B.; Lopez-Contreras, J.; Coll, P.; Mancebo, J.; Wise, M.P.; Morgan, M.P.G.; Depuydt, P.; Boelens, J.; et al. Decontamination Strategies and Bloodstream Infections With Antibiotic-Resistant Microorganisms in Ventilated Patients A Randomized Clinical Trial. J. Am. Med. Assoc. 2018, 320, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Anantharajah, A.; Glupczynski, Y.; Hoebeke, M.; Bogaerts, P.; Declercq, P.; Denis, O.; Descy, J.; Flore, K.; Magerman, K.; Rodriguez-Villalobos, H.; et al. Multicenter study of automated systems for colistin susceptibility testing. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Richter, S.S.; Karichu, J.; Otiso, J.; Van Heule, H.; Keller, G.; Cober, E.; Rojas, L.J.; Hujer, A.M.; Hujer, K.M.; Marshall, S.; et al. Evaluation of Sensititre Broth Microdilution Plate for determining the susceptibility of carbapenem-resistant Klebsiella pneumoniae to polymyxins. Diagn. Microbiol. Infect. Dis. 2018, 91, 89–92. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters; 2023. Available online: https://www.eucast.org/ (accessed on 15 December 2023).
- Rajakani, S.G.; Xavier, B.B.; Sey, A.; Mariem, E.; Lammens, C.; Goossens, H.; Glupczynski, Y.; Malhotra-Kumar, S. Insight into Antibiotic Synergy Combinations for Eliminating Colistin Heteroresistant. Genes 2023, 14, 1426. [Google Scholar] [CrossRef] [PubMed]
- Band, V.I.; Hufnagel, D.A.; Jaggavarapu, S.; Sherman, E.X.; Wozniak, J.E.; Satola, S.W.; Farley, M.M.; Jacob, J.T.; Burd, E.M.; Weiss, D.S. Antibiotic combinations that exploit heteroresistance to multiple drugs effectively control infection. Nat. Microbiol. 2019, 4, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Xavier, B.B.; Mysara, M.; Bolzan, M.; Ribeiro-Goncalves, B.; Alako, B.T.F.; Harrison, P.; Lammens, C.; Kumar-Singh, S.; Goossens, H.; Carrico, J.A.; et al. BacPipe: A Rapid, User-Friendly Whole-Genome Sequencing Pipeline for Clinical Diagnostic Bacteriology. iScience 2020, 23, 100769. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Juhasz, E.; Ivan, M.; Pinter, E.; Pongracz, J.; Kristof, K. Colistin resistance among blood culture isolates at a tertiary care centre in Hungary. J. Glob. Antimicrob. Resist. 2017, 11, 167–170. [Google Scholar] [CrossRef]
- Sherman, E.X.; Wozniak, J.E.; Weiss, D.S. Methods to Evaluate Colistin Heteroresistance in Acinetobacter baumannii. Methods Mol. Biol. 2019, 1946, 39–50. [Google Scholar] [CrossRef]
- Morales-Leon, F.; Lima, C.A.; Gonzalez-Rocha, G.; Opazo-Capurro, A.; Bello-Toledo, H. Colistin Heteroresistance among Extended Spectrum beta-lactamases-Producing Klebsiella pneumoniae. Microorganisms 2020, 8, 1279. [Google Scholar] [CrossRef]
- Band, V.I.; Satola, S.W.; Smith, R.D.; Hufnagel, D.A.; Bower, C.; Conley, A.B.; Rishishwar, L.; Dale, S.E.; Hardy, D.J.; Vargas, R.L.; et al. Colistin Heteroresistance Is Largely Undetected among Carbapenem-Resistant Enterobacterales in the United States. mBio 2021, 12, e02881-20. [Google Scholar] [CrossRef]
- Kon, H.; Hameir, A.; Nutman, A.; Temkin, E.; Keren Paz, A.; Lellouche, J.; Schwartz, D.; Weiss, D.S.; Kaye, K.S.; Daikos, G.L.; et al. Prevalence and Clinical Consequences of Colistin Heteroresistance and Evolution into Full Resistance in Carbapenem-Resistant Acinetobacter baumannii. Microbiol. Spectr. 2023, 11, e05093-22. [Google Scholar] [CrossRef]
- Howard-Anderson, J.; Davis, M.; Page, A.M.; Bower, C.W.; Smith, G.; Jacob, J.T.; Andersson, D.I.; Weiss, D.S.; Satola, S.W. Prevalence of colistin heteroresistance in carbapenem-resistant Pseudomonas aeruginosa and association with clinical outcomes in patients: An observational study. J. Antimicrob. Chemother. 2022, 77, 793–798. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention Control. Antimicrobial Consumption in the EU/EEA (ESAC-Net)—Annual Epidemiological Report 2021. 2022. Available online: https://www.ecdc.europa.eu/ (accessed on 12 January 2024).
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D.D. Emerging Antimicrobial-Resistant High-Risk Klebsiella pneumoniae Clones ST307 and ST147. Antimicrob. Agents Chemother. 2020, 64, e01148-20. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Desai, S.; Passet, V.; Gajjar, D.; Brisse, S. Genomic evolution of the globally disseminated multidrug-resistant Klebsiella pneumoniae clonal group 147. Microb. Genom. 2022, 8, 000737. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Leon, I.; Perez-Nadales, E.; Marin-Sanz, J.A.; Garcia-Martinez, T.; Martinez-Martinez, L. Heteroresistance to colistin in wild-type Klebsiella pneumoniae isolates from clinical origin. Microbiol. Spectr. 2023, 11, e0223823. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Xu, L.; Wang, Y.; Fu, H.; Xiao, T.; Yu, W.; Zhou, W.; Zhang, K.; Shen, J.; Ji, J.; et al. Clinical relevance, mechanisms and evolution of polymyxin B heteroresistance carbapenem-resistant Klebsiella pneumoniae: A genomic, retrospective cohort study. Clin. Microbiol. Infect. 2024. [Google Scholar] [CrossRef]
- El-Halfawy, O.M.; Valvano, M.A. Antimicrobial heteroresistance: An emerging field in need of clarity. Clin. Microbiol. Rev. 2015, 28, 191–207. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Results | No. of Isolates | |
|---|---|---|
| Fulfilling C1 | Growth on the plates containing 4 and/or 8 mg/L of colistin with a frequency of at least 1 × 10−6 | 25 |
| Fulfilling only C2 | Growth on at least all the plates containing colistin at a concentration up to and including eight-fold the MIC of the isolate at a frequency of minimally 1 × 10−7, minimum concentration at which there should be growth was 4 mg/L | 83 |
| Not fulfilling either classification but growth > 2 mg/L | Growth on 4 and/or 8 mg/L plate but frequency < 1 × 10−7 | 45 |
| For MIC 0.0625–0.5 mg/L: growth with frequency ≥ 1 × 10−7 on 8 mg/L plate but <1 × 10−7 on 4 mg/L plate | 8 | |
| For MIC 1–2 mg/L: growth with frequency ≥ 1 × 10−7 on 4 and/or 8 mg/L plate but <1 × 10−7 on plates ≥ eight-fold the MIC | 30 | |
| Growth with frequency ≥ 1 × 10−7 on plates ≥ eight-fold the MIC but frequency of 1 × 10−7 not reached on all plates < eight-fold the MIC | 19 | |
| No growth at 4 and 8 mg/L | 71 | |
| No growth at 1, 2, 4 and 8 mg/L | 7 | |
| Isolate ID | MIC (mg/L) | ST | PAP Assay Plate Conc. (mg/L) | Mutations in mgrB | Other Mutations |
|---|---|---|---|---|---|
| IT0307CP (CR) | 128 | ST409 | IS1R of IS1 family interruption at nt 107 | ||
| IT0636C (CR) | 128 | ST409 | ISKpn34 of IS3 family interruption at nt 46 | ||
| IT0915C (CR) | 64 | ST409 | IS903B of IS5 family interruption at nt 34 | ||
| IT0244CP (CHR 1st PAP) | 0.5 | ST409 | 2 | ISKpn34 of IS3 family interruption at promoter | |
| 8 | IS903B of IS5 family interruption at nt 117 | ||||
| 16 | IS1S of IS1 family interruption at promoter | ||||
| IT0244CP (CHR 3rd PAP) | 0.5 | ST409 | 2 | IS1X2 of IS1 family interruption at nt 123 | |
| AN1505CP2 (CHR 1st PAP) | 1 | ST323 | 4 | Deleted | |
| 8 | IS903B of IS5 family interruption at nt 70 | ||||
| AN1505CP2 (CHR 2nd PAP) | 1 | ST323 | 8 | pmrB: T157P | |
| AN1505CP2 (CHR 3rd PAP) | 1 | ST323 | 8 | Q30X | |
| 16 | Deleted |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braspenning, A.J.M.M.; Rajakani, S.G.; Sey, A.; El Bounja, M.; Lammens, C.; Glupczynski, Y.; Malhotra-Kumar, S. Assessment of Colistin Heteroresistance among Multidrug-Resistant Klebsiella pneumoniae Isolated from Intensive Care Patients in Europe. Antibiotics 2024, 13, 281. https://doi.org/10.3390/antibiotics13030281
Braspenning AJMM, Rajakani SG, Sey A, El Bounja M, Lammens C, Glupczynski Y, Malhotra-Kumar S. Assessment of Colistin Heteroresistance among Multidrug-Resistant Klebsiella pneumoniae Isolated from Intensive Care Patients in Europe. Antibiotics. 2024; 13(3):281. https://doi.org/10.3390/antibiotics13030281
Chicago/Turabian StyleBraspenning, Anouk J. M. M., Sahaya Glingston Rajakani, Adwoa Sey, Mariem El Bounja, Christine Lammens, Youri Glupczynski, and Surbhi Malhotra-Kumar. 2024. "Assessment of Colistin Heteroresistance among Multidrug-Resistant Klebsiella pneumoniae Isolated from Intensive Care Patients in Europe" Antibiotics 13, no. 3: 281. https://doi.org/10.3390/antibiotics13030281
APA StyleBraspenning, A. J. M. M., Rajakani, S. G., Sey, A., El Bounja, M., Lammens, C., Glupczynski, Y., & Malhotra-Kumar, S. (2024). Assessment of Colistin Heteroresistance among Multidrug-Resistant Klebsiella pneumoniae Isolated from Intensive Care Patients in Europe. Antibiotics, 13(3), 281. https://doi.org/10.3390/antibiotics13030281