

Liposomes-Based Drug Delivery Systems of Anti-Biofilm Agents to Combat Bacterial Biofilm Formation


Abstract

1. Introduction
2. Bacterial Biofilms
3. Challenges in Targeting Biofilms
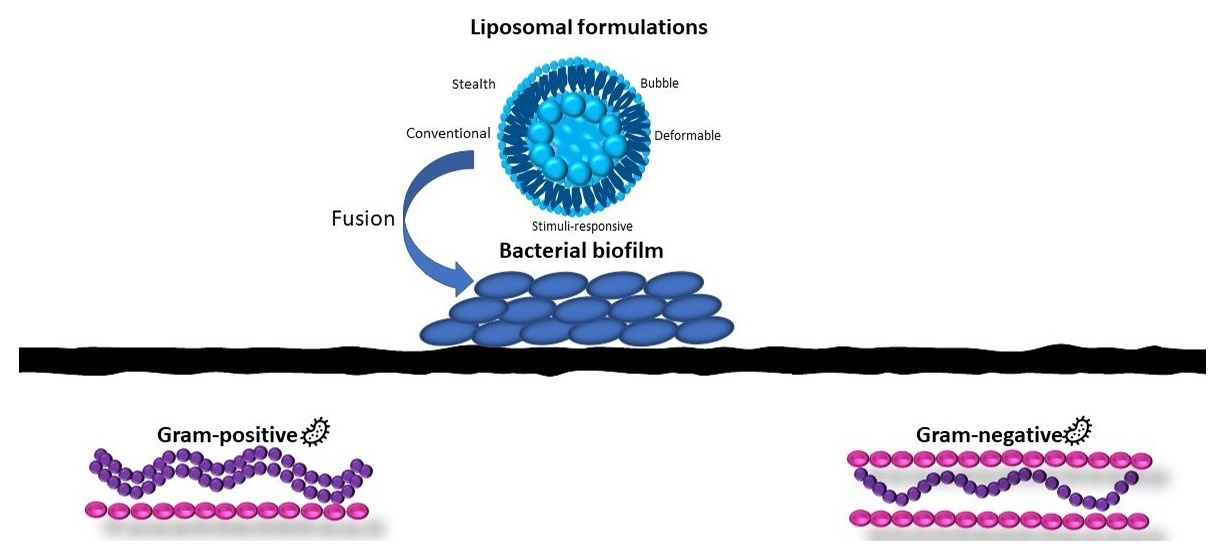
4. Liposomes: Versatile Drug Delivery Systems
5. Liposomal Formulations for the Treatment of Gram-Negative Biofilms
5.1. Pseudomonas aeruginosa Biofilms
5.2. Escherichia coli Biofilms
5.3. Acinetobacter baumannii Biofilms
5.4. Salmonella Species Biofilms

{kind=link}
{kind=link}
| Bacterial Strain | Liposome Type | Active Compound | Dosage | Efficiency | Refs. |
|---|---|---|---|---|---|
| Pseudomonas aeruginosa | Cationic and anionic with/out PEGylation | None | 0–8 μmol/mL. | Anionic liposomes (-) Cationic liposomes (+) PEGylated cationic liposomes (++) | [81] |
| Surface cationic lysozyme liposomes | Gentamicin | 10 μL | Liposomal gentamicin with surface cationic lysozyme (++) Control (-) | [84] | |
| Cationic liposomes | Tobramycin + anti-biofilm peptide | ≥4 and ≤32 μg/mL | Tobramycin enclosed in liposomes, with/out peptide (++) | [36] | |
| Neutral: NLG Negatively charged liposomes: NLG-1 and NLG-2. | Gentamicin | NLG: 0.125–0.5 mg/L. | Pseudomonas aeruginosa: NLG and NLG-1 (++) Control and NLG-2 (+) K. oxytoca: Control (-) NLG (++) NELG-1, and NELG-2 (+) | [78] | |
| Cationic Liposomes + photodynamic therapy | DVDMS | 10 μg/mL of DVDMS | Cationic lipid-mediated nano-DVDMS with Photodynamic therapy (++) Control (-) | [87] | |
| Photodynamic liposome | Perfluorohexane and photosensitizer | 100 μL | Photodynamic liposomal mixture (++) | [69] | |
| Bimodal photodynamic and photothermal liposomes | Phospholipid-porphyrin conjugates + pheophorbide-a (PhxLPC)/pyropheophorbide-a (PyrxLPC) | Ph3LPC:1.63 μM Pyr3LPC: 5.45 μM | Ph3LPC (++) Pyr3LPC (+) | [67] | |
| Thermosensitive liposomes of varying DSPC: cypate: Betainylated Cholesterol ratios TSL1: (4:0.5:1) TSL2: (4:0.5:2) TSL3: (4:0.5:3) | Tobramycin + cyanine dye | 15.6 ug/mL | TSL1 (+) TSL2 (+) TSL3 (++) | [85] | |
| Light responsive liposomes | Tobramycin + gold nanoparticles (Lip-AuNP) + graphene quantum dots (GQD) | 16 µg mL−1 | Tobramycin with Lip-AuNP or GQD (-) | [89] | |
| Temperature-sensitive liposomes + alternating magnetic fields | Ciprofloxacin | MIC = 0.5 µg/mL | Free/liposomal ciprofloxacin (+) | [88] | |
| Nanoarchaeosome | Azithromycin (AZ) | 8 μg/mL | AZ-nanovesicles/nanoarchaeosome (++) Free AZ (-) | [90] | |
| Neutral liposomes | Meropenem | 6.25 µg/mL | Liposomal meropenem (++) Control (+) | [40] | |
| Adjuvant liposomes | Azithromycin (AZI) + (2-nitroimidazole derivative, 6-NIH) + (nitric oxide donor, DETA NONOate). | 120 μg/mL | Liposomal mixture (++) Control (+) | [91] | |
| Neutral liposomes | Cefoperazone sodium | IC50: 0.42 μg/mL | Liposomal cefoperazone (++) Control (+) | [83] | |
| Neutral liposomes | Gentamicin (GEN) or erythromycin (ERY) + PABN | Liposomal GEN with PABN: 1–8 mg/mL Liposomal ERY with PABN: 64–128 mg/mL | Against PA01: PABNA with liposomal GEN/ERY (++) Against PA11: PABNA with liposomal GEN (++) PABNA with liposomal ERY (+) | [93] | |
| Proliposomes | Tobramycin + clarithromycin (TOB/CLA-CPROLips) | 0.06 to 16 mg/L | TOB/CLA (-) CPROLips (++) Control (+) | [108] | |
| Neutral liposomes | Perfluorohexane followed by aztreonam | 100 μL | Sequential treatment (++) Control (-) | [16] | |
| Neutral liposomes | Chitosan + phosphatidylcholine + gentamicin | 10 μL of liposomes loaded with 20 μg/mL gentamicin | Liposomal mixture (++) Control (+) | [95] | |
| Neutral liposomes in Chitosan hydrogels. | Chlorhexidine | 100 μL | Chitosan hydrogels loaded with liposomal chlorhexidine (++) Control (+) | [94] | |
| Neutral liposomes | Copper | - | Liposomes loaded with copper nanoparticles (-) | [96] | |
| Neutral liposomes | Lipopeptide surfactant + copper nanoparticles | 89 μg/mL | Liposomes enclosing lipopeptide surfactant with copper nanoparticles (+) Control (+) | [44] | |
| Phosphatidylcholine/cholesterol liposomes with stearylamine: (LP-SA/CDC) and (LP-SA/PF) | Novel quorum sensing inhibitors: “CDC” and “PF” | 1–10 µM | LP-SA/CDC (+) LP-SA/PF (++) | [97] | |
| Liposomes made of cholesteryl linoleate (CL-PL) | human beta-defensin 2 (HBD2) | CL:125 µg/mL HBD2: 0.25 µM. | CL-PL with HBD2 (++) Control (+) | [98] | |
| Neutral liposomes | Commercial ozonated sunflower oil + hypromellose | Final concentration of 20% of the ozonated sunflower oil | Liposomal formulations (++) | [109] | |
| Escherichia coli Pseudomonas aeruginosa | Rhamnosome nano-vesicles | Nisin Z | Escherichia coli: 25 μg/mL Pseudomonas aeruginosa 100 μg/mL | Nisin Z encapsulating rhamnosomes (++) Control (+) | [100] |
| Escherichia coli | Thermal Liposomes | Tungsten sulfide quantum dots (WS2QDs) and vancomycin | 50 μg/mL | Liposomal formulations with NIR (++) Control (-) | [68] |
| Cationic liposomes made of DPPC:Chol:DOTAC. Lip1 (1:0.5:0.3) Lip 2 (1:0.5:0.5) Lip 3 (1:0.5:0.8) Lip 4 (1:0.5:1.2) | Methylene blue | Final molar concentration of MB of 0.02% w/v | Lip3, Lip2, Lip1 (+) Lip4, Control (-) | [101] | |
| Neutral liposomes | Azithromycin (LA) or azithromycin/N-acetylcysteine (LAN) | 2.5–3 μg/mL | Strain SA057: LA and LAN (++) Free drugs+ Strain SA10: LA (+) LAN and Control (++) | [99] | |
| Negative liposomes with rigid bilayers (CL-3), propylene glycol liposomes (PGL-2), or deformable propylene glycol liposomes (DPGL-2) | Azithromycin (AZI) | Varied among strains and liposomal formulations within 1.26–45.65 µg/mL | DPGL-2 (++) CL-3 and PGL-2 (+) | [102] | |
| Solid liposomes | Clove oil | 0.5 mg/mL | Liposomal formulations (+) | [110] | |
| Functionalized PEG-liposomes. | MTAB | In 3.6 µmol of lipids, 67.5 µg/mL of attached MTAB | Liposomal MTAB (++) Free MTAB (+) | [111] | |
| Acinetobacter baumannii | Chitosan-modified liposomes (CLPs) + ultrasound microbubbles (USMBs) | Polymyxin B | 8 ± 2 µg/mL | USMBs with CLPs (++) Control (+) | [71] |
| Silver sulfadiazine liposomes (AgSD-NLs@Cur) | Curcumin | 7.8 μg/mL | LED plus AgSD-NLs@Cur (++) Control (-) | [105] | |
| Neutral liposomes | Thymoquinone (TQ) | TQ: 2 and Lip-TQ: 4 µg/mL | Control (++) Lip-TQ (+) | [104] | |
| Acinetobacter baumanii Escherichia coli Klebsiella pneumoniae | Neutral liposomes | Tobramycin or tobramycin-N-acetylcysteine | Varying across strain and formulation within 8 to 128 mg/L. | Control (+) TNL and TL (++) | [103] |
| Salmonella enterica | Neutral liposomes | Thymol and carvacrol | 0.662 mg/mL | Control (++) TCL (+) | [107] |
| Salmonella typhimurium | Neutral liposomes | Geraniol | 0.10% | Geraniol-Loaded Liposomal (+) | [106] |
| Aeromonas hydrophila Serratia grimesii | Neutral liposomes | Curcumin | 0.35 mg/mL | Curcumin liposomes (++) | [112] |
| Aeromonas sobria | Neutral liposomes | Curcumin | 420 µg/mL | Control and liposomal curcumin (++) | [113] |
| Porphyromonas gingivalis | Liposomes consisting of quaternary ammonium chitosan (TMC) | Doxycycline (DOX) | 1 mL | TMC-lip-DOX (++) Control (+) | [114] |
| Porphyromonas gingivalis and Prevotella intermedia mixed biofilm | Liposomes (TMC-Lip-DOX NPs) composed of TMC | Doxycycline | 1 mL | TMC-Lip-DOX NPs (++) Control (+) | [115] |
6. Liposomal Formulations for the Treatment of Gram-Positive Biofilms
6.1. Staphylococcus aureus Biofilms
6.2. Staphylococcus epidermis
6.3. Staphylococcus saprophyticus subsp. Bovis Biofilms
6.4. Streptococci Species
6.5. Other Gram-Positive Species Biofilms
| Bacterial Strain | Liposome Type | Active Compound | Dosage | Efficiency | Refs. |
|---|---|---|---|---|---|
| MRSA | Glycosylated cationic liposomes | Trans-resveratrol | 1.2 mM MIC | Galactosylated liposomes (++) Mannosylated liposomes (+) Glucosylated liposomes (-) | [35] |
| Mannose-modified liposomes | Platensimycin | 0.5 to 8 μg/mL | Mannose-modified liposomal platensimycin and control (+) | [126] | |
| Liposome microbubbles | HBD-3 | - | Liposomal formulations (+) | [72] | |
| Neutral Liposomes | SME | 0.1 mg/mL SME | Liposomal formulations (+) | [32] | |
| Multilamellar liposomes | Lipopeptide surfactant + copper | 105 μg/mL | Liposomal formulations (+) | [44] | |
| Neutral liposomes | Biosurfactant | 1.25–5 mg/mL biosurfactant | Liposomal formulations (+) | [131] | |
| Conventional neutral, deformable, propylene glycol, and cationic liposomes | Azithromycin | 0.5–8 µg/mL MBIC | Cationic liposomes (++) neutral, deformable, and propylene glycol liposomes (+) | [34] | |
| Neutral liposomes | Berberine + curcumin | 8 µg/mL berberine + 10 µg/mL curcumin MIC | Liposomal formulations (+) | [128] | |
| Stimuli-responsive multilamellar liposomes | Vancomycin | 0.97 μg/mL | Liposomal formulations (+) | [132] | |
| MSSA and MRSA | Conventional neutral, fusogenic, and cationic LUVs | Vancomycin | 0.78–1.56 µg/mL MIC | Fusogenic liposomes (++) Cationic and neutral liposomes (+) | [39] |
| MSSA | Stealth liposomes | Nafcillin | 0.5 μg/mL PEGylated formulation MBIC 1 μg/mL unPEGylated formulation MBIC | PEGylated liposomes (++) UnPEGylated liposomes (+) | [133] |
| MSSA ATCC | Cationic, anionic, and neutral liposomes | Rifabutin | 0.006 µg/mL MIC | Liposomal formulations (+) | [134] |
| Vancomycin-intermediate S. aureus | Stimuli-responsive liposome | Tungsten sulfide quantum dots + vancomycin | 50 μg/mL | Liposomal formulations (+) | [68] |
| Staphylococcus aureus | Immunoliposomes | Isosorbide mononitrate and anti-α-hemolysin | 45 mg/mL | Isosorbide mononitrate immunoliposomes (++) Control (+) | [59] |
| Neutral liposomes | Commercial ozonated sunflower oil “Ozodrop” and “Ozodrop gel” | 20% Ozodrop 20% Ozodrop gel | Liposomal formulations (+) | [109] | |
| Chitosan/gum arabic-coated liposomes | Alizarin | 2, 10, 20, and 50 μg/mL | Liposomal formulations (+) | [143] | |
| Neutral Liposomes | Copper | Unmentioned | Liposomal formulations (-) | [96] | |
| Neutral liposomes | Toluidine Blue O | 5, 10, and 25 ppm | Liposomal formulations (+) | [142] | |
| Cationic liposomes | Cefquinome sulfate | 0.48 μg/mL MIC, and 0.5, 1.5, 2, and 2.5 of the MIC | Liposomal formulations (+) | [136] | |
| Neutral liposomes | Levofloxacin and serratiopeptidase | gg/mL serratiopeptidase | Liposomal formulations (+) | [137] | |
| Gold nanoparticle-coated liposomes | Curcumin | 200 µg/mL | Liposomal formulations (+) | [140] | |
| Anionic liposomes | Lysozyme and gentamicin (resulting in cationic formulation) | Unmentioned | Liposomal formulations (+) | [84] | |
| Neutral liposomes | Lysozyme and levofloxacin | Unmentioned | Liposomal formulations (-) | [139] | |
| Staphylococcus aureus | Neutral liposomes | Azithromycin | 4 μg/mL MIC | Liposomal formulations:
| [90] |
| Staphylococcus aureus | Phospholipid-porphyrin liposome-like assemblies | Photosensitizer | 100 μM photosensitizer | Liposomal formulations (+) | [67] |
| Staphylococcus aureus | Neutral liposomes coated with chitosan hydrogel | Chlorhexidine | Unmentioned | Liposomal formulations (+) | [94] |
| Staphylococcus aureus | Stimuli-responsive liposomes | LysRODI | 40 µg/mL LysRODI | Liposomal formulations (+) | [141] |
| Staphylococcus aureus | Stimuli-responsive liposomes | Vancomycin, rifampicin, and SA31 aptamer | 25 μg/mL MBEC | Liposomal formulations (+) | [60] |
| Streptococcus epidermidis | Liposomes incorporated into hydrogels | Tetracycline HCl and tretinoin | 0.25 µg/mL MBIC | Liposomal formulations (+) | [145] |
| Cationic liposomes | Rifampin and N-acetylcysteine | Unmentioned | Liposomal rifampin/N-acetylcysteine or N-acetylcysteine:
| [144] | |
| Neutral liposomes | Moxifloxacin | 43 µM lipid + 0.3 µM moxifloxacin | Liposomal formulations (+) | [146] | |
| Neutral liposomes | Vancomycin and C2DA | 0.83 mg/mL vancomycin + 0.64 mg/mL C2DA | Liposomal formulations (+) | [147] | |
| Staphylococcus epidermidis | Glycosylated cationic liposomes | Usnic acid | 1 mM total lipids + 0.05 mM (MIC) or 0.01 mM (1/5 MIC) usnic acid | Liposomal formulations (+) | [148] |
| Staphylococcus saprophyticus subspecies bovis | Neutral liposomes | Hypericin and (Hyp-HPβCD) | Unmentioned | Liposomal formulations (+) | [149] |
| Neutral liposomes | Hypericin and HPβCD | 40 µM | Liposomal formulations (+) | [150] | |
| S. pneumonaie | Deformable stealth or sodium cholate liposomes | Endolysin MSlys | 4 μM MSlys | Stealth liposomes (++) Sodium cholate liposomes (+) | [153] |
| S. oralis and S. mutans | Anionic liposomes | α-TP | (+) α-TP 0.8 mM | In positive buffer (+) In negative buffer (-) | [45] |
| C. acnes | Cationic liposomes | SME, DNase I, and proteinase K | 150 μg/mL SME + 25 μg/mL DNase I + 10 μg/mL proteinase K | Liposomal formulations (+) | [157] |
| Bacillus subtilis | Neutral liposomes | Energy-coupling factor transporter inhibitor (HIPS5031) | 9.64 µM | Liposomal formulations (+) | [158] |
| Mycobacterium avium | Neutral liposomes | Amikacin | >16 μg/mL | Liposomal formulations (+) | [160] |
| Mycobacterium avium subsp. hominissuis and Mycobacterium abscessus | Neutral liposomes | Ciprofloxacin | g/mL | Liposomal formulations (+) | [159] |
| Listeria monocytogenes and Staphylococcus aureus | Neutral liposomes | Rhamnolipid and nisin | Listeria monocytogenes: 2.5 μg/mL MIC Staphylococcus aureus: 1.25 μg/mL MIC | Liposomal formulations (+) | [100] |
| Listeria monocytogenes | Cationic liposomes | Alpep7 protein | 52.78 μg/mL (4 μg/mL of Alpep7) | Liposomal formulations (+) | [161] |
| Listeria monocytogenes | Liposomal chitosan | Gentamicin | 20 μg/mL gentamicin | Liposomal formulations (+) | [95] |
7. Literature Gaps in the Treatment of Gram-Negative and Gram-Positive Biofilms Using Liposomal Systems
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 6-AIH | 2-aminomidazole derivative |
| 6-NIH | 6-(2-nitroimidazole) hexylamine |
| α-TP | alpha-tocopherol phosphate |
| BC | Betainylated Cholesterol |
| C2DA | cis-2-decenoic acid |
| DODAB | dimethyldioctadecylammonium bromide |
| DOPE | 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine |
| DOTAC | dimethyldioctadecyl-ammonium chloride |
| DOTAP | 2,3-dioleoyloxy-propyl)-trimethylammonium-chloride |
| DPPC | dipalmitoylphosphatidyl-choline cholesterol |
| DSPC | distearoylphosphtidylcholine |
| DVDMS | nano-Sinoporphyrin sodium |
| DVDMS | Sinoporphyrin sodium |
| HBD-3 | Human beta-Defensin-3 |
| Hyp-HPβCD | (2-hydroxypropyl)-β-cyclodextrine |
| LysRODI | antistaphylococcal endolysin |
| MTAB | (11-mercaptoundecyl)-N,N,N-trimethylammonium bromide |
| PAβN | phenylalanine arginine beta-naphthylamide |
| PGP-me | 2,3-di-O-phytanyl-sn-glycero-1-phospho-(3′-sn-glycerol-1′-methylphosphate) |
| SME | soyaethyl morpholinium ethosulfate |
| TMC | N,N,N-trimethyl chitosan |
| TRIS | tris-(hydroxymethyl)aminomethane |
References
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial Biofilms and Their Implications in Pathogenesis and Food Safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial Biofilm Inhibition: A Focused Review on Recent Therapeutic Strategies for Combating the Biofilm Mediated Infections. Front. Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, R.; Omar, N.; Dawaba, M.; Moemen, D. Bacterial Biofilm Dependent Catheter Associated Urinary Tract Infections: Characterization, Antibiotic Resistance Pattern and Risk Factors. Egypt. J. Basic Appl. Sci. 2021, 8, 64–74. [Google Scholar] [CrossRef]
- Bi, Y.; Xia, G.; Shi, C.; Wan, J.; Liu, L.; Chen, Y.; Wu, Y.; Zhang, W.; Zhou, M.; He, H.; et al. Therapeutic Strategies against Bacterial Biofilms. Fundam. Res. 2021, 1, 193–212. [Google Scholar] [CrossRef]
- An, A.Y.; Choi, K.-Y.G.; Baghela, A.S.; Hancock, R.E.W. An Overview of Biological and Computational Methods for Designing Mechanism-Informed Anti-Biofilm Agents. Front. Microbiol. 2021, 12, 640787. [Google Scholar] [CrossRef]
- Singh, A.; Amod, A.; Pandey, P.; Bose, P.; Pingali, M.S.; Shivalkar, S.; Varadwaj, P.K.; Sahoo, A.K.; Samanta, S.K. Bacterial Biofilm Infections, Their Resistance to Antibiotics Therapy and Current Treatment Strategies. Biomed. Mater. 2022, 17, 022003. [Google Scholar] [CrossRef]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural Products as Inspiration for the Development of Bacterial Antibiofilm Agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- Wang, D.-Y.; van der Mei, H.C.; Ren, Y.; Busscher, H.J.; Shi, L. Lipid-Based Antimicrobial Delivery-Systems for the Treatment of Bacterial Infections. Front. Chem. 2020, 7, 872. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Anu Mary Ealia, S.; Saravanakumar, M.P. A Review on the Classification, Characterisation, Synthesis of Nanoparticles and Their Application. IOP Conf. Ser. Mater. Sci. Eng. 2017, 263, 032019. [Google Scholar] [CrossRef]
- Ijaz, I.; Gilani, E.; Nazir, A.; Bukhari, A. Detail Review on Chemical, Physical and Green Synthesis, Classification, Characterizations and Applications of Nanoparticles. Green Chem. Lett. Rev. 2020, 13, 223–245. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Ferreira, M.; Ogren, M.; Dias, J.N.R.; Silva, M.; Gil, S.; Tavares, L.; Aires-da-Silva, F.; Gaspar, M.M.; Aguiar, S.I. Liposomes as Antibiotic Delivery Systems: A Promising Nanotechnological Strategy against Antimicrobial Resistance. Molecules 2021, 26, 2047. [Google Scholar] [CrossRef]
- Rukavina, Z.; Vanić, Ž. Current Trends in Development of Liposomes for Targeting Bacterial Biofilms. Pharmaceutics 2016, 8, 18. [Google Scholar] [CrossRef]
- Wang, Y. Liposome as a Delivery System for the Treatment of Biofilm-mediated Infections. J. Appl. Microbiol. 2021, 131, 2626–2639. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zou, L.; Yu, W.; Jia, F.; Han, H.; Yao, K.; Jin, Q.; Ji, J. Relief of Biofilm Hypoxia Using an Oxygen Nanocarrier: A New Paradigm for Enhanced Antibiotic Therapy. Adv. Sci. 2020, 7, 2000398. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Wu, H.; Høiby, N.; Molin, S.; Song, Z. Current Understanding of Multi-species Biofilms. Int. J. Oral Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef]
- Nadar, S.; Khan, T.; Patching, S.G.; Omri, A. Development of Antibiofilm Therapeutics Strategies to Overcome Antimicrobial Drug Resistance. Microorganisms 2022, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Abebe, G.M. The Role of Bacterial Biofilm in Antibiotic Resistance and Food Contamination. Int. J. Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and Its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of Intervention, Treatment, and Antibiotic Resistance of Biofilm-Forming Microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial Biofilm and Associated Infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Mahamuni-Badiger, P.P.; Patil, P.M.; Badiger, M.V.; Patel, P.R.; Thorat-Gadgil, B.S.; Pandit, A.; Bohara, R.A. Biofilm Formation to Inhibition: Role of Zinc Oxide-Based Nanoparticles. Mater. Sci. Eng. C 2020, 108, 110319. [Google Scholar] [CrossRef] [PubMed]
- Hawas, S.; Verderosa, A.D.; Totsika, M. Combination Therapies for Biofilm Inhibition and Eradication: A Comparative Review of Laboratory and Preclinical Studies. Front. Cell. Infect. Microbiol. 2022, 12, 850030. [Google Scholar] [CrossRef]
- Ghosh, A.; Jayaraman, N.; Chatterji, D. Small-Molecule Inhibition of Bacterial Biofilm. ACS Omega 2020, 5, 3108–3115. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial Biofilm Eradication Agents: A Current Review. Front. Chem. 2019, 7, 824. [Google Scholar] [CrossRef]
- Haney, E.F.; Trimble, M.J.; Hancock, R.E.W. Microtiter Plate Assays to Assess Antibiofilm Activity against Bacteria. Nat. Protoc. 2021, 16, 2615–2632. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, Y.; Lin, S.; Zhang, W.; Shu, G.; Lin, J.; Li, H.; Xu, F.; Tang, H.; Peng, G.; et al. Strategies for Interfering with Bacterial Early Stage Biofilms. Front. Microbiol. 2021, 12, 675843. [Google Scholar] [CrossRef]
- Khan, F.; Pham, D.T.N.; Oloketuyi, S.F.; Manivasagan, P.; Oh, J.; Kim, Y.-M. Chitosan and Their Derivatives: Antibiofilm Drugs against Pathogenic Bacteria. Colloids Surf. B Biointerfaces 2020, 185, 110627. [Google Scholar] [CrossRef]
- Lin, Y.-K.; Yang, S.-C.; Hsu, C.-Y.; Sung, J.-T.; Fang, J.-Y. The Antibiofilm Nanosystems for Improved Infection Inhibition of Microbes in Skin. Molecules 2021, 26, 6392. [Google Scholar] [CrossRef]
- Dos Santos Ramos, M.A.; Da Silva, P.; Spósito, L.; De Toledo, L.; Bonifácio, B.; Rodero, C.F.; Dos Santos, K.; Chorilli, M.; Bauab, T.M. Nanotechnology-Based Drug Delivery Systems for Control of Microbial Biofilms: A Review. Int. J. Nanomed. 2018, 13, 1179–1213. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-H.; Hung, C.-F.; Aljuffali, I.A.; Sung, C.T.; Huang, C.-T.; Fang, J.-Y. Cationic Amphiphile in Phospholipid Bilayer or Oil–Water Interface of Nanocarriers Affects Planktonic and Biofilm Bacteria Killing. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Nsairat, H.; Khater, D.; Sayed, U.; Odeh, F.; Al Bawab, A.; Alshaer, W. Liposomes: Structure, Composition, Types, and Clinical Applications. Heliyon 2022, 8, e09394. [Google Scholar] [CrossRef]
- Rukavina, Z.; Šegvić Klarić, M.; Filipović-Grčić, J.; Lovrić, J.; Vanić, Ž. Azithromycin-Loaded Liposomes for Enhanced Topical Treatment of Methicillin-Resistant Staphyloccocus Aureus (MRSA) Infections. Int. J. Pharm. 2018, 553, 109–119. [Google Scholar] [CrossRef]
- Aiello, S.; Pagano, L.; Ceccacci, F.; Simonis, B.; Sennato, S.; Bugli, F.; Martini, C.; Torelli, R.; Sanguinetti, M.; Ciogli, A.; et al. Mannosyl, Glucosyl or Galactosyl Liposomes to Improve Resveratrol Efficacy against Methicillin Resistant Staphylococcus Aureus Biofilm. Colloids Surf. A Physicochem. Eng. Asp. 2021, 617, 126321. [Google Scholar] [CrossRef]
- Alzahrani, N.M.; Booq, R.Y.; Aldossary, A.M.; Bakr, A.A.; Almughem, F.A.; Alfahad, A.J.; Alsharif, W.K.; Jarallah, S.J.; Alharbi, W.S.; Alsudir, S.A.; et al. Liposome-Encapsulated Tobramycin and IDR-1018 Peptide Mediated Biofilm Disruption and Enhanced Antimicrobial Activity against Pseudomonas Aeruginosa. Pharmaceutics 2022, 14, 960. [Google Scholar] [CrossRef]
- Kluzek, M.; Oppenheimer-Shaanan, Y.; Dadosh, T.; Morandi, M.I.; Avinoam, O.; Raanan, C.; Goldsmith, M.; Goldberg, R.; Klein, J. Designer Liposomic Nanocarriers Are Effective Biofilm Eradicators. ACS Nano 2022, 16, 15792–15804. [Google Scholar] [CrossRef]
- Nicolosi, D.; Scalia, M.; Nicolosi, V.M.; Pignatello, R. Encapsulation in Fusogenic Liposomes Broadens the Spectrum of Action of Vancomycin against Gram-Negative Bacteria. Int. J. Antimicrob. Agents 2010, 35, 553–558. [Google Scholar] [CrossRef]
- Scriboni, A.B.; Couto, V.M.; de Morais Ribeiro, L.N.; Freires, I.A.; Groppo, F.C.; de Paula, E.; Franz-Montan, M.; Cogo-Müller, K. Fusogenic Liposomes Increase the Antimicrobial Activity of Vancomycin Against Staphylococcus Aureus Biofilm. Front. Pharmacol. 2019, 10, 1401. [Google Scholar] [CrossRef]
- Zahra, M.-J.; Hamed, H.; Mohammad, R.-Y.; Nosratollah, Z.; Akbarzadeh, A.; Morteza, M. Evaluation and Study of Antimicrobial Activity of Nanoliposomal Meropenem against Pseudomonas Aeruginosa Isolates. Artif. Cells Nanomed. Biotechnol. 2017, 45, 975–980. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, A.; Silletti, M.F.; Faraone, I.; Milella, L. Nanoparticulate Antibiotic Systems as Antibacterial Agents and Antibiotic Delivery Platforms to Fight Infections. J. Nanomater. 2020, 2020, 6905631. [Google Scholar] [CrossRef]
- Antoniou, A.I.; Giofrè, S.; Seneci, P.; Passarella, D.; Pellegrino, S. Stimulus-Responsive Liposomes for Biomedical Applications. Drug Discov. Today 2021, 26, 1794–1824. [Google Scholar] [CrossRef]
- Kannan, S.; Solomon, A.; Krishnamoorthy, G.; Marudhamuthu, M. Liposome Encapsulated Surfactant Abetted Copper Nanoparticles Alleviates Biofilm Mediated Virulence in Pathogenic Pseudomonas Aeruginosa and MRSA. Sci. Rep. 2021, 11, 1102. [Google Scholar] [CrossRef]
- Harper, R.A.; Carpenter, G.H.; Proctor, G.B.; Harvey, R.D.; Gambogi, R.J.; Geonnotti, A.R.; Hider, R.; Jones, S.A. Diminishing Biofilm Resistance to Antimicrobial Nanomaterials through Electrolyte Screening of Electrostatic Interactions. Colloids Surf. B Biointerfaces 2019, 173, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.M.A.; Abdallah, O.Y.; Naggar, V.F.; Khalafallah, N.M. Deformable Liposomes and Ethosomes: Mechanism of Enhanced Skin Delivery. Int. J. Pharm. 2006, 322, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Nayak, D.; Tippavajhala, V.K. A Comprehensive Review on Preparation, Evaluation and Applications of Deformable Liposomes. Iran. J. Pharm. Res. 2021, 20, 186. [Google Scholar] [CrossRef]
- Knop, K.; Hoogenboom, R.; Fischer, D.; Schubert, U.S. Poly(Ethylene Glycol) in Drug Delivery: Pros and Cons as Well as Potential Alternatives. Angew. Chem. Int. Ed. 2010, 49, 6288–6308. [Google Scholar] [CrossRef]
- Gabizon, A.; Catane, R.; Uziely, B.; Kaufman, B.; Safra, T.; Cohen, R.; Martin, F.; Huang, A.; Barenholz, Y. Prolonged Circulation Time and Enhanced Accumulation in Malignant Exudates of Doxorubicin Encapsulated in Polyethylene-Glycol Coated Liposomes. Cancer Res. 1994, 54, 987–992. [Google Scholar]
- Ghaferi, M.; Raza, A.; Koohi, M.; Zahra, W.; Akbarzadeh, A.; Ebrahimi Shahmabadi, H.; Alavi, S.E. Impact of PEGylated Liposomal Doxorubicin and Carboplatin Combination on Glioblastoma. Pharmaceutics 2022, 14, 2183. [Google Scholar] [CrossRef]
- Maruyama, K.; Yuda, T.; Okamoto, A.; Kojima, S.; Suginaka, A.; Iwatsuru, M. Prolonged Circulation Time in Vivo of Large Unilamellar Liposomes Composed of Distearoyl Phosphatidylcholine and Cholesterol Containing Amphipathic Poly(Ethylene Glycol). Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1992, 1128, 44–49. [Google Scholar] [CrossRef]
- McNeeley, K.M.; Annapragada, A.; Bellamkonda, R.V. Decreased Circulation Time Offsets Increased Efficacy of PEGylated Nanocarriers Targeting Folate Receptors of Glioma. Nanotechnology 2007, 18, 385101. [Google Scholar] [CrossRef]
- Zhang, L.; Han, L.; Sun, X.; Gao, D.; Qin, J.; Wang, J. The Use of PEGylated Liposomes to Prolong the Circulation Lifetime of Salvianolic Acid B. Fitoterapia 2012, 83, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Bendas, G. Immunoliposomes: A Promising Approach to Targeting Cancer Therapy. BioDrugs 2001, 15, 215–224. [Google Scholar] [CrossRef]
- Gholizadeh, S.; Dolman, E.M.; Wieriks, R.; Sparidans, R.W.; Hennink, W.E.; Kok, R.J. Anti-GD2 Immunoliposomes for Targeted Delivery of the Survivin Inhibitor Sepantronium Bromide (YM155) to Neuroblastoma Tumor Cells. Pharm. Res. 2018, 35, 85. [Google Scholar] [CrossRef]
- Huwyler, J.; Wu, D.; Pardridge, W.M. Brain Drug Delivery of Small Molecules Using Immunoliposomes. Proc. Natl. Acad. Sci. USA 1996, 93, 14164–14169. [Google Scholar] [CrossRef]
- Loureiro, J.A.; Gomes, B.; Fricker, G.; Cardoso, I.; Ribeiro, C.A.; Gaiteiro, C.; Coelho, M.A.N.; do Carmo Pereira, M.; Rocha, S. Dual Ligand Immunoliposomes for Drug Delivery to the Brain. Colloids Surf. B Biointerfaces 2015, 134, 213–219. [Google Scholar] [CrossRef]
- Robinson, A.M.; Creeth, J.E.; Jones, M.N. The Use of Immunoliposomes for Specific Delivery of Antimicrobial Agents to Oral Bacteria Immobilized on Polystyrene. J. Biomater. Sci. Polym. Ed. 2000, 11, 1381–1393. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Y.; Dong, D.; Li, X.; Li, Z.; Li, S.; Wang, J. Effects of Isosorbide Mononitrate Loaded Nanoparticles Conjugated with Anti-Staphylococcus Aureus A-toxin on Staphylococcus Aureus Biofilms. Exp. Med. 2019, 19, 1267–1274. [Google Scholar] [CrossRef]
- Ommen, P.; Hansen, L.; Hansen, B.K.; Vu-Quang, H.; Kjems, J.; Meyer, R.L. Aptamer-Targeted Drug Delivery for Staphylococcus Aureus Biofilm. Front. Cell. Infect. Microbiol. 2022, 12, 814340. [Google Scholar] [CrossRef]
- Wei, Z.; Zhou, Y.; Wang, R.; Wang, J.; Chen, Z. Aptamers as Smart Ligands for Targeted Drug Delivery in Cancer Therapy. Pharmaceutics 2022, 14, 2561. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Gitter, B.; Rüger, R.; Wieland, G.D.; Chen, M.; Liu, X.; Albrecht, V.; Fahr, A. Antimicrobial Peptide-Modified Liposomes for Bacteria Targeted Delivery of Temoporfin in Photodynamic Antimicrobial Chemotherapy. Photochem. Photobiol. Sci. 2011, 10, 1593–1601. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Al-Othman, A.; Al-Sayah, M.H. Multifunctional Stimuli-Responsive Hybrid Nanogels for Cancer Therapy: Current Status and Challenges. J. Control. Release 2022, 351, 476–503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Bai, J.; Yang, W. Stimuli-Responsive Nanocarriers for Therapeutic Applications in Cancer. Cancer Biol. Med. 2021, 18, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Zhao, W.; Song, L.-J.; Luan, S.-F. Stimuli-Responsive Nanocarriers for Bacterial Biofilm Treatment. Rare Met. 2022, 41, 482–498. [Google Scholar] [CrossRef]
- Koga, K.; Tagami, T.; Ozeki, T. Gold Nanoparticle-Coated Thermosensitive Liposomes for the Triggered Release of Doxorubicin, and Photothermal Therapy Using a near-Infrared Laser. Colloids Surf. A Physicochem. Eng. Asp. 2021, 626, 127038. [Google Scholar] [CrossRef]
- Cressey, P.; Bronstein, L.-G.; Benmahmoudi, R.; Rosilio, V.; Regeard, C.; Makky, A. Novel Liposome-like Assemblies Composed of Phospholipid-Porphyrin Conjugates with Photothermal and Photodynamic Activities against Bacterial Biofilms. Int. J. Pharm. 2022, 623, 121915. [Google Scholar] [CrossRef]
- Xu, M.; Hu, Y.; Xiao, Y.; Zhang, Y.; Sun, K.; Wu, T.; Lv, N.; Wang, W.; Ding, W.; Li, F.; et al. Near-Infrared-Controlled Nanoplatform Exploiting Photothermal Promotion of Peroxidase-like and OXD-like Activities for Potent Antibacterial and Anti-Biofilm Therapies. ACS Appl. Mater. Interfaces 2020, 12, 50260–50274. [Google Scholar] [CrossRef]
- Zou, L.; Hu, D.; Wang, F.; Jin, Q.; Ji, J. The Relief of Hypoxic Microenvironment Using an O2 Self-Sufficient Fluorinated Nanoplatform for Enhanced Photodynamic Eradication of Bacterial Biofilms. Nano Res. 2022, 15, 1636–1644. [Google Scholar] [CrossRef]
- Alamoudi, K.; Martins, P.; Croissant, J.G.; Patil, S.; Omar, H.; Khashab, N.M. Thermoresponsive Pegylated Bubble Liposome Nanovectors for Efficient SiRNA Delivery via Endosomal Escape. Nanomedicine 2017, 12, 1421–1433. [Google Scholar] [CrossRef]
- Fu, Y.-Y.; Zhang, L.; Yang, Y.; Liu, C.; He, Y.-N.; Li, P.; Yu, X. Synergistic Antibacterial Effect of Ultrasound Microbubbles Combined with Chitosan-Modified Polymyxin B-Loaded Liposomes on Biofilm-Producing Acinetobacter Baumannii. Int. J. Nanomed. 2019, 14, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Fang, S.; Kong, R.; Zhang, W.; Wu, K.; Xia, R.; Shang, X.; Zhu, C. Effect of Low Frequency Ultrasound plus Fluorescent Composite Carrier in the Diagnosis and Treatment of Methicillin-Resistant Staphylococcus Aureus Biofilm Infection of Bone Joint Implant. Int. J. Clin. Exp. Med. 2018, 11, 799–805. [Google Scholar]
- Raza, A.; Hayat, U.; Rasheed, T.; Bilal, M.; Iqbal, H.M.N. “Smart” Materials-Based near-Infrared Light-Responsive Drug Delivery Systems for Cancer Treatment: A Review. J. Mater. Res. Technol. 2019, 8, 1497–1509. [Google Scholar] [CrossRef]
- Andra, V.V.S.N.L.; Pammi, S.V.N.; Bhatraju, L.V.K.P.; Ruddaraju, L.K. A Comprehensive Review on Novel Liposomal Methodologies, Commercial Formulations, Clinical Trials and Patents. Bionanoscience 2022, 12, 274–291. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the Essentiality of Lipopolysaccharide to Gram-Negative Bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef]
- Bertani, B.; Ruiz, N. Function and Biogenesis of Lipopolysaccharides. EcoSal Plus 2018, 8, ecosalplus.ESP-0001-2018. [Google Scholar] [CrossRef]
- Alhariri, M.; Majrashi, M.A.; Bahkali, A.H.; Almajed, F.S.; Azghani, A.O.; Khiyami, M.; Alyamani, E.J.; Aljohani, S.M.; Halwani, M.A. Efficacy of Neutral and Negatively Charged Liposome-Loaded Gentamicin on Planktonic Bacteria and Biofilm Communities. Int. J. Nanomed. 2017, 12, 6949–6961. [Google Scholar] [CrossRef]
- Mena, K.D.; Gerba, C.P. Risk Assessment of Pseudomonas Aeruginosa in Water. In Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Reviews of Environmental Contamination and Toxicology; Springer: Boston, MA, USA, 2009; Volume 201, pp. 71–115. ISBN 978-1-4419-0031-9. [Google Scholar]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas Aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Ibaraki, H.; Kanazawa, T.; Chien, W.-Y.; Nakaminami, H.; Aoki, M.; Ozawa, K.; Kaneko, H.; Takashima, Y.; Noguchi, N.; Seta, Y. The Effects of Surface Properties of Liposomes on Their Activity against Pseudomonas Aeruginosa PAO-1 Biofilm. J. Drug Deliv. Sci. Technol. 2020, 57, 101754. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas Aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Ghodake, V.; Vishwakarma, J.; Vavilala, S.L.; Patravale, V. Cefoperazone Sodium Liposomal Formulation to Mitigate P. Aeruginosa Biofilm in Cystic Fibrosis Infection: A QbD Approach. Int. J. Pharm. 2020, 587, 119696. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, Z.; Zhang, P.; Bai, H.; Sun, Y.; Duan, J.; Mu, H. Lysozyme Associated Liposomal Gentamicin Inhibits Bacterial Biofilm. Int. J. Mol. Sci. 2017, 18, 784. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dai, X.; Wei, X.; Yu, Y.; Chen, X.; Zhang, X.; Li, C. Near-Infrared Light-Activated Thermosensitive Liposomes as Efficient Agents for Photothermal and Antibiotic Synergistic Therapy of Bacterial Biofilm. ACS Appl. Mater. Interfaces 2018, 10, 14426–14437. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Chi, L.; Hui, P. Review of Stimuli-Responsive Polymers in Drug Delivery and Textile Application. Molecules 2019, 24, 2547. [Google Scholar] [CrossRef]
- Mai, B.; Gao, Y.; Li, M.; Jia, M.; Liu, S.; Wang, X.; Zhang, K.; Liu, Q.; Wang, P. Tailoring the Cationic Lipid Composition of Lipo-DVDMS Augments the Phototherapy Efficiency of Burn Infection. Biomater. Sci. 2021, 9, 2053–2066. [Google Scholar] [CrossRef]
- Munaweera, I.; Shaikh, S.; Maples, D.; Nigatu, A.S.; Sethuraman, S.N.; Ranjan, A.; Greenberg, D.E.; Chopra, R. Temperature-Sensitive Liposomal Ciprofloxacin for the Treatment of Biofilm on Infected Metal Implants Using Alternating Magnetic Fields. Int. J. Hyperth. 2018, 34, 189–200. [Google Scholar] [CrossRef]
- Teirlinck, E.; Barras, A.; Liu, J.; Fraire, J.C.; Lajunen, T.; Xiong, R.; Forier, K.; Li, C.; Urtti, A.; Boukherroub, R.; et al. Exploring Light-Sensitive Nanocarriers for Simultaneous Triggered Antibiotic Release and Disruption of Biofilms Upon Generation of Laser-Induced Vapor Nanobubbles. Pharmaceutics 2019, 11, 201. [Google Scholar] [CrossRef]
- Altube, M.J.; Martínez, M.M.B.; Malheiros, B.; Maffía, P.C.; Barbosa, L.R.S.; Morilla, M.J.; Romero, E.L. Fast Biofilm Penetration and Anti-PAO1 Activity of Nebulized Azithromycin in Nanoarchaeosomes. Mol. Pharm. 2020, 17, 70–83. [Google Scholar] [CrossRef]
- Rao, Y.; Sun, Y.; Li, P.; Xu, M.; Chen, X.; Wang, Y.; Chen, Y.; Deng, X.; Yu, S.; Hu, H. Hypoxia-Sensitive Adjuvant Loaded Liposomes Enhance the Antimicrobial Activity of Azithromycin via Phospholipase-Triggered Releasing for Pseudomonas Aeruginosa Biofilms Eradication. Int. J. Pharm. 2022, 623, 121910. [Google Scholar] [CrossRef]
- Ben, J.; Zhu, X.; Zhang, H.; Chen, Q. Class A1 Scavenger Receptors in Cardiovascular Diseases: SR-A1 and Cardiovascular Diseases. Br. J. Pharm. 2015, 172, 5523–5530. [Google Scholar] [CrossRef] [PubMed]
- Gbian, D.L.; Omri, A. The Impact of an Efflux Pump Inhibitor on the Activity of Free and Liposomal Antibiotics against Pseudomonas Aeruginosa. Pharmaceutics 2021, 13, 577. [Google Scholar] [CrossRef] [PubMed]
- Hemmingsen, L.M.; Giordani, B.; Pettersen, A.K.; Vitali, B.; Basnet, P.; Škalko-Basnet, N. Liposomes-in-Chitosan Hydrogel Boosts Potential of Chlorhexidine in Biofilm Eradication in Vitro. Carbohydr. Polym. 2021, 262, 117939. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Xu, D.; Sui, G.; Wang, D.; Wu, M.; Han, L.; Mu, H.; Duan, J. Gentamicin Decorated Phosphatidylcholine-Chitosan Nanoparticles against Biofilms and Intracellular Bacteria. Int. J. Biol. Macromol. 2020, 156, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Sarcina, L.; García-Manrique, P.; Gutiérrez, G.; Ditaranto, N.; Cioffi, N.; Matos, M.; del Carmen Blanco-López, M. Cu Nanoparticle-Loaded Nanovesicles with Antibiofilm Properties. Part I: Synthesis of New Hybrid Nanostructures. Nanomaterials 2020, 10, 1542. [Google Scholar] [CrossRef]
- Hallan, S.S.; Marchetti, P.; Bortolotti, D.; Sguizzato, M.; Esposito, E.; Mariani, P.; Trapella, C.; Rizzo, R.; Cortesi, R. Design of Nanosystems for the Delivery of Quorum Sensing Inhibitors: A Preliminary Study. Molecules 2020, 25, 5655. [Google Scholar] [CrossRef]
- Beadell, B.A.; Chieng, A.; Parducho, K.R.; Dai, Z.; Ho, S.O.; Fujii, G.; Wang, Y.; Porter, E. Nano- and Macroscale Imaging of Cholesterol Linoleate and Human Beta Defensin 2-Induced Changes in Pseudomonas Aeruginosa Biofilms. Antibiotics 2021, 10, 1279. [Google Scholar] [CrossRef]
- Aljihani, S.A.; Alehaideb, Z.; Alarfaj, R.E.; Alghoribi, M.F.; Akiel, M.A.; Alenazi, T.H.; Al-Fahad, A.J.; Al Tamimi, S.M.; Albakr, T.M.; Alshehri, A.; et al. Enhancing Azithromycin Antibacterial Activity by Encapsulation in Liposomes/Liposomal-N-Acetylcysteine Formulations against Resistant Clinical Strains of Escherichia Coli. Saudi J. Biol. Sci. 2020, 27, 3065–3071. [Google Scholar] [CrossRef]
- Niaz, T.; Shabbir, S.; Noor, T.; Imran, M. Antimicrobial and Antibiofilm Potential of Bacteriocin Loaded Nano-Vesicles Functionalized with Rhamnolipids against Foodborne Pathogens. LWT 2019, 116, 108583. [Google Scholar] [CrossRef]
- Boccalini, G.; Conti, L.; Montis, C.; Bani, D.; Bencini, A.; Berti, D.; Giorgi, C.; Mengoni, A.; Valtancoli, B. Methylene Blue-Containing Liposomes as New Photodynamic Anti-Bacterial Agents. J. Mater. Chem. B 2017, 5, 2788–2797. [Google Scholar] [CrossRef]
- Vanić, Ž.; Rukavina, Z.; Manner, S.; Fallarero, A.; Uzelac, L.; Kralj, M.; Amidžić Klarić, D.; Bogdanov, A.; Raffai, T.; Virok, D.P.; et al. Azithromycin-Liposomes as a Novel Approach for Localized Therapy of Cervicovaginal Bacterial Infections. Int. J. Nanomed. 2019, 14, 5957–5976. [Google Scholar] [CrossRef] [PubMed]
- Alarfaj, R.E.; Alkhulaifi, M.M.; Al-Fahad, A.J.; Aljihani, S.; Yassin, A.E.B.; Alghoribi, M.F.; Halwani, M.A. Antibacterial Efficacy of Liposomal Formulations Containing Tobramycin and N-Acetylcysteine against Tobramycin-Resistant Escherichia Coli, Klebsiella Pneumoniae, and Acinetobacter Baumannii. Pharmaceutics 2022, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Aljaghwani, A.; Alnuqaydan, A.M.; Khalilullah, H.; Younus, H.; El-Kady, A.M.; Aldakheel, F.M.; Khan, A.A.; et al. Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice. Biomedicines 2021, 9, 1673. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Partoazar, A.; Alaeddini, M.; Etemad-Moghadam, S.; Bahador, A. Photodisinfection Effects of Silver Sulfadiazine Nanoliposomes Doped-Curcumin on Acinetobacter Baumannii: A Mouse Model. Nanomedicine 2020, 15, 437–452. [Google Scholar] [CrossRef]
- Ekonomou, S.I.; Akshay Thanekar, P.; Lamprou, D.A.; Weaver, E.; Doran, O.; Stratakos, A.C. Development of Geraniol-Loaded Liposomal Nanoformulations against Salmonella Colonization in the Pig Gut. J. Agric. Food Chem. 2022, 70, 7004–7014. [Google Scholar] [CrossRef]
- Engel, J.B.; Heckler, C.; Tondo, E.C.; Daroit, D.J.; da Silva Malheiros, P. Antimicrobial Activity of Free and Liposome-Encapsulated Thymol and Carvacrol against Salmonella and Staphylococcus Aureus Adhered to Stainless Steel. Int. J. Food Microbiol. 2017, 252, 18–23. [Google Scholar] [CrossRef]
- Ye, T.; Sun, S.; Sugianto, T.D.; Tang, P.; Parumasivam, T.; Chang, Y.K.; Astudillo, A.; Wang, S.; Chan, H.-K. Novel Combination Proliposomes Containing Tobramycin and Clarithromycin Effective against Pseudomonas Aeruginosa Biofilms. Int. J. Pharm. 2018, 552, 130–138. [Google Scholar] [CrossRef]
- Zerillo, L.; Polvere, I.; Varricchio, R.; Madera, J.R.; D’Andrea, S.; Voccola, S.; Franchini, I.; Stilo, R.; Vito, P.; Zotti, T. Antibiofilm and Repair Activity of Ozonated Oil in Liposome. Microb. Biotechnol. 2022, 15, 1422–1433. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, C.; Li, C.; Lin, L. Inhibition of Escherichia Coli O157:H7 Biofilm on Vegetable Surface by Solid Liposomes of Clove Oil. LWT 2020, 117, 108656. [Google Scholar] [CrossRef]
- Montefusco-Pereira, C.V.; Formicola, B.; Goes, A.; Re, F.; Marrano, C.A.; Mantegazza, F.; Carvalho-Wodarz, C.; Fuhrmann, G.; Caneva, E.; Nicotra, F.; et al. Coupling Quaternary Ammonium Surfactants to the Surface of Liposomes Improves Both Antibacterial Efficacy and Host Cell Biocompatibility. Eur. J. Pharm. Biopharm. 2020, 149, 12–20. [Google Scholar] [CrossRef]
- Ding, T.; Li, T.; Li, J. Impact of Curcumin Liposomes with Anti-Quorum Sensing Properties against Foodborne Pathogens Aeromonas Hydrophila and Serratia Grimesii. Microb. Pathog. 2018, 122, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Li, T.; Wang, Z.; Li, J. Curcumin Liposomes Interfere with Quorum Sensing System of Aeromonas Sobria and in Silico Analysis. Sci. Rep. 2017, 7, 8612. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Hu, F.; Hu, S.; Kong, M.; Feng, C.; Liu, Y.; Cheng, X.; Ji, Q.; Chen, X. PH-Activated Nanoparticles with Targeting for the Treatment of Oral Plaque Biofilm. J. Mater. Chem. B 2018, 6, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Zhou, Z.; Xu, Q.; Fan, C.; Wang, L.; Ren, H.; Xu, S.; Ji, Q.; Chen, X. A Novel PH-Responsive Quaternary Ammonium Chitosan-Liposome Nanoparticles for Periodontal Treatment. Int. J. Biol. Macromol. 2019, 129, 1113–1119. [Google Scholar] [CrossRef]
- Jubeh, B.; Breijyeh, Z.; Karaman, R. Resistance of Gram-Positive Bacteria to Current Antibacterial Agents and Overcoming Approaches. Molecules 2020, 25, 2888. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.-F.; Alarcon, E.I. Bacterial Biofilm Formation on Implantable Devices and Approaches to Its Treatment and Prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef]
- Oliva, A.; Stefani, S.; Venditti, M.; Di Domenico, E.G. Biofilm-Related Infections in Gram-Positive Bacteria and the Potential Role of the Long-Acting Agent Dalbavancin. Front. Microbiol. 2021, 12, 749685. [Google Scholar] [CrossRef]
- Vazquez-Guillamet, C.; Kollef, M.H. Treatment of Gram-Positive Infections in Critically Ill Patients. BMC Infect. Dis. 2014, 14, 92. [Google Scholar] [CrossRef]
- Lebeaux, D.; Ghigo, J.-M.; Beloin, C. Biofilm-Related Infections: Bridging the Gap between Clinical Management and Fundamental Aspects of Recalcitrance toward Antibiotics. Microbiol Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef]
- Römling, U.; Balsalobre, C. Biofilm Infections, Their Resilience to Therapy and Innovative Treatment Strategies. J. Intern. Med. 2012, 272, 541–561. [Google Scholar] [CrossRef]
- Cha, J.-O.; Yoo, J.I.; Yoo, J.S.; Chung, H.-S.; Park, S.-H.; Kim, H.S.; Lee, Y.S.; Chung, G.T. Investigation of Biofilm Formation and Its Association with the Molecular and Clinical Characteristics of Methicillin-Resistant Staphylococcus Aureus. Osong Public Health Res. Perspect. 2013, 4, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Leshem, T.; Schnall, B.-S.; Azrad, M.; Baum, M.; Rokney, A.; Peretz, A. Incidence of Biofilm Formation among MRSA and MSSA Clinical Isolates from Hospitalized Patients in Israel. J. Appl. Microbiol. 2022, 133, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Neyra, R.C.; Frisancho, J.A.; Rinsky, J.L.; Resnick, C.; Carroll, K.C.; Rule, A.M.; Ross, T.; You, Y.; Price, L.B.; Silbergeld, E.K. Multidrug-Resistant and Methicillin-Resistant Staphylococcus Aureus (MRSA) in Hog Slaughter and Processing Plant Workers and Their Community in North Carolina (USA). Environ. Health Perspect. 2014, 122, 471–477. [Google Scholar] [CrossRef]
- Piechota, M.; Kot, B.; Frankowska-Maciejewska, A.; Grużewska, A.; Woźniak-Kosek, A. Biofilm Formation by Methicillin-Resistant and Methicillin-Sensitive Staphylococcus Aureus Strains from Hospitalized Patients in Poland. BioMed Res. Int. 2018, 2018, 4657396. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Peng, Y.; Su, M.; Zhu, S.; Pan, J.; Shen, B.; Duan, Y.; Huang, Y. Platensimycin-Encapsulated Liposomes or Micelles as Biosafe Nanoantibiotics Exhibited Strong Antibacterial Activities against Methicillin-Resistant Staphylococcus Aureus Infection in Mice. Mol. Pharm. 2020, 17, 2451–2462. [Google Scholar] [CrossRef]
- Allegrone, G.; Ceresa, C.; Rinaldi, M.; Fracchia, L. Diverse Effects of Natural and Synthetic Surfactants on the Inhibition of Staphylococcus Aureus Biofilm. Pharmaceutics 2021, 13, 1172. [Google Scholar] [CrossRef]
- Bhatia, E.; Sharma, S.; Jadhav, K.; Banerjee, R. Combinatorial Liposomes of Berberine and Curcumin Inhibit Biofilm Formation and Intracellular Methicillin Resistant Staphylococcus Aureus Infections and Associated Inflammation. J. Mater. Chem. B 2021, 9, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Tigabu, B.; Getachew, A. Treatment of Antibiotic-Resistant Bacteria by Nanoparticles: Current Approaches and Prospects. Ann Adv Chem 2022, 6, 001–009. [Google Scholar] [CrossRef]
- Inamuddin; Ahamed, M.I.; Prasad, R. (Eds.) Microbial Biosurfactants: Preparation, Properties and Applications; Environmental and Microbial Biotechnology; Springer: Singapore, 2021; ISBN 9789811566066. [Google Scholar]
- Giordani, B.; Costantini, P.E.; Fedi, S.; Cappelletti, M.; Abruzzo, A.; Parolin, C.; Foschi, C.; Frisco, G.; Calonghi, N.; Cerchiara, T.; et al. Liposomes Containing Biosurfactants Isolated from Lactobacillus Gasseri Exert Antibiofilm Activity against Methicillin Resistant Staphylococcus Aureus Strains. Eur. J. Pharm. Biopharm. 2019, 139, 246–252. [Google Scholar] [CrossRef]
- Omolo, C.A.; Hassan, D.; Devnarain, N.; Jaglal, Y.; Mocktar, C.; Kalhapure, R.S.; Jadhav, M.; Govender, T. Formulation of PH Responsive Multilamellar Vesicles for Targeted Delivery of Hydrophilic Antibiotics. Colloids Surf. B Biointerfaces 2021, 207, 112043. [Google Scholar] [CrossRef]
- Alavi, S.E.; Koohi, M.E.M.; Raza, A.; Adelnia, H.; Ebrahimi Shahmabadi, H. PEG-Grafted Liposomes for Enhanced Antibacterial and Antibiotic Activities: An in Vivo Study. NanoImpact 2022, 25, 100384. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Pinto, S.N.; Aires-da-Silva, F.; Bettencourt, A.; Aguiar, S.I.; Gaspar, M.M. Liposomes as a Nanoplatform to Improve the Delivery of Antibiotics into Staphylococcus Aureus Biofilms. Pharmaceutics 2021, 13, 321. [Google Scholar] [CrossRef]
- Sulahian, T.H.; Imrich, A.; DeLoid, G.; Winkler, A.R.; Kobzik, L. Signaling Pathways Required for Macrophage Scavenger Receptor-Mediated Phagocytosis: Analysis by Scanning Cytometry. Respir. Res. 2008, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, S.; Dou, H.; Wu, W.; Liu, Q.; Zhang, L.; Shen, Y.; Shu, G.; Yuan, Z.; Lin, J.; et al. Preparation of Cefquinome Sulfate Cationic Proliposome and Evaluation of Its Efficacy on Staphylococcus Aureus Biofilm. Colloids Surf. B Biointerfaces 2019, 182, 110323. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.V.; Nirwane, A.M.; Belubbi, T.; Nagarsenker, M.S. Pulmonary Delivery of Synergistic Combination of Fluoroquinolone Antibiotic Complemented with Proteolytic Enzyme: A Novel Antimicrobial and Antibiofilm Strategy. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2371–2384. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, Y.; Wang, L.; Guo, J.; Liu, W.; Meng, G.; Zhang, L.; Li, M.; Cong, L.; Sun, M. Recent Insights into the Prognostic and Therapeutic Applications of Lysozymes. Front. Pharmacol. 2021, 12, 767642. [Google Scholar] [CrossRef]
- Gupta, P.V.; Nirwane, A.M.; Nagarsenker, M.S. Inhalable Levofloxacin Liposomes Complemented with Lysozyme for Treatment of Pulmonary Infection in Rats: Effective Antimicrobial and Antibiofilm Strategy. AAPS PharmSciTech 2018, 19, 1454–1467. [Google Scholar] [CrossRef]
- Alvi, S.B.; Rajalakshmi, P.S.; Jogdand, A.; Sanjay, A.Y.; John, R.; Rengan, A.K. Iontophoresis Mediated Localized Delivery of Liposomal Gold Nanoparticles for Photothermal and Photodynamic Therapy of Acne. Biomater. Sci. 2021, 9, 1421–1430. [Google Scholar] [CrossRef]
- Portilla, S.; Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Encapsulation of the Antistaphylococcal Endolysin LysRODI in PH-Sensitive Liposomes. Antibiotics 2020, 9, 242. [Google Scholar] [CrossRef]
- Rout, B.; Liu, C.-H.; Wu, W.-C. Photosensitizer in Lipid Nanoparticle: A Nano-Scaled Approach to Antibacterial Function. Sci. Rep. 2017, 7, 7892. [Google Scholar] [CrossRef]
- Raj, V.; Kim, Y.; Kim, Y.-G.; Lee, J.-H.; Lee, J. Chitosan-Gum Arabic Embedded Alizarin Nanocarriers Inhibit Biofilm Formation of Multispecies Microorganisms. Carbohydr. Polym. 2022, 284, 118959. [Google Scholar] [CrossRef] [PubMed]
- Bazrgari, F.; Khameneh, B.; Bazzaz, B.S.F.; Mahmoudi, A.; Bizhan, M.-N. Effect of the Nanoliposomal Formulations of Rifampin and N-Acetyl Cysteine on Staphylococcus Epidermidis Biofilm. Nanomed. J. 2020, 7, 131–137. [Google Scholar]
- Eroğlu, İ.; Aslan, M.; Yaman, Ü.; Gultekinoglu, M.; Çalamak, S.; Kart, D.; Ulubayram, K. Liposome-Based Combination Therapy for Acne Treatment. J. Liposome Res. 2020, 30, 263–273. [Google Scholar] [CrossRef]
- Natsaridis, E.; Gkartziou, F.; Mourtas, S.; Stuart, M.C.A.; Kolonitsiou, F.; Klepetsanis, P.; Spiliopoulou, I.; Antimisiaris, S.G. Moxifloxacin Liposomes: Effect of Liposome Preparation Method on Physicochemical Properties and Antimicrobial Activity against Staphylococcus Epidermidis. Pharmaceutics 2022, 14, 370. [Google Scholar] [CrossRef]
- Gerayelou, G.; Khameneh, B.; Malaekeh-Nikouei, B.; Mahmoudi, A.; Fazly Bazzaz, B.S. Dual Antibiotic and Diffusible Signal Factor Combination Nanoliposomes for Combating Staphylococcus Epidermidis Biofilm. Adv. Pharm. Bull. 2020, 11, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Francolini, I.; Giansanti, L.; Piozzi, A.; Altieri, B.; Mauceri, A.; Mancini, G. Glucosylated Liposomes as Drug Delivery Systems of Usnic Acid to Address Bacterial Infections. Colloids Surf. B Biointerfaces 2019, 181, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Vögeling, H.; Plenagl, N.; Seitz, B.S.; Duse, L.; Pinnapireddy, S.R.; Dayyoub, E.; Jedelska, J.; Brüßler, J.; Bakowsky, U. Synergistic Effects of Ultrasound and Photodynamic Therapy Leading to Biofilm Eradication on Polyurethane Catheter Surfaces Modified with Hypericin Nanoformulations. Mater. Sci. Eng. C 2019, 103, 109749. [Google Scholar] [CrossRef] [PubMed]
- Plenagl, N.; Seitz, B.S.; Duse, L.; Pinnapireddy, S.R.; Jedelska, J.; Brüßler, J.; Bakowsky, U. Hypericin Inclusion Complexes Encapsulated in Liposomes for Antimicrobial Photodynamic Therapy. Int. J. Pharm. 2019, 570, 118666. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus Pneumoniae: Transmission, Colonization and Invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef]
- Chao, Y.; Marks, L.R.; Pettigrew, M.M.; Hakansson, A.P. Streptococcus Pneumoniae Biofilm Formation and Dispersion during Colonization and Disease. Front. Cell. Infect. Microbiol. 2015, 4, 194. [Google Scholar] [CrossRef]
- Silva, M.D.; Paris, J.L.; Gama, F.M.; Silva, B.F.B.; Sillankorva, S. Sustained Release of a Streptococcus Pneumoniae Endolysin from Liposomes for Potential Otitis Media Treatment. ACS Infect. Dis. 2021, 7, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Love, M.; Bhandari, D.; Dobson, R.; Billington, C. Potential for Bacteriophage Endolysins to Supplement or Replace Antibiotics in Food Production and Clinical Care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Ingendoh-Tsakmakidis, A.; Eberhard, J.; Falk, C.S.; Stiesch, M.; Winkel, A. In Vitro Effects of Streptococcus Oralis Biofilm on Peri-Implant Soft Tissue Cells. Cells 2020, 9, 1226. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto-Nakano, M. Role of Streptococcus Mutans Surface Proteins for Biofilm Formation. Jpn. Dent. Sci. Rev. 2018, 54, 22–29. [Google Scholar] [CrossRef]
- Fang, J.-Y.; Chou, W.-L.; Lin, C.-F.; Sung, C.T.; Alalaiwe, A.; Yang, S.-C. Facile Biofilm Penetration of Cationic Liposomes Loaded with DNase I/Proteinase K to Eradicate Cutibacterium Acnes for Treating Cutaneous and Catheter Infections. Int. J. Nanomed. 2021, 16, 8121–8138. [Google Scholar] [CrossRef]
- Drost, M.; Diamanti, E.; Fuhrmann, K.; Goes, A.; Shams, A.; Haupenthal, J.; Koch, M.; Hirsch, A.K.H.; Fuhrmann, G. Bacteriomimetic Liposomes Improve Antibiotic Activity of a Novel Energy-Coupling Factor Transporter Inhibitor. Pharmaceutics 2021, 14, 4. [Google Scholar] [CrossRef]
- Blanchard, J.D.; Elias, V.; Cipolla, D.; Gonda, I.; Bermudez, L.E. Effective Treatment of Mycobacterium Avium Subsp. Hominissuis and Mycobacterium Abscessus Species Infections in Macrophages, Biofilm, and Mice by Using Liposomal Ciprofloxacin. Antimicrob. Agents Chemother. 2018, 62, e00440-18. [Google Scholar] [CrossRef]
- Zhang, J.; Leifer, F.; Rose, S.; Chun, D.Y.; Thaisz, J.; Herr, T.; Nashed, M.; Joseph, J.; Perkins, W.R.; DiPetrillo, K. Amikacin Liposome Inhalation Suspension (ALIS) Penetrates Non-Tuberculous Mycobacterial Biofilms and Enhances Amikacin Uptake into Macrophages. Front. Microbiol. 2018, 9, 915. [Google Scholar] [CrossRef]
- Pu, C.; Tang, W. The Antibacterial and Antibiofilm Efficacies of a Liposomal Peptide Originating from Rice Bran Protein against Listeria Monocytogenes. Food Funct. 2017, 8, 4159–4169. [Google Scholar] [CrossRef]
- Ruiz, A.; Herráez, M.; Costa-Gutierrez, S.B.; Molina-Henares, M.A.; Martínez, M.J.; Espinosa-Urgel, M.; Barriuso, J. The Architecture of a Mixed Fungal–Bacterial Biofilm Is Modulated by Quorum-sensing Signals. Environ. Microbiol. 2021, 23, 2433–2447. [Google Scholar] [CrossRef]
- Allkja, J.; Azevedo, A.S. Characterization of Social Interactions and Spatial Arrangement of Individual Bacteria in MultiStrain or Multispecies Biofilm Systems Using Nucleic Acid Mimics-Fluorescence in situ Hybridization. In Fluorescence In Situ Hybridization (FISH) for Microbial Cells; Azevedo, N.F., Almeida, C., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2246, pp. 97–109. ISBN 978-1-07-161114-2. [Google Scholar]
- Chitlapilly Dass, S.; Bosilevac, J.M.; Weinroth, M.; Elowsky, C.G.; Zhou, Y.; Anandappa, A.; Wang, R. Impact of Mixed Biofilm Formation with Environmental Microorganisms on E. Coli O157:H7 Survival against Sanitization. npj Sci. Food. 2020, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Alrbyawi, H.; Poudel, I.; Annaji, M.; Arnold, R.D.; Tiwari, A.K.; Babu, R.J. Recent Advancements of Stimuli-Responsive Targeted Liposomal Formulations for Cancer Drug Delivery. Pharm. Nanotechnol. 2022, 10, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Estelrich, J.; Busquets, M. Iron Oxide Nanoparticles in Photothermal Therapy. Molecules 2018, 23, 1567. [Google Scholar] [CrossRef] [PubMed]
- Shipunova, V.O.; Belova, M.M.; Kotelnikova, P.A.; Shilova, O.N.; Mirkasymov, A.B.; Danilova, N.V.; Komedchikova, E.N.; Popovtzer, R.; Deyev, S.M.; Nikitin, M.P. Photothermal Therapy with HER2-Targeted Silver Nanoparticles Leading to Cancer Remission. Pharmaceutics 2022, 14, 1013. [Google Scholar] [CrossRef]
- Sun, P.; Ye, L.; Tan, X.; Peng, J.; Zhao, L.; Zhou, Y. Silver Nanoparticle-Assisted Photodynamic Therapy for Biofilm Eradication. ACS Appl. Nano Mater. 2022, 5, 8251–8259. [Google Scholar] [CrossRef]

| Liposome Type | Description | Advantages | Limitations | Refs. |
|---|---|---|---|---|
| Conventional (cationic, anionic, neutral) | Bilayer composed of neutral, positively or negatively charged phospholipids | Reduced drug-associated toxicity | Rapid removal from circulation | [41,42] |
| Deformable (elastic) | Elastic lipid bilayer | Deeper skin penetration for topical applications | Cargo release during prolonged storage times | [47] |
| Stealth | PEG-coated liposomes | Prolonged circulation time | Non-biodegradability of high molar mass PEG and toxicity of low molar mass PEG | [42,48] |
| Targeted | Liposomes surface modified with targeting moieties such as antibodies | Specificity | Rapid elimination from the bloodstream | [54] |
| Stimuli-responsive | Liposomes change conformation and release encapsulated cargo in response to external and internal triggers | Reduced off-target toxicity | * | [43,63,64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhlouf, Z.; Ali, A.A.; Al-Sayah, M.H. Liposomes-Based Drug Delivery Systems of Anti-Biofilm Agents to Combat Bacterial Biofilm Formation. Antibiotics 2023, 12, 875. https://doi.org/10.3390/antibiotics12050875
Makhlouf Z, Ali AA, Al-Sayah MH. Liposomes-Based Drug Delivery Systems of Anti-Biofilm Agents to Combat Bacterial Biofilm Formation. Antibiotics. 2023; 12(5):875. https://doi.org/10.3390/antibiotics12050875
Chicago/Turabian StyleMakhlouf, Zinb, Amaal Abdulraqeb Ali, and Mohammad Hussein Al-Sayah. 2023. "Liposomes-Based Drug Delivery Systems of Anti-Biofilm Agents to Combat Bacterial Biofilm Formation" Antibiotics 12, no. 5: 875. https://doi.org/10.3390/antibiotics12050875
APA StyleMakhlouf, Z., Ali, A. A., & Al-Sayah, M. H. (2023). Liposomes-Based Drug Delivery Systems of Anti-Biofilm Agents to Combat Bacterial Biofilm Formation. Antibiotics, 12(5), 875. https://doi.org/10.3390/antibiotics12050875