

Evaluation of Efflux-Mediated Resistance and Biofilm formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections

Abstract
1. Introduction
2. Results
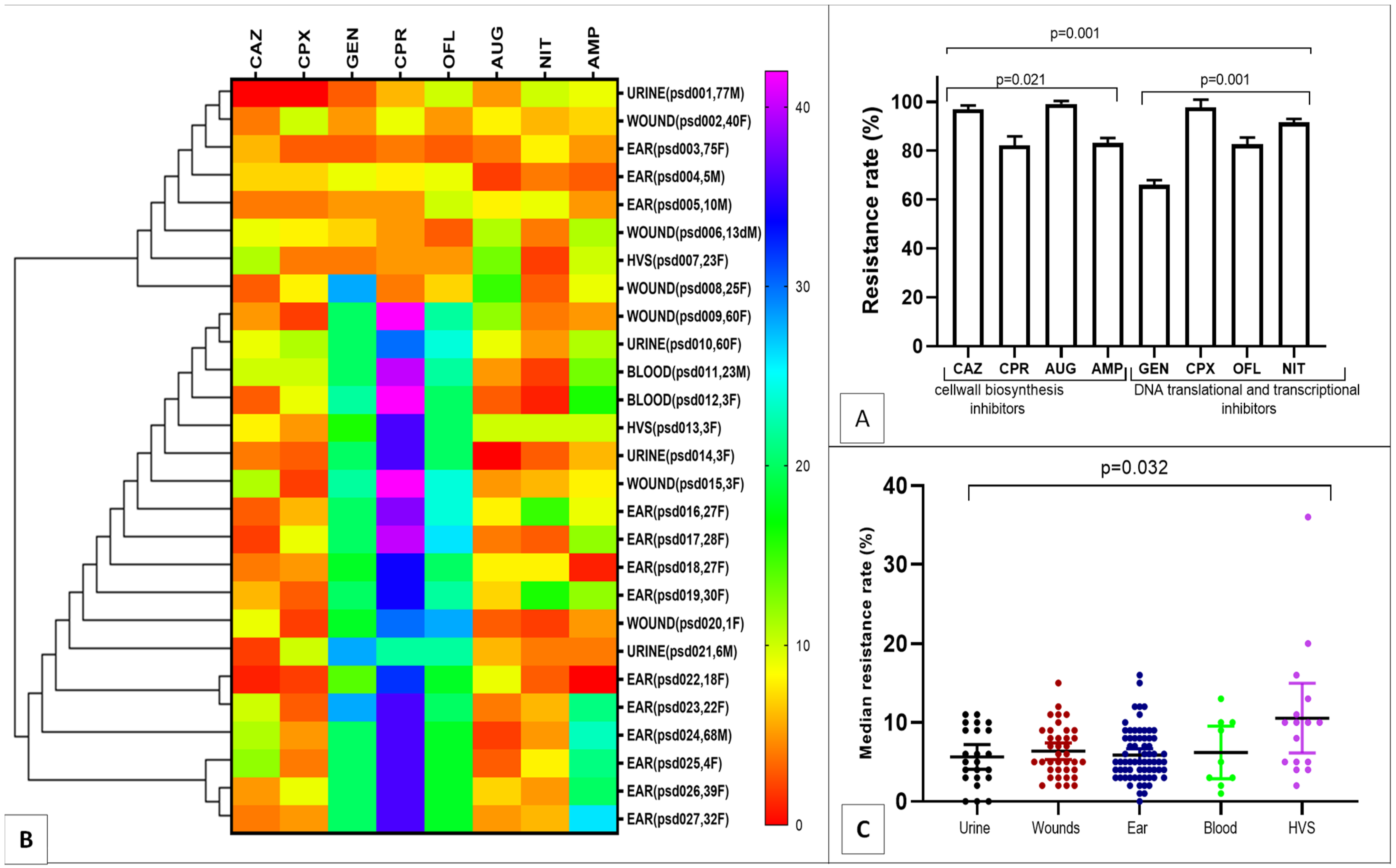
2.1. MDR Pseudomonas aeruginosa Resistance Pattern
2.2. Risk Factors for MDR Pseudomonas aeruginosa Infection
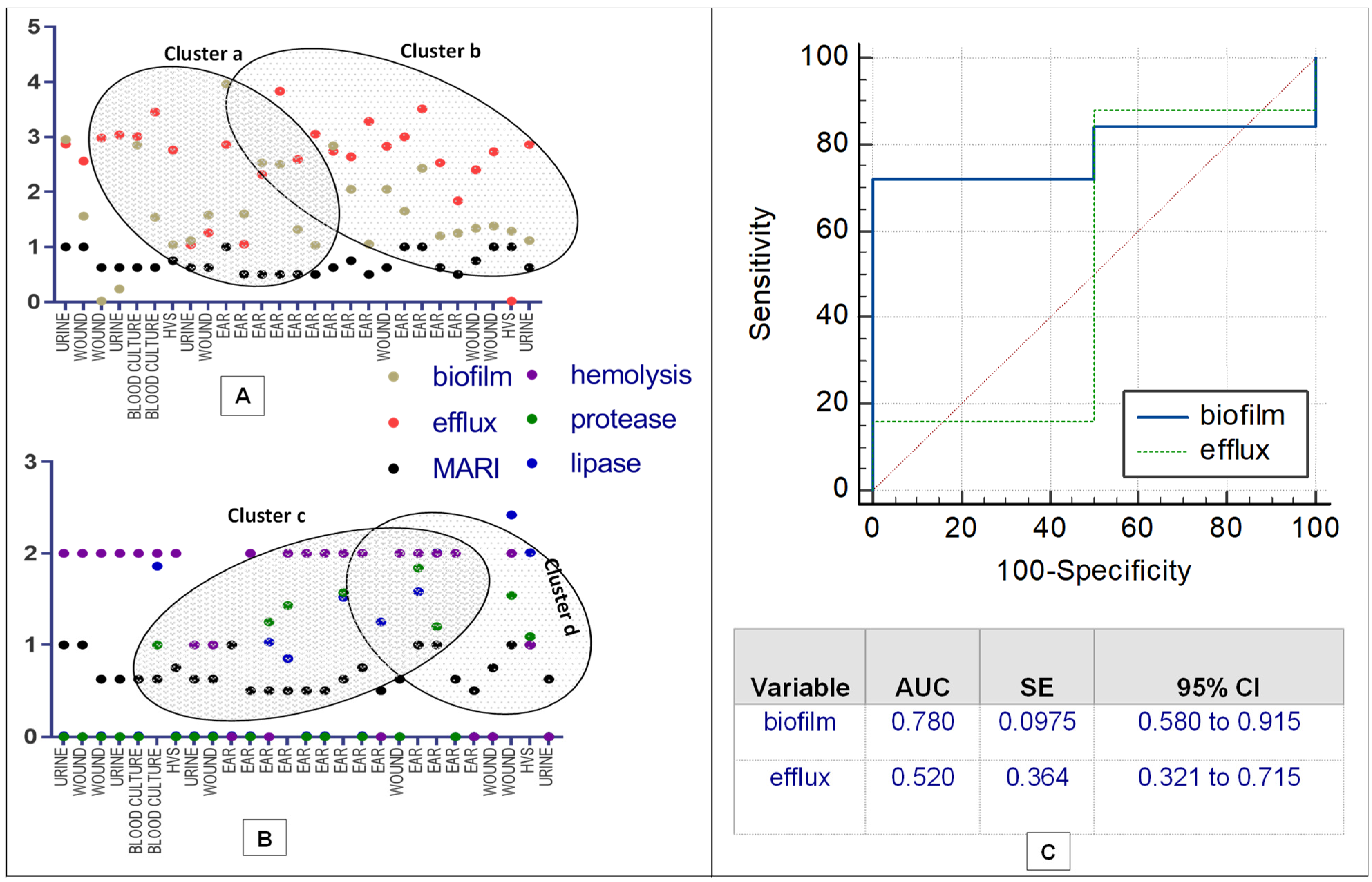
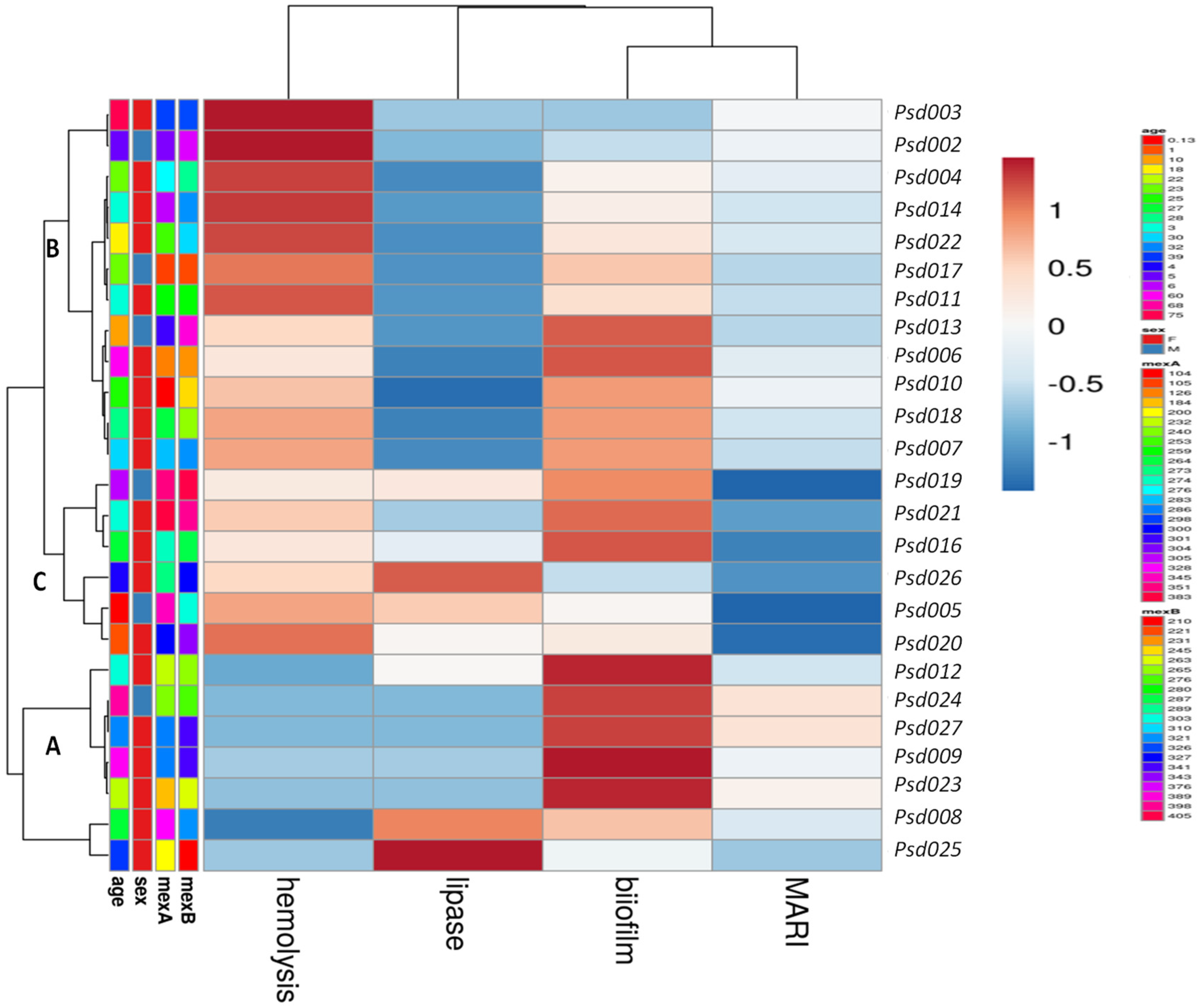
2.3. Phenotypic Virulence Factor Expression
2.4. Correlation and Prediction Performance of EP Activity and Expressed Biofilm
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain Collections
4.2. Antimicrobial Susceptibility
4.3. Biofilm Assay
4.4. Phenotypic Detection of Strain Efflux Pump (EP) Activity
4.5. Phenotypic Detection of Virulence Factors
4.6. Efflux Gene Genotyping
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Rojek, S.; Gozdzik, W.; Duszynska, W. Pseudomonas aeruginosa device associated–healthcare associated infections and its multidrug resistance at intensive care unit of University Hospital: Polish, 8.5-year, prospective, single-centre study. BMC Infect. Dis. 2021, 21, 180. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, O.; Lamikanra, A. Pattern of Esbls in Uro-Pathogens Obtained from a Nigerian Tertiary Hospital. Niger. J. Pharm. Res. 2021, 16, 139–147. [Google Scholar] [CrossRef]
- Araos, R.; D’Agata, E. Pseudomonas aeruginosa and Other Pseudomonas Species. In Mandell, Douglas, and Benett’s Principles and Practice of Infectious Diseases; Churchill Livingstone Elsevier: Philadelphia, PA, USA, 2019; pp. 2686–2699. [Google Scholar]
- Herrera, S.; Bodro, M.; Soriano, A. Predictors of multidrug resistant Pseudomonas aeruginosa involvement in bloodstream infections. Curr. Opin. Infect. Dis. 2021, 34, 686–692. [Google Scholar] [CrossRef]
- Langendonk, R.F.; Neill, D.R.; Fothergill, J.L. The building blocks of antimicrobial resistance in Pseudomonas aeruginosa: Implications for current resistance-breaking therapies. Front. Cell. Infect. Microbiol. 2021, 11, 307. [Google Scholar] [CrossRef]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum sensing mediated pathogenicity in Pseudomonas aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Zahedani, S.S.; Tahmasebi, H.; Jahantigh, M. Coexistence of Virulence Factors and Efflux Pump Genes in Clinical Isolates of Pseudomonas aeruginosa: Analysis of Biofilm-Forming Strains from Iran. Int. J. Microbiol. 2021, 2021, 5557361. [Google Scholar] [CrossRef]
- Rahbar, M.; Hamidi-Farahani, R.; Asgari, A.; Esmailkhani, A.; Soleiman-Meigooni, S. Expression of RND efflux pumps mediated antibiotic resistance in Pseudomonas aeruginosa clinical strains. Microb. Pathog. 2021, 153, 104789. [Google Scholar]
- Mohanty, S.; Baliyarsingh, B.; Nayak, S.K. Antimicrobial Resistance in Pseudomonas aeruginosa: A Concise. Antimicrob. Resist. A One Health Perspect. 2021, 49, 177–192. [Google Scholar]
- Palaniappan, B.; Solomon, A.P. Targeting AgrA quorum sensing regulator by bumetanide attenuates virulence in Staphylococcus aureus–A drug repurposing approach. Life Sci. 2021, 273, 119306. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Dawson, D., III; Emecen-Huja, P.; Nagarajan, R.; Howard, K.; Grady, M.E.; Gonzalez, O.A. The periodontal war: Microbes and immunity. Periodontology 2017, 75, 52–115. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.E.; Martinez, J.J. Modulation of host lipid pathways by pathogenic intracellular bacteria. Pathogens 2020, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Angus, A.A.; Lee, A.A.; Augustin, D.K.; Lee, E.J.; Evans, D.J.; Fleiszig, S.M. Pseudomonas aeruginosa induces membrane blebs in epithelial cells, which are utilized as a niche for intracellular replication and motility. Infect. Immun. 2008, 76, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Cianciotto, N.P.; White, R.C. Expanding role of type II secretion in bacterial pathogenesis and beyond. Infect. Immun. 2017, 85, e00014-17. [Google Scholar] [CrossRef] [PubMed]
- Olayinka, A.T.; Onile, B.A.; Olayinka, B.O. Prevalence of multi-drug resistant (MDR) Pseudomonas aeruginosa isolates in surgical units of Ahmadu Bello University Teaching Hospital, Zaria, Nigeria: An indication for effective control measures. Ann. Afr. Med. 2004, 3, 13–16. [Google Scholar]
- Omoregie, R.; Erebor, J.O.; Ahonkhai, I.; Isibor, J.O.; Ogefere, H.O. Observed changes in the prevalence of uropathogens in Benin City, Nigeria. N. Z. J. Med. Lab. Sci. 2008, 62, 29–31. [Google Scholar]
- Agwu, E.; Lhongbe, J.; Inyang, N. Prevalence of Quinolone-susceptible Pseudomonas aeruginosa and Staphylococcus aureus in Delayed-healing DFU’s in Ekpoma, Nigeria. Int. Wound J. 2010, 22, 100–105. [Google Scholar]
- Garba, I.; Lusa, Y.H.; Bawa, E.; Tijjani, M.B.; Aliyu, M.S.; Zango, U.U.; Raji, M.I.O. Antibiotics susceptibility pattern of Pseudomonas aeruginosa isolated from wounds in patients attending Ahmadu Bello University Teaching Hospital, Zaria, Nigeria. Niger. J. Basic Appl. Sci. 2012, 20, 32–34. [Google Scholar]
- Zubair, K.O.; Iregbu, K.C. Resistance pattern and detection of metallo-beta-lactamase genes in clinical isolates of Pseudomonas aeruginosa in a central Nigeria tertiary hospital. Niger. J. Clin. Pract. 2018, 21, 176–182. [Google Scholar]
- Ugwuanyi, F.C.; Ajayi, A.; Ojo, D.A.; Adeleye, A.I.; Smith, S.I. Evaluation of efflux pump activity and biofilm formation in multidrug resistant clinical isolates of Pseudomonas aeruginosa isolated from a Federal Medical Center in Nigeria. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 11. [Google Scholar] [CrossRef]
- George, C.R.R.; Jeffery, H.E.; Lahra, M.M. Infection of Mother and Baby. Keeling’s Fetal Neonatal Pathol. 2022, 1, 207–245. [Google Scholar]
- Alshaikh, B.N.; Reyes Loredo, A.; Knauff, M.; Momin, S.; Moossavi, S. The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis. Nutrients 2022, 14, 145. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, K.; Khan, M.H. Antimicrobial Mechanisms and Mode of Actions of Nanoemulsion Against Drug-Resistant ESKAPE Pathogens. In Handbook of Research on Nanoemulsion Applications in Agriculture, Food, Health, and Biomedical Sciences; IGI Global: Hershey, PA, USA, 2022; pp. 142–168. [Google Scholar]
- da Cruz, D.G.; de Magalhães, R.F.; Padilha, G.A.; da Silva, M.C.; Braga, C.L.; Silva, A.R.; Silva, P.L. Impact of positive biphasic pressure during low and high inspiratory efforts in Pseudomonas aeruginosa-induced pneumonia. PLoS ONE 2021, 16, e0246891. [Google Scholar] [CrossRef] [PubMed]
- Gudiol, C.; Durà-Miralles, X.; Aguilar-Company, J.; Hernández-Jiménez, P.; Martínez-Cutillas, M.; Fernandez-Avilés, F.; Carratalà, J. Co-infections and superinfections complicating COVID-19 in cancer patients: A multicentre, international study. J. Infect. 2021, 83, 306–313. [Google Scholar] [CrossRef]
- Johnson, D.I. Beck. In Bacterial Pathogens and Their Virulence Factors; Springer: Cham, Switzerland, 2018; pp. 363–379. [Google Scholar]
- Mobarak-Qamsari, E.; Kasra-Kermanshahi, R.; Moosavi-Nejad, Z. Isolation and identification of a novel, lipase-producing bacterium, Pseudomnas aeruginosa KM110. Iran. J. Microbiol. 2011, 3, 92–98. [Google Scholar]
- Ali, N.M.; Rehman, S.; Mazhar, S.A.; Liaqat, I.; Mazhar, B. Pseudomonas aeruginosa-Associated Acute and Chronic Pulmonary Infections. In Pathogenic Bacteria; IntechOpen: London, UK, 2020. [Google Scholar]
- Winter, M.; Buckling, A.; Harms, K.; Johnsen, P.J.; Vos, M. Antimicrobial resistance acquisition via natural transformation: Context is everything. Curr. Opin. Microbiol. 2021, 64, 133–138. [Google Scholar] [CrossRef]
- Brzozowski, M.; Krukowska, Ż.; Galant, K.; Jursa-Kulesza, J.; Kosik-Bogacka, D. Genotypic characterisation and antimicrobial resistance of Pseudomonas aeruginosa strains isolated from patients of different hospitals and medical centres in Poland. BMC Infect. Dis. 2020, 20, 693. [Google Scholar] [CrossRef]
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef]
- Ohore, H.U.; Akinduti, P.A.; Ahuekwe, E.F.; Ajayi, A.S.; Olasehinde, G.I. Molecular Detection of ESBLs, TEM, SHV, and CTX-M in Clinical Pseudomonas aeruginosa Isolates in Ogun State. Bioenergy Biochem. Process. Technol. 2022, 127–136. [Google Scholar]
- Ferrand, A.; Vergalli, J.; Davin-Regli, A. An intertwined network of regulation controls membrane permeability Including drug influx and efflux in Enterobacteriaceae. Microorganisms 2020, 8, 833. [Google Scholar] [CrossRef]
- Ashwath, P.; Sannejal, A.D. A quest to the therapeutic arsenal: Novel strategies to combat multidrug-resistant bacteria. Curr. Gene Ther. 2022, 22, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ayepola, O.O.; Olasupo, N.A.; Egwari, L.O.; Schaumburg, F. Characterization of Panton–Valentine leukocidin-positive Staphylococcus aureus from skin and soft tissue infections and wounds in Nigeria: A cross-sectional study. F1000Research 2018, 7, 30345027. [Google Scholar] [CrossRef] [PubMed]
- da Cruz Nizer, W.S.; Inkovskiy, V.; Versey, Z.; Strempel, N.; Cassol, E.; Overhage, J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens 2021, 10, 1187. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, Sixteenth Informational Supplement; Document M100-S20; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- EUCAST Clinical Breakpoints and Dosing. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 5 August 2021).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Paterson, D.L. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Sadat, A.; El-Sherbiny, H.; Zakaria, A.; Ramadan, H.; Awad, A. Prevalence, antibiogram and virulence characterization of Vibrio isolates from fish and shellfish in Egypt: A possible zoonotic hazard to humans. J. Appl. Microbiol. 2020, 131, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.J.; Fatma, T.; Rathan, A. Detection of biofilm formation among the clinical isolates of Staphylococci: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–29. [Google Scholar] [CrossRef]
- Akinduti, A.P.; Osiyemi, J.A.; Banjo, T.T.; Ejilude, O.; El-Ashker, M.; Adeyemi, A.G.; Isibor, P.O. Clonal diversity and spatial dissemination of multi-antibiotics resistant Staphylococcus aureus pathotypes in Southwest Nigeria. PLoS ONE 2021, 16, e0247013. [Google Scholar] [CrossRef]
- Edberg, S.C.; Gallo, P.; Kontnick, C. Analysis of the virulence characteristics of bacteria isolated from bottled, water cooler, and tap water. Microb. Ecol. Health Dis. 1996, 9, 67–77. [Google Scholar] [CrossRef]
- Lopes, E.S.; Cardoso, W.M.; Nishi, D.M.; Horn, R.V.; Albuquerque, A.H.; Lima, S.V.G.; Beleza, A.J.F.; Gaio, F.C.; Carmo, C.; Pascoal Filho, M.N.; et al. Sero-group identification, phenotypic detection of hemolysis and extended spectrum beta-lactamases of Escherichia coli isolated from psittacine of illegal wildlife trade in Fortaleza, Brazil. Arq. Bras. Med. Vet. Zootec. 2018, 70, 823–829. [Google Scholar] [CrossRef]
- Suganthi, C.; Mageswari, A.; Karthikeyan, S.; Anbalagan, M.; Sivakumar, A.; Gothandam, K.M. Screening and optimization of protease production from a halotolerant Bacillus licheniformis isolated from saltern sediments. J. Genet. Eng. Biotechnol. 2013, 11, 47–52. [Google Scholar] [CrossRef]
- Banoth, L.; Devarapalli, K.; Paul, I.; Thete, K.N.; Pawar, S.V.; Banerjee, U.C. Screening, isolation and selection of a potent lipase producing microorganism and its use in the kinetic resolution of drug intermediates. J. Indian Chem. Soc. 2021, 98, 100143. [Google Scholar] [CrossRef]
- Su, F.; Wang, J. Berberine inhibits the MexXY-OprM efflux pump to reverse imipenem resistance in a clinical carbapenem-resistant Pseudomonas aeruginosa isolate in a planktonic state. Exp. Ther. Med. 2018, 15, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gupta, P.; Kumar, R.; Bhardwaj, A. dPABBs: A novel in silico approach for predicting and designing anti-biofilm peptides. Sci. Rep. 2016, 6, 21839. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | MDR-Psa n/N(%) | OR(CI) | p Value |
|---|---|---|---|
| Age (years) | |||
| Children (0–12) | 10/21(47.6) | ||
| Teenagers (13–19) | 1/15(6.7) | ||
| Adolescents (20–23) | 3/45(6.7) | 0.986(0.946–1.027) | 0.024 |
| Adults (24–50) | 8/32(25.0) | ||
| Elders (51–Above) | 5/29(17.2) | ||
| Gender | |||
| Female | 20/96(20.8) | 1.440(0.211–9.827) | 0.001 |
| Male | 7/51(13.7) | ||
| Diagnosis | |||
| Urinary tract infections | 4/36(11.1) | ||
| Wounds * | 6/28(21.4) | ||
| Blood stream infection | 2/11(18.2) | 0.860(0.438–1.688) | 0.002 |
| High vaginal infection | 2/25(8.0) | ||
| Otorhea infections | 13/35(37.1) | ||
| Others ** | 0/8(0.0) |
| Functional Activities | Otorrhea | Blood Stream Infection | UTI | Vaginal Infection | Wound Infection | 95% CI | p Value | |
|---|---|---|---|---|---|---|---|---|
| n(%) | ||||||||
| Biofilm | Producer | 13(48.2) | 2(7.4) | 3(11.1) | 2(7.4) | 5(18.5) | 0.820–1.032 | 0.001 |
| Efflux (EtBr (mg/L)) | Efflux(0.5) | 12(44.4) | 2(7.4) | 4(14.8) | 1(3.7) | 5(18.5) | 0.762–1.016 | 0.001 |
| Efflux(1.0) | 8(29.6) | 2(7.4) | 3(11.1) | 1(3.7) | 4(14.8) | 0.209–0.606 | 0.001 | |
| Efflux(1.5) | 8(29.6) | 2(7.4) | 1(3.7) | 0(0.0) | 0(0.0) | 0.477–0.857 | 0.001 | |
| Efflux(2.0) | 12(44.4) | 2(7.4) | 3(11.1) | 1(3.7) | 6(22.2) | 0.762–1.016 | 0.001 | |
| Virulence factor | Protease | 5(18.5) | 1(3.7) | 0(0.0) | 1(3.7) | 1(3.7) | 0.112–0.480 | 0.003 |
| Lipase | 11(40.7) | 2(7.4) | 2(7.4) | 2(7.4) | 3(11.1) | 0.143–0.523 | 0.001 | |
| Hemolysin | 9(33.3) | 2(7.4) | 2(7.4) | 1(3.7) | 6(22.2) | 1.109–1.780 | 0.001 | |
| Parameters | Multi-Antibiotic Resistance Index (MARI) | |||
|---|---|---|---|---|
| R Coefficient | SE | 95% CI | p Value | |
| efflux | −0.85 | 0.04511 | −0.1119–0.0743 | 0.6807 |
| biofilm | −0.08 | 0.04424 | −0.0226–0.1601 | 0.1334 |
| lipase | 0.36 | 0.09143 | −0.0800–0.2982 | 0.2448 |
| protease | 0.28 | 0.1168 | −0.2824–0.2009 | 0.7305 |
| hemolysis | 0.13 | 0.04520 | −0.0673–0.1198 | 0.5669 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinduti, P.A.; George, O.W.; Ohore, H.U.; Ariyo, O.E.; Popoola, S.T.; Adeleye, A.I.; Akinwande, K.S.; Popoola, J.O.; Rotimi, S.O.; Olufemi, F.O.; et al. Evaluation of Efflux-Mediated Resistance and Biofilm formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections. Antibiotics 2023, 12, 626. https://doi.org/10.3390/antibiotics12030626
Akinduti PA, George OW, Ohore HU, Ariyo OE, Popoola ST, Adeleye AI, Akinwande KS, Popoola JO, Rotimi SO, Olufemi FO, et al. Evaluation of Efflux-Mediated Resistance and Biofilm formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections. Antibiotics. 2023; 12(3):626. https://doi.org/10.3390/antibiotics12030626
Chicago/Turabian StyleAkinduti, Paul A., Onome W. George, Hannah U. Ohore, Olusegun E. Ariyo, Samuel T. Popoola, Adenike I. Adeleye, Kazeem S. Akinwande, Jacob O. Popoola, Solomon O. Rotimi, Fredrick O. Olufemi, and et al. 2023. "Evaluation of Efflux-Mediated Resistance and Biofilm formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections" Antibiotics 12, no. 3: 626. https://doi.org/10.3390/antibiotics12030626
APA StyleAkinduti, P. A., George, O. W., Ohore, H. U., Ariyo, O. E., Popoola, S. T., Adeleye, A. I., Akinwande, K. S., Popoola, J. O., Rotimi, S. O., Olufemi, F. O., Omonhinmin, C. A., & Olasehinde, G. I. (2023). Evaluation of Efflux-Mediated Resistance and Biofilm formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections. Antibiotics, 12(3), 626. https://doi.org/10.3390/antibiotics12030626