

Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. The Antimicrobial Activity of Selected Bacillus Strains
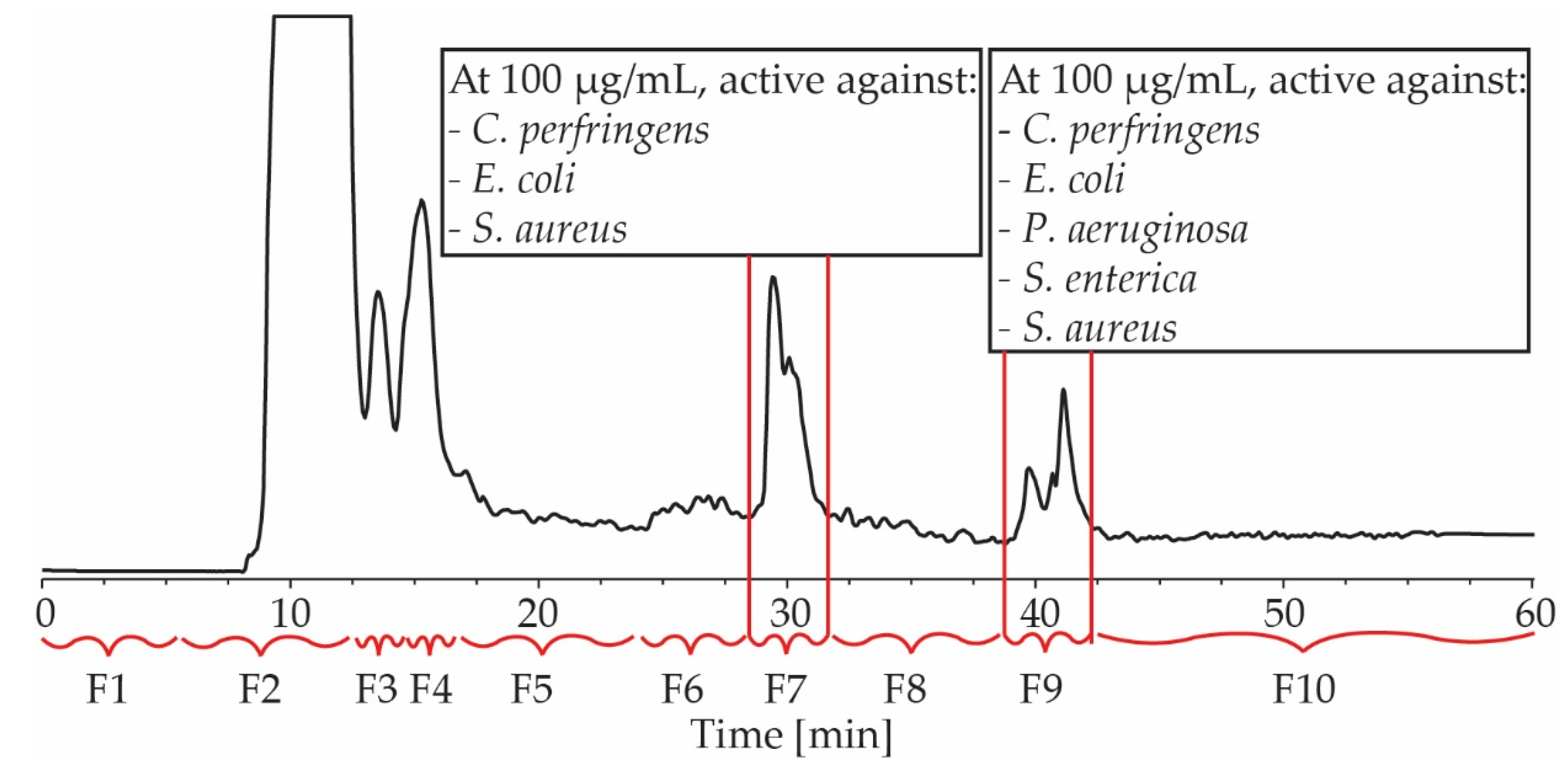
2.2. Isolation and Structure Elucidation of Antimicrobial Compounds
2.3. Animal Trial
2.4. Caecum Content Analysis
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedure
4.2. Bacillus Strains and Fermentation
4.3. Compound Isolation
4.3.1. Extraction
4.3.2. Isolation
4.3.3. Purification
- C16 surfactin C (1): white powder; [α]D23.0 +7.3 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S1; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S1–S5;
- C15 surfactin C (2): white powder; [α]D23.0 +7.0 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S2; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S6–S10;
- C14 surfactin C (3): white powder; [α]D23.0 +7.8 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S3; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S11–S15;
- C13 surfactin C (4): white solid; [α]D23.0 +8.1 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S4; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S16–S20;
- Maculosin (5): white solid; [α]D23.0 −20.1 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S5; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S21–S25;
- Maculosine 2 (6): white solid; [α]D23.0 −16.0 (c 0.1, MeOH); 1H NMR and 13C NMR data see Table S6; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S26–S30;
- Genistein (7): brown powder; 1H NMR and 13C NMR data see Table S7; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S31–S35;
- Daidzein (8): brown powder; 1H NMR and 13C NMR data see Table S8; LRESIMS, 1H, COSY, HSQC and HMBC NMR spectra see Figure S36–S40.
4.4. Antimicrobial Assay
4.5. The Animal Trial
4.6. Targeted Metabolomic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Callaway, T.R.; Lillehoj, H.; Chuanchuen, R.; Gay, C.G. Alternatives to Antibiotics: A Symposium on the Challenges and Solutions for Animal Health and Production. Antibiotics 2021, 10, 471. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fang, P.; Yi, X.; Kumar, V.; Peng, M. Probiotics Bacillus cereus and B. subtilis reshape the intestinal microbiota of Pengze crucian carp (Carassius auratus var. Pengze) fed with high plant protein diets. Front. Nutr. 2022, 9, 1027641. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well known antibiotics on gram-positive bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef]
- Fasina, Y.O.; Newman, M.M.; Stough, J.M.; Liles, M.R. Effect of Clostridium perfringens infection and antibiotic administration on microbiota in the small intestine of broiler chickens. Poult. Sci. 2016, 95, 247–260. [Google Scholar] [CrossRef]
- Kruse, S.; Schenk, M.; Pierre, F.; Morlock, G.E. Bacillus. subtilis spores in probiotic feed quantified via bacterial metabolite using planar chromatography. Anal. Chim. Acta 2022, 1221, 340124. [Google Scholar] [CrossRef]
- Feighner, S.D.; Dashkevicz, M.P. Subtherapeutic levels of antibiotics in poultry feeds and their effects on weight gain, feed efficiency, and bacterial cholyltaurine hydrolase activity. Appl. Environ. Microbiol. 1987, 53, 331–336. [Google Scholar] [CrossRef]
- Arsène, M.M.J.; Davares, A.K.L.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe production and as potential alternatives to antibiotics. Vet. World 2021, 14, 319–328. [Google Scholar] [CrossRef]
- Jeni, R.E.; Dittoe, D.K.; Olson, E.G.; Lourenco, J.; Corcionivoschi, N.; Ricke, S.C.; Callaway, T.R. Probiotics and potential applications for alternative poultry production systems. Poult. Sci. 2021, 100, 101156. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Joint FAO/WHO Working Group: London, ON, Canada, 2002; pp. 1–11. [Google Scholar]
- Nurmi, E.; Rantala, M. New aspects of Salmonella infection in broiler production. Nature 1973, 241, 210–211. [Google Scholar] [CrossRef]
- Yi, R.; Pan, Y.; Long, X.; Tan, F.; Zhao, X. Enzyme Producing Activity of Probiotics and Preparation of Compound Enzyme. J. Chem. 2020, 2020, 9140281. [Google Scholar] [CrossRef]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef]
- Nagpal, R.; Yadav, H.; Puniya, A.K.; Singh, K.; Jain, S.; Marotta, F. Potential of probiotics and prebiotics for synbiotic functional dairy foods: An overview. Int. J. Probiotics Prebiotic 2007, 2, 75. [Google Scholar]
- Yadav, H.; Jain, S.; Sinha, P.R. Oral administration of dahi containing probiotic Lactobacillus acidophilus and Lactobacillus casei delayed the progression of streptozotocin-induced diabetes in rats. J. Dairy Res. 2008, 75, 189–195. [Google Scholar] [CrossRef]
- Nalla, K.; Manda, N.K.; Dhillon, H.S.; Kanade, S.R.; Rokana, N.; Hess, M.; Puniya, A.K. Impact of Probiotics on Dairy Production Efficiency. Front. Microbiol. 2022, 13, 805963. [Google Scholar] [CrossRef]
- Anand, U.; Vaishnav, A.; Sharma, S.K.; Sahu, J.; Ahmad, S.; Sunita, K.; Suresh, S.; Dey, A.; Bontempi, E.; Singh, A.K.; et al. Current advances and research prospects for agricultural and industrial uses of microbial strains available in world collections. Sci. Total Environ. 2022, 842, 156641. [Google Scholar] [CrossRef]
- Maresca, D.; Zotta, T.; Mauriello, G. Adaptation to Aerobic Environment of Lactobacillus johnsonii/gasseri Strains. Front. Microbiol. 2018, 9, 157. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Zaineldin, A.I.; Hegazi, S.; Koshio, S.; Ishikawa, M.; Bakr, A.; El-Keredy, A.M.S.; Dawood, M.A.O.; Dossou, S.; Wang, W.; Yukun, Z. Bacillus subtilis as probiotic candidate for red sea bream: Growth performance, oxidative status, and immune response traits. Fish Shellfish Immunol. 2018, 79, 303–312. [Google Scholar] [CrossRef]
- Mazanko, M.S.; Popov, I.V.; Prazdnova, E.V.; Refeld, A.G.; Bren, A.B.; Zelenkova, G.A.; Chistyakov, V.A.; Algburi, A.; Weeks, R.M.; Ermakov, A.M.; et al. Beneficial Effects of Spore-Forming Bacillus Probiotic Bacteria Isolated From Poultry Microbiota on Broilers’ Health, Growth Performance, and Immune System. Front. Vet. Sci. 2022, 9, 877360. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Arias, S.; Chaignepain, S.; Denayrolles, M.; Schmitter, J.M.; Bressollier, P.; Urdaci, M.C. Identification of surface proteins involved in the adhesion of a probiotic Bacillus cereus strain to mucin and fibronectin. Microbiology 2009, 155, 1708–1716. [Google Scholar] [CrossRef] [PubMed]
- Biermann, R.; Niemeyer, L.; Rösner, L.; Ude, C.; Lindner, P.; Bice, I.; Beutel, S. Facilitated endospore detection for Bacillus spp. through automated algorithm-based image processing. Eng. Life Sci. 2022, 22, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Franz, C.M.; Ben Omar, N.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.I.; García, J.M.; Roget, D.; Ferrer, A.; Wieme, A.D.; Vandamme, P.; Rodríguez, S.; Llauradó, G.; Lescaylle, Y.; Peña, L.; et al. Isolation and Identification of a Bacillus sp. from Freshwater Sediment Displaying Potent Activity Against Bacteria and Phytopathogen Fungi. Curr. Microbiol. 2022, 79, 398. [Google Scholar] [CrossRef]
- Koistinen, V.M.; Hedberg, M.; Shi, L.; Johansson, A.; Savolainen, O.; Lehtonen, M.; Aura, A.-M.; Hanhineva, K.; Landberg, R. Metabolite Pattern Derived from Lactiplantibacillus plantarum—Fermented Rye Foods and In Vitro Gut Fermentation Synergistically Inhibits Bacterial Growth. Mol. Nutr. Food Res. 2022, 66, 2101096. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Rood, J.I.; Moore, R.J.; Titball, R.W. Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens. Trends Microbiol. 2009, 17, 32–36. [Google Scholar] [CrossRef]
- Arima, K.; Kakinuma, A.; Tamura, G. Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: Isolation, characterization and its inhibition of fibrin clot formation. Biochem. Biophys. Res. Commun. 1968, 31, 488–494. [Google Scholar] [CrossRef]
- Stierle, A.C.; Cardellina, J.H.; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternata. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef]
- Yap, A.C.; Chan, K.G.; Sim, K.S.; Choo, Y.M. A new oxolane from Enterobacter cloacae. Nat. Prod. Res. 2016, 30, 783–788. [Google Scholar] [CrossRef]
- Hong, H.; Landauer, M.R.; Foriska, M.A.; Ledney, G.D. Antibacterial activity of the soy isoflavone genistein. J. Basic Microbiol. 2006, 46, 329–335. [Google Scholar] [CrossRef]
- Roh, C.; Choi, K.-Y.; Pandey, B.P.; Kim, B.-G. Hydroxylation of daidzein by CYP107H1 from Bacillus subtilis 168. J. Mol. Catal. B Enzym. 2009, 59, 248–253. [Google Scholar] [CrossRef]
- Pang, Y.; Yang, J.; Chen, X.; Jia, Y.; Li, T.; Jin, J.; Liu, H.; Jiang, L.; Hao, Y.; Zhang, H.; et al. An Antifungal Chitosanase from Bacillus subtilis SH21. Molecules 2021, 26, 1863. [Google Scholar] [CrossRef]
- Pang, X.; Zhao, J.; Fang, X.; Liu, H.; Zhang, Y.; Cen, S.; Yu, L. Surfactin derivatives from Micromonospora sp. CPCC 202787 and their anti-HIV activities. J. Antibiot. 2017, 70, 105–108. [Google Scholar] [CrossRef]
- Ross 308/308; FF Broiler: Performance Objectives. Aviagen: Huntsville, AL, USA, 2022. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_Broiler/RossxRoss308-BroilerPerformanceObjectives2022-EN.pdf (accessed on 30 January 2023).
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Invernici, M.M.; Furlaneto, F.A.C.; Messora, M.R. Effectiveness of Multi-strain Versus Single-strain Probiotics: Current Status and Recommendations for the Future. J. Clin. Gastroenterol. 2018, 52, S35–S40. [Google Scholar] [CrossRef]
- Cochrane, S.A.; Findlay, B.; Bakhtiary, A.; Acedo, J.Z.; Rodriguez-Lopez, E.M.; Mercier, P.; Vederas, J.C. Antimicrobial lipopeptide tridecaptin A1 selectively binds to Gram-negative lipid II. Proc. Natl. Acad. Sci. USA 2016, 113, 11561–11566. [Google Scholar] [CrossRef]
- Sheppard, J.D.; Jumarie, C.; Cooper, D.G.; Laprade, R. Ionic channels induced by surfactin in planar lipid bilayer membranes. Biochim. Biophys. Acta 1991, 1064, 13–23. [Google Scholar] [CrossRef]
- Grau, A.; Ortiz, A.; de Godos, A.; Gómez-Fernández, J.C. A biophysical study of the interaction of the lipopeptide antibiotic iturin A with aqueous phospholipid bilayers. Arch. Biochem. Biophys. 2000, 377, 315–323. [Google Scholar] [CrossRef]
- Inès, M.; Dhouha, G. Lipopeptide surfactants: Production, recovery and pore forming capacity. Peptides 2015, 71, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Zhu, F.; Du, P.; Yang, X.; Qiu, D.; Yu, Z.; Chen, J.; Zhao, X. Lipopeptide induces apoptosis in fungal cells by a mitochondria-dependent pathway. Peptides 2010, 31, 1978–1986. [Google Scholar] [CrossRef] [PubMed]
- Ambroggio, E.E.; Separovic, F.; Bowie, J.H.; Fidelio, G.D.; Bagatolli, L.A. Direct visualization of membrane leakage induced by the antibiotic peptides: Maculatin, citropin, and aurein. Biophys. J. 2005, 89, 1874–1881. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Zhao, H.; Pang, B.; Qu, R.; Lian, Z.; Jiang, C.; Shao, D.; Huang, Q.; Jin, M.; Shi, J. Capability of iturin from Bacillus subtilis to inhibit Candida albicans in vitro and in vivo. Appl. Microbiol. Biotechnol. 2019, 103, 4377–4392. [Google Scholar] [CrossRef]
- Peypoux, F.; Besson, F.; Michel, G.; Delcambe, L. Preparation and antibacterial activity upon Micrococcus luteus of derivatives of iturin A, mycosubtilin and bacillomycin L, antibiotics from Bacillus subtilis. J. Antibiot. 1979, 32, 136–140. [Google Scholar] [CrossRef]
- Liu, J.; Li, W.; Zhu, X.; Zhao, H.; Lu, Y.; Zhang, C.; Lu, Z. Surfactin effectively inhibits Staphylococcus aureus adhesion and biofilm formation on surfaces. Appl. Microbiol. Biotechnol. 2019, 103, 4565–4574. [Google Scholar] [CrossRef]
- Steigenberger, J.; Verleysen, Y.; Geudens, N.; Martins, J.C.; Heerklotz, H. The Optimal Lipid Chain Length of a Membrane-Permeabilizing Lipopeptide Results From the Balance of Membrane Partitioning and Local Damage. Front. Microbiol. 2021, 12, 669709. [Google Scholar] [CrossRef]
- González-Jaramillo, L.M.; Aranda, F.J.; Teruel, J.A.; Villegas-Escobar, V.; Ortiz, A. Antimycotic activity of fengycin C biosurfactant and its interaction with phosphatidylcholine model membranes. Colloids. Surf. B Biointerfaces 2017, 156, 114–122. [Google Scholar] [CrossRef]
- González, O.; Ortíz-Castro, R.; Díaz-Pérez, C.; Díaz-Pérez, A.L.; Magaña-Dueñas, V.; López-Bucio, J.; Campos-García, J. Non-ribosomal Peptide Synthases from Pseudomonas aeruginosa Play a Role in Cyclodipeptide Biosynthesis, Quorum-Sensing Regulation, and Root Development in a Plant Host. Microb. Ecol. 2017, 73, 616–629. [Google Scholar] [CrossRef]
- Wang, X.; Feng, Z.; Li, C.; Cai, X.; Long, H.; Zhang, X.; Huang, A.; Zeng, Y.; Ren, W.; Xie, Z. Analysis of the Antioxidant Composition of Low Molecular Weight Metabolites from the Agarolytic Bacterium Alteromonas macleodii QZ9-9: Possibilities for High-Added Value Utilization of Macroalgae. Antioxidants 2022, 11, 1977. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, D.; Chen, M.; Zhang, Z.; Lian, X.-Y. A rare diketopiperazine glycoside from marine-sourced Streptomyces sp. ZZ446. Nat. Prod. Res. 2020, 34, 1046–1050. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Altuntas, E.G.; Juneja, V.K. Microbial Fermentation in Food Preservation. In Microbial Control and Food Preservation: Theory and Practice; Springer: New York, NY, USA, 2017; pp. 281–298. [Google Scholar]
- Polkowski, K.; Mazurek, A.P. Biological properties of genistein. A review of in vitro and in vivo data. Acta Pol. Pharm. 2000, 57, 135–155. [Google Scholar]
- Taylor, C.K.; Levy, R.M.; Elliott, J.C.; Burnett, B.P. The effect of genistein aglycone on cancer and cancer risk: A review of in vitro, preclinical, and clinical studies. Nutr. Rev. 2009, 67, 398–415. [Google Scholar] [CrossRef]
- Buchmann, D.; Schultze, N.; Borchardt, J.; Böttcher, I.; Schaufler, K.; Guenther, S. Synergistic antimicrobial activities of epigallocatechin gallate, myricetin, daidzein, gallic acid, epicatechin, 3-hydroxy-6-methoxyflavone and genistein combined with antibiotics against ESKAPE pathogens. J. Appl. Microbiol. 2022, 132, 949–963. [Google Scholar] [CrossRef]
- Verdrengh, M.; Collins, L.V.; Bergin, P.; Tarkowski, A. Phytoestrogen genistein as an anti-staphylococcal agent. Microb. Infect. 2004, 6, 86–92. [Google Scholar] [CrossRef]
- Yamagishi, T.; Otsuka, E.; Hagiwara, H. Reciprocal control of expression of mRNAs for osteoclast differentiation factor and OPG in osteogenic stromal cells by genistein: Evidence for the involvement of topoisomerase II in osteoclastogenesis. Endocrinology 2001, 142, 3632–3637. [Google Scholar] [CrossRef]
- Chen, P.; Sun, J.; Liang, Z.; Xu, H.; Du, P.; Li, A.; Meng, Y.; Reshetnik, E.I.; Liu, L.; Li, C. The bioavailability of soy isoflavones in vitro and their effects on gut microbiota in the simulator of the human intestinal microbial ecosystem. Food Res. Int. 2022, 152, 110868. [Google Scholar] [CrossRef]
- Medina, B.; Girard, I.D.; Jacotot, E.; Julliand, V. Effect of a preparation of Saccharomyces cerevisiae on microbial profiles and fermentation patterns in the large intestine of horses fed a high fiber or a high starch diet. J. Anim. Sci. 2002, 80, 2600–2609. [Google Scholar] [CrossRef]
- Cavazzoni, V.; Adami, A.; Castrovilli, C. Performance of broiler chickens supplemented with Bacillus coagulans as probiotic. Br. Poult. Sci. 1998, 39, 526–529. [Google Scholar] [CrossRef]
- Spiehs, M.J.; Varel, V.H. Nutrient excretion and odorant production in manure from cattle fed corn wet distillers grains with solubles. J. Anim. Sci. 2009, 87, 2977–2984. [Google Scholar] [CrossRef]
- Santoso, U.; Tanaka, K.; Ohtani, S. Effect of dried Bacillus subtilis culture on growth, body composition and hepatic lipogenic enzyme activity in female broiler chicks. Br. J. Nutr. 1995, 74, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, T.; Amentani, A.; Kiuchi, K.; Kaminogawa, S. Improved growth and viability of lactobacilli in the presence of Bacillus subtilis (natto), catalase, or subtilisin. Can. J. Microbiol. 2000, 46, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Whitley, N.C.; Cazac, D.; Rude, B.J.; Jackson-O’Brien, D.; Parveen, S. Use of a commercial probiotic supplement in meat goats. J. Anim. Sci. 2009, 87, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, M.A.; Joshi, R.S.; Savaliya, F.P.; Patel, A.B.; Mishra, R.K.; Bhagora, N.J. Study on inclusion of probiotic, prebiotic and its combination in broiler diet and their effect on carcass characteristics and economics of commercial broilers. Vet. World 2015, 8, 225–231. [Google Scholar] [CrossRef]
- Stephens, T.P.; Loneragan, G.H.; Karunasena, E.; Brashears, M.M. Reduction of Escherichia coli O157 and Salmonella in feces and on hides of feedlot cattle using various doses of a direct-fed microbial. J. Food Prot. 2007, 70, 2386–2391. [Google Scholar] [CrossRef]
- Feng, J.; Liu, X.; Xu, Z.R.; Lu, Y.P.; Liu, Y.Y. Effect of fermented soybean meal on intestinal morphology and digestive enzyme activities in weaned piglets. Dig. Dis. Sci. 2007, 52, 1845–1850. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Jacobs, D.M.; Deltimple, N.; van Velzen, E.; van Dorsten, F.A.; Bingham, M.; Vaughan, E.E.; van Duynhoven, J. (1)H NMR metabolite profiling of feces as a tool to assess the impact of nutrition on the human microbiome. NMR Biomed. 2008, 21, 615–626. [Google Scholar] [CrossRef]
- Chamberlain, M.; O’Flaherty, S.; Cobián, N.; Barrangou, R. Metabolomic Analysis of Lactobacillus acidophilus, L. gasseri, L. crispatus, and Lacticaseibacillus rhamnosus Strains in the Presence of Pomegranate Extract. Front. Microbiol. 2022, 13, 863228. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, H. The Roles of Glutamine in the Intestine and Its Implication in Intestinal Diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef]
- De Lima, L.B.; da Silva, W.A.V.; Dos Santos, E.C.F.; Machado, J.C.B.; Procópio, T.F.; de Moura, M.C.; Napoleão, T.H.; Ferreira, M.R.A.; Soares, L.A.L. Evaluation of Antioxidant, Antibacterial and Enhancement of Antibiotic Action by Punica granatum Leaves Crude Extract and Enriched Fraction against Multidrug-Resistant Bacteria. Chem. Biodivers. 2021, 18, e2100538. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef]
- Li, B.; Liu, K.; Kwok, L.-Y.; Guo, S.; Bai, L.; Yang, X.; Chen, Y. Development of a non-target metabolomics-based screening method for elucidating metabolic and probiotic potential of bifidobacteria. Innov. Food Sci. Emerg. Technol. 2022, 77, 102971. [Google Scholar] [CrossRef]
- Martias, C.; Baroukh, N.; Mavel, S.; Blasco, H.; Lefèvre, A.; Roch, L.; Montigny, F.; Gatien, J.; Schibler, L.; Dufour-Rainfray, D.; et al. Optimization of Sample Preparation for Metabolomics Exploration of Urine, Feces, Blood and Saliva in Humans Using Combined NMR and UHPLC-HRMS Platforms. Molecules 2021, 26, 4111. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Strain Type | Origin | CBS Number * |
|---|---|---|---|
| BPR-11 | B. amyloliquefaciens | Soil and vegetation | 141692 |
| BPR-12 | B. licheniformis | Soil and vegetation | 141691 |
| BPR-13 | B. subtilis | Soil and vegetation | 141693 |
| BPR-14 | B. amyloliquefaciens | Soil and vegetation | 142360 |
| BPR-16 | B. velezensis | Soil and vegetation | 148295 |
| BPR-17 | B. amyloliquefaciens | Soil and vegetation | 148296 |
| Samples | C. perfringens | E. coli | P. aeruginosa | S. enterica | S. aureus |
|---|---|---|---|---|---|
| C16 surfactin C | 12.5 | 6.25 | 25 | 25 | 6.25 |
| C15 surfactin C | 12.5 | 6.25 | 25 | 25 | 6.25 |
| C14 surfactin C | 12.5 | 6.25 | 25 | 25 | 6.25 |
| C13 surfactin C | 12.5 | 6.25 | 25 | 25 | 12.5 |
| Maculosin | 25 | 25 | NA | NA | 25 |
| Maculosine 2 | 25 | 25 | NA | NA | 25 |
| Genistein | NA | NA | NA | NA | NA |
| Daidzein | NA | NA | NA | NA | NA |
| Probiotic Strains | Specification |
|---|---|
| Bacillus amyloliquefaciens (BPR-11) | 2 × 108 CFU */g |
| Bacillus amyloliquefaciens (BPR-16) | 2 × 108 CFU */g |
| Bacillus amyloliquefaciens (BPR-17) | 2 × 108 CFU */g |
| Treatment Group (n = 128) | p Value | ||
|---|---|---|---|
| Control | Probiotic | ||
| BW0 (g) | 42.38 ± 0.26 | 42.97 ± 0.27 | 0.11 |
| BW7 (g) | 152.30 ± 1.57 | 155.91 ± 1.74 | 0.14 |
| BW14 (g) | 416.54 ± 4.38 | 420.24 ± 4.59 | 0.53 |
| BW21 (g) | 885.46 ± 9.47 | 910.57 ± 9.02 | 0.04 * |
| BW28 (g) | 1573.27 ± 13.43 | 1606.12 ± 15.64 | 0.12 |
| ADG0–7 (g) | 15.70 ± 0.23 | 16.14 ± 0.24 | 0.23 |
| ADG7–14 (g) | 37.75 ± 0.58 | 37.76 ± 0.73 | 0.99 |
| ADG14–21 (g) | 66.99 ± 0.75 | 70.05 ± 0.66 | 0.005 * |
| ADG21–28 (g) | 98.26 ± 1.55 | 99.36 ± 1.15 | 0.57 |
| ADG0–28 (g) | 56.19 ± 1.94 | 57.36 ± 2.14 | 0.12 |
| ADFI0–7 (g) | 16.47 ± 0.29 | 16.52 ± 0.21 | 0.90 |
| ADFI7–14 (g) | 46.50 ± 0.59 | 46.78 ± 0.61 | 0.74 |
| ADFI14–21 (g) | 83.49 ± 0.70 | 87.58 ± 0.80 | 0.00061 * |
| ADFI21–28 (g) | 138.12 ± 1.67 | 138.69 ± 1.57 | 0.80 |
| ADFI0–28 (g) | 71.95 ± 2.00 | 72.47 ± 2.68 | 0.63 |
| FCR0–7 (g/g) | 0.76 ± 0.01 | 0.74 ± 0.00 | 0.17 |
| FCR7–14 (g/g) | 1.23 ± 0.01 | 1.24 ± 0.01 | 0.58 |
| FCR14–21 (g/g) | 1.25 ± 0.01 | 1.25 ± 0.00 | 0.70 |
| FCR21–28 (g/g) | 1.41 ± 0.01 | 1.40 ± 0.01 | 0.29 |
| FCR0–28 (g/g) | 1.28 ± 0.00 | 1.26 ± 0.00 | 0.01 * |
| Mortality (%) | 3.13 | 1.56 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, C.; Horyanto, D.; Stanley, D.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters. Antibiotics 2023, 12, 407. https://doi.org/10.3390/antibiotics12020407
Tran C, Horyanto D, Stanley D, Cock IE, Chen X, Feng Y. Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters. Antibiotics. 2023; 12(2):407. https://doi.org/10.3390/antibiotics12020407
Chicago/Turabian StyleTran, Charlie, Darwin Horyanto, Dragana Stanley, Ian E. Cock, Xiaojing Chen, and Yunjiang Feng. 2023. "Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters" Antibiotics 12, no. 2: 407. https://doi.org/10.3390/antibiotics12020407
APA StyleTran, C., Horyanto, D., Stanley, D., Cock, I. E., Chen, X., & Feng, Y. (2023). Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters. Antibiotics, 12(2), 407. https://doi.org/10.3390/antibiotics12020407