Assessing the Potential of Gallic Acid and Methyl Gallate to Enhance the Efficacy of β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus by Targeting β-Lactamase: In Silico and In Vitro Studies


Abstract
:1. Introduction
2. Results
2.1. Antibacterial Susceptibility
2.2. Enhancing Effects of GA and MG on Anti-MRSA Activity of β-Lactam Antibiotics
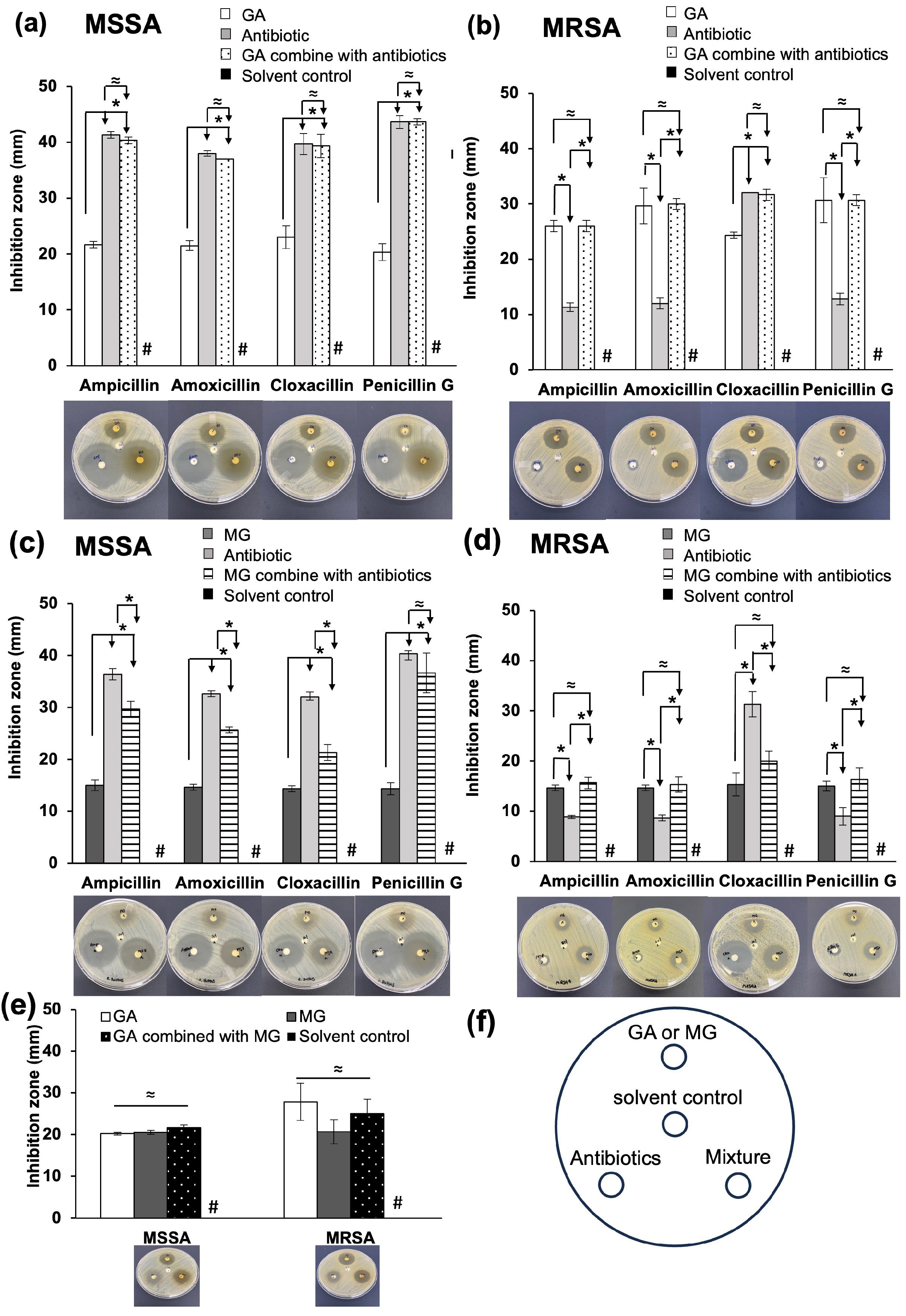
2.2.1. Disc Diffusion Assay
2.2.2. Synergistic Effects of GA and MG on the Anti-MRSA Activity of β-Lactam Antibiotics
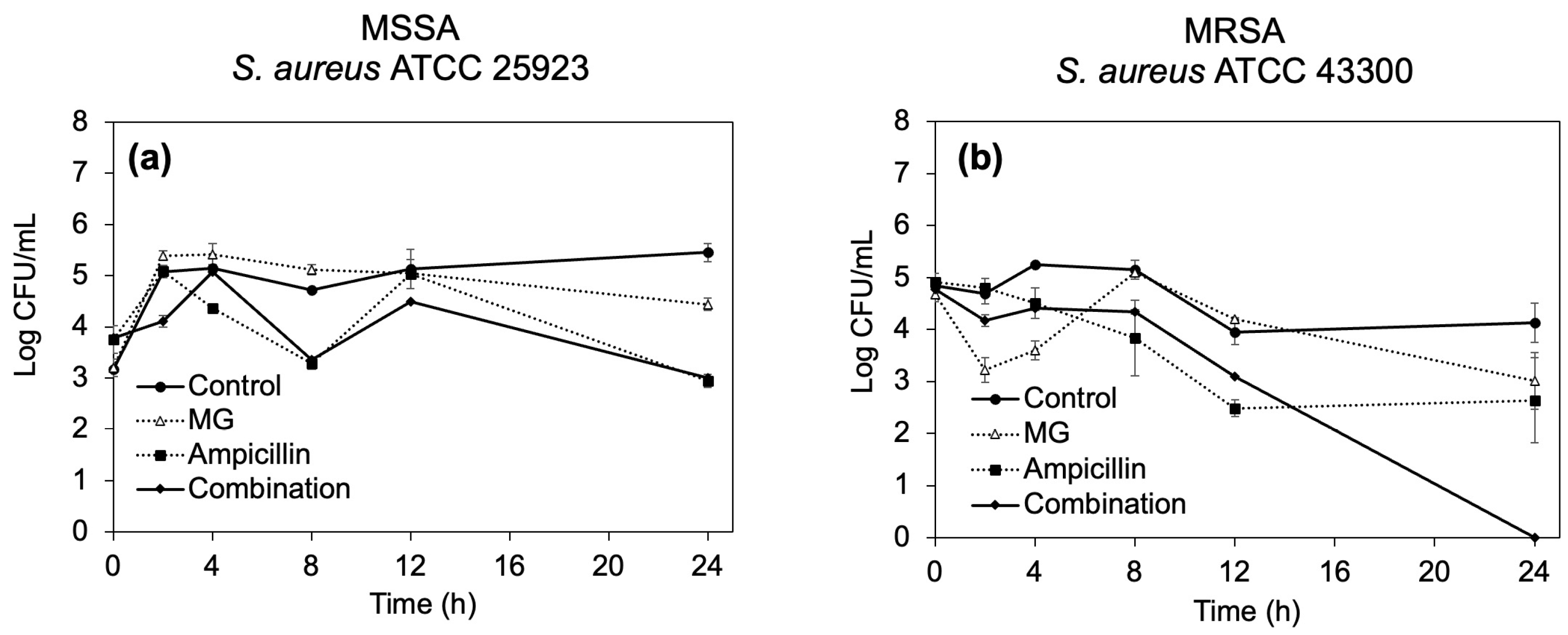
2.2.3. Time–Kill Curves
2.3. β-Lactamase Inhibition
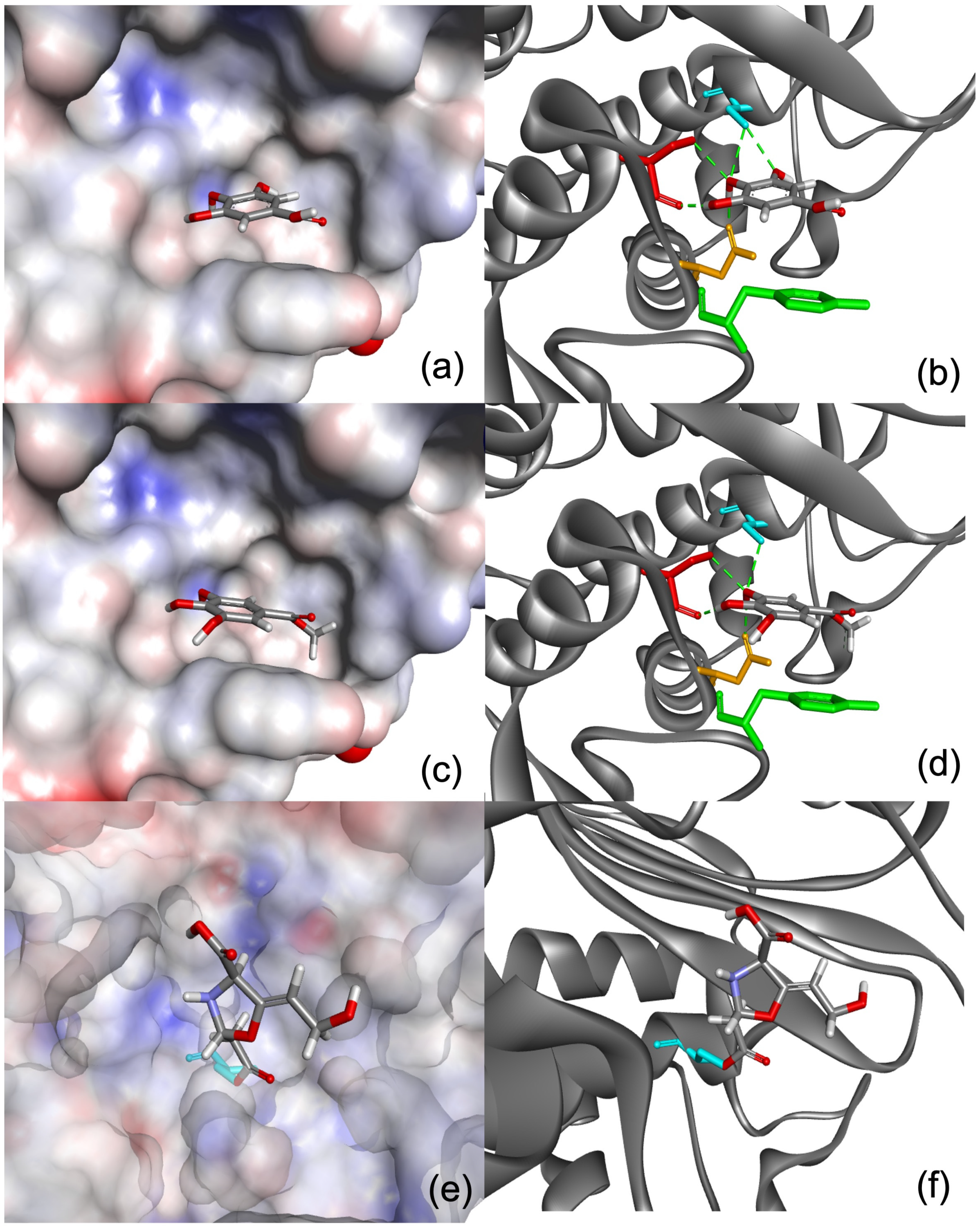
2.3.1. In Silico Molecular Modelling
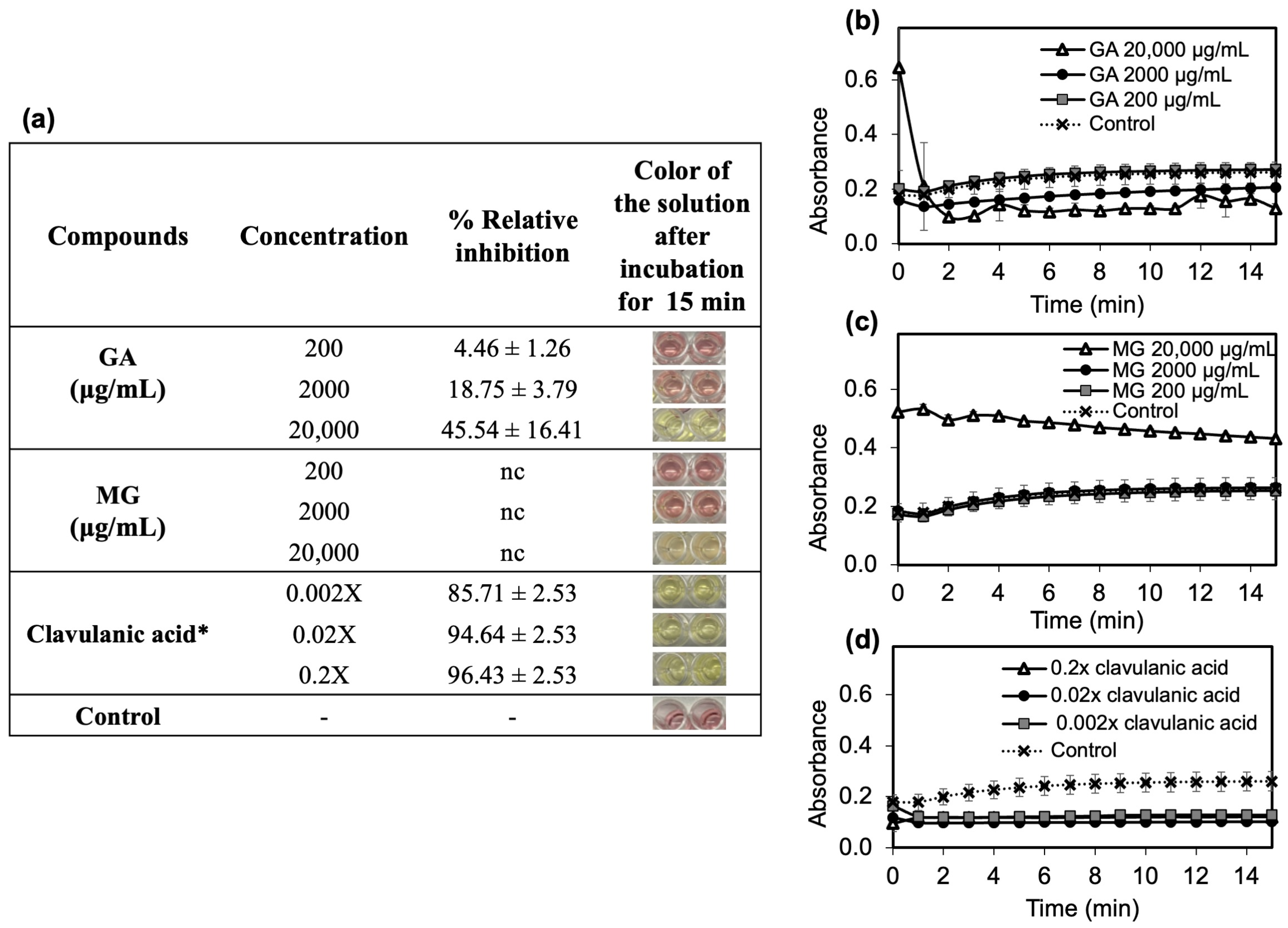
2.3.2. In Vitro β-Lactamase Inhibition
2.4. SEM
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bacterial Strains and Culture Conditions
4.3. Antibacterial Susceptibility Test
4.4. Enhancing Effects of GA and MG on Anti-MRSA Activity of β-Lactam Antibiotics
4.4.1. Disc Diffusion Assay
4.4.2. Checkerboard Assay
4.4.3. Time–Kill Assay
4.5. β-Lactamase Inhibition Assays
4.5.1. Molecular Docking Study
4.5.2. In Vitro β-Lactamase Inhibition Assay
4.6. SEM
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, K.; Bradford, P.A. β-lactams and β-lactamase Inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur. J. Med. Chem. 2020, 208, 112829. [Google Scholar] [CrossRef] [PubMed]
- Lobanovska, M.; Pilla, G. Penicillin’s discovery and antibiotic resistance: Lessons for the future? Yale J. Biol. Med. 2017, 90, 135–145. [Google Scholar] [PubMed]
- Novy, E.; Martinière, H.; Roger, C. The current status and future perspectives of beta-lactam therapeutic drug monitoring in critically Ill patients. Antibiotics 2023, 12, 681. [Google Scholar] [CrossRef]
- Stapleton, P.D.; Taylor, P.W. Methicillin resistance in Staphylococcus aureus: Mechanisms and modulation. Sci. Prog. 2002, 85, 57–72. [Google Scholar] [CrossRef]
- Essack, S.Y. The development of β-lactam antibiotics in response to the evolution of β-lactamases. Pharm. Res. 2001, 18, 1391–1399. [Google Scholar] [CrossRef]
- Marin, M. Methicillin resistant Staphylococcus. Medicina 2002, 62, 30–35. [Google Scholar]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Davis, J.S.; van Hal, S.; Tong, S. Combination antibiotic treatment of serious methicillin-resistant Staphylococcus aureus infections. Semin. Respir. Crit. Care Med. 2015, 36, 3–16. [Google Scholar]
- González-Bello, C.; Rodríguez, D.; Pernas, M.; Rodríguez, Á.; Colchón, E. β-Lactamase inhibitors to restore the efficacy of antibiotics against superbugs. J. Med. Chem. 2020, 63, 1859–1881. [Google Scholar] [CrossRef]
- Liang, H.; Huang, Q.; Zou, L.; Wei, P.; Lu, J.; Zhang, Y. Methyl gallate: Review of pharmacological activity. Pharmacol. Res. 2023, 194, 106849. [Google Scholar] [CrossRef] [PubMed]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and diseases: A mechanistic review. Iran. J. Basic Med. Sci. 2019, 22, 225–237. [Google Scholar] [PubMed]
- Rasool, M.K.; Sabina, E.P.; Ramya, S.R.; Preety, P.; Patel, S.; Mandal, N.; Mishra, P.P.; Samuel, J. Hepatoprotective and antioxidant effects of gallic acid in paracetamol-induced liver damage in mice. J. Pharm. Pharmacol. 2010, 62, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Pei, J.; Zheng, Y.; Miao, Y.; Duan, B.; Huang, L. Gallic acid: A potential anti-cancer agent. Chin. J. Integr. Med. 2022, 28, 661–671. [Google Scholar] [CrossRef]
- Huang, C.; Chang, Y.; Wei, P.; Hung, C.; Wang, W. Methyl gallate, gallic acid-derived compound, inhibit cell proliferation through increasing ROS production and apoptosis in hepatocellular carcinoma cells. PLoS ONE 2021, 16, e0248521. [Google Scholar] [CrossRef]
- Sharanya, C.S.; Arun, K.G.; Vijaytha, V.; Sabu, A.; Haeidas, M. Designing of enzyme inhibitors based on active site specificity: Lessons from methyl gallate and its lipoxygenase inhibitory profile. J. Recept. Signal Transduct. 2018, 38, 256–265. [Google Scholar]
- Toyama, M.H.; Rogero, A.; de Moraes, L.L.F.; Fernandes, G.A.; da Cruz Costa, C.R.; Belchor, M.N.; De Carli, A.M.; de Oliveira, M.A. Gallic acid as a non-selective inhibitor of α/β-hydrolase fold enzymes involved in the inflammatory process: The two sides of the same coin. Pharmaceutics 2022, 14, 368. [Google Scholar] [CrossRef]
- Shao, D.; Li, J.; Li, J.; Tang, R.; Liu, L.; Shi, J.; Huang, Q.; Yang, H. Inhibition of gallic acid on the growth and biofilm formation of Escherichia coli and Streptococcus mutans. J. Food Sci. 2015, 80, M1299–M1305. [Google Scholar] [CrossRef]
- Kang, M.; Oh, J.; Kang, I.; Hong, S.; Choi, C. Inhibitory effect of methyl gallate and gallic acid on oral bacteria. J. Microbiol. 2008, 46, 744–750. [Google Scholar] [CrossRef]
- Rivero-Buceta, E.; Carrero, P.; Doyagueez, E.G.; Madrona, A.; Quesada, E.; Camarasa, M.J.; Peréz-Pérez, M.J.; Leyssen, P.; Paeshuyse, J.; Balzarini, J. Linear and branched alkyl-esters and amides of gallic acid and other (mono-, di-and tri-) hydroxy benzoyl derivatives as promising anti-HCV inhibitors. Eur. J. Med. Chem. 2015, 92, 656–671. [Google Scholar] [CrossRef]
- Qin, W.; Wang, Y.; Mouhamed, F.; Hamaker, B.; Zhang, G. Impact of the solubility of phenolic compounds from highland barley (Hordeum vulgare L.) on their antioxidant property and protein binding affinity. LWT 2023, 186, 115251. [Google Scholar] [CrossRef]
- Lobiuc, A.; Pavăl, N.-E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.-C.; Amăriucăi-Mantu, D.; Stoleru, V. Future antimicrobials: Natural and functionalized phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Wang, L.; Niu, X.; Cai, H.; Zhong, X.; Deng, X.; Wang, T.; Wang, D. The use of chlorogenic acid and its analogues as inhibitors: An investigation of the inhibition of sortase A of Staphylococcus aureus using molecular docking and dynamic simulation. Biotechnol. Lett. 2016, 38, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-H.; Hu, Z.-Q.; Hara, Y.; Shimamura, T. Inhibition of penicillinase by epigallocatechin gallate resulting in restoration of antibacterial activity of penicillin against penicillinase-producing Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2266–2268. [Google Scholar] [CrossRef]
- Jiamboonsri, P.; Pithayanukul, P.; Bavovada, R.; Chomnawang, M.T. The inhibitory potential of Thai mango seed kernel extract against methicillin-resistant Staphylococcus aureus. Molecules 2011, 16, 6255–6270. [Google Scholar] [CrossRef]
- Gobin, M.; Proust, R.; Lack, S.; Duciel, L.; Des Courtils, C.; Pauthe, E.; Gand, A.; Seyer, D. A Combination of the natural molecules gallic acid and carvacrol eradicates P. aeruginosa and S. aureus mature biofilms. Int. J. Mol. Sci. 2022, 23, 7118. [Google Scholar] [CrossRef]
- Kubo, I.; Xiao, P.; Fujita, K.I. Anti-MRSA activity of alkyl gallates. Bioorg. Med. Chem. Lett. 2002, 12, 113–116. [Google Scholar] [CrossRef]
- Dehbashi, S.; Tahmasebi, H.; Zeyni, B.; Arabestani, M.R. Regulation of virulence and β-lactamase gene expression in Staphylococcus aureus isolates: Cooperation of two-component systems in bloodstream superbugs. BMC Microbiol. 2021, 21, 192. [Google Scholar] [CrossRef]
- Peacock, S.J.; Paterson, G.K. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef]
- Dos Santos, J.F.S.; Tintino, S.R.; De Freitas, T.S.; Campina, F.F.; Menezes, I.R.D.A.; Siqueira-Júnior, J.P.; Coutinho, H.D.M.; Cunha, F.A.B. In vitro e in silico evaluation of the inhibition of Staphylococcus aureus efflux pumps by caffeic and gallic acid. Comp. Immunol. Microbiol. Infect. Dis. 2018, 57, 22–28. [Google Scholar] [CrossRef]
- Montanari, M.P.; Piccoli, L.; Mingoia, M.; Marchetti, F.; Varaldo, P.E. Synergistic potential of ceftazidime plus amikacin or levofloxacin against Pseudomonas aeruginosa as determined using a checkerboard and a disk diffusion technique. Diagn. Microbiol. Infect. Dis. 2005, 53, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Gaudereto, J.J.; Neto, L.V.P.; Leite, G.C.; Espinoza, E.P.S.; Martins, R.C.R.; Villas Boa Prado, G.; Rossi, F.; Guimarães, T.; Levin, A.S.; Costa, S.F. Comparison of methods for the detection of in vitro synergy in multidrug-resistant gram-negative bacteria. BMC Microbiol. 2020, 20, 97. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Kondo, K.; Katsuyama, R.; Kawazoe, K.; Sato, Y.; Murakami, K.; Takaishi, Y.; Arakaki, N.; Higuti, T. Alkyl gallates, intensifiers of β-lactam susceptibility in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Bae, J.; Park, M.; Jeon, B. Potentiation of beta-lactams against methicillin-resistant Staphylococcus aureus (MRSA) using octyl gallate, a food-grade antioxidant. Antibiotics 2022, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Okoliegbe, I.N.; Hijazi, K.; Cooper, K.; Ironside, C.; Gould, I.M. Antimicrobial synergy testing: Comparing the tobramycin and ceftazidime gradient diffusion methodology used in assessing synergy in cystic fibrosis-derived multidrug-resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 967. [Google Scholar] [CrossRef]
- March, G.A.; Bratos, M.A. A meta-analysis of in vitro antibiotic synergy against Acinetobacter baumannii. J. Microbiol. Methods 2015, 119, 31–36. [Google Scholar] [CrossRef]
- Thieme, L.; Klinger-Strobel, M.; Hartung, A.; Stein, C.; Makarewicz, O.; Pletz, M.W. In vitro synergism and anti-biofilm activity of ampicillin, gentamicin, ceftaroline and ceftriaxone against Enterococcus faecalis. J. Antimicrob. Chemother. 2018, 73, 1553–1561. [Google Scholar] [CrossRef]
- Bush, K. CHAPTER 14—β-Lactam antibiotics: Penicillins. In Antibiotic and Chemotherapy, 9th ed.; Finch, R.G., Greenwood, D., Norrby, S.R., Whitley, R.J., Eds.; W.B. Saunders: London, UK, 2010; pp. 200–225. [Google Scholar]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding phenolic acids inhibition of α-amylase and α-glucosidase and influence of reaction conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef]
- Nithitanakool, S.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular docking studies and anti-tyrosinase activity of Thai mango seed kernel extract. Molecules 2009, 14, 257–265. [Google Scholar] [CrossRef]
- Vankar, P.S.; Srivastava, J. Evaluation of anthocyanin content in red and blue flowers. Int. J. Food Eng. 2010, 6, 4. [Google Scholar] [CrossRef]
- Song, H.; Chen, C.; Zhao, S.; Ge, F.; Liu, D.; Shi, D.; Zhang, T. Interaction of gallic acid with trypsin analyzed by spectroscopy. J. Food Drug. Anal. 2015, 23, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Wei, S.; Su, H.; Zheng, S.; Xu, S.; Liu, M.; Bo, R.; Li, J. Bactericidal activity of gallic acid against multi-drug resistance Escherichia coli. Microb. Pathog. 2022, 173, 105824. [Google Scholar] [CrossRef] [PubMed]
- Mechesso, A.F.; Yixian, Q.; Park, S.-C. Methyl gallate and tylosin synergistically reduce the membrane integrity and intracellular survival of Salmonella Typhimurium. PLoS ONE 2019, 14, e0221386. [Google Scholar] [CrossRef] [PubMed]
- Jiamboonsri, P.; Kanchanadumkerng, P. Influence of gallic acid and Thai culinary essential oils on antibacterial activity of nisin against Streptococcus mutans. Adv. Pharmacol. Pharm. Sci. 2021, 2021, 5539459. [Google Scholar] [CrossRef]
- Faleiro, M.L.; Miguel, M.G. Chapter 6—Use of essential oils and their components against multidrug-resistant bacteria. In Fighting Multidrug Resistance with Herbal Extracts, Essential Oils and Their Components; Rai, M.K., Kon, K.V., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 65–94. [Google Scholar]
- Petersen, P.J.; Labthavikul, P.; Jones, C.H.; Bradford, P.A. In vitro antibacterial activities of tigecycline in combination with other antimicrobial agents determined by chequerboard and time-kill kinetic analysis. J. Antimicrob. Chemother. 2006, 57, 573–576. [Google Scholar] [CrossRef]
- Allinger, N.L. Conformational analysis. 130. MM2. A hydrocarbon force field utilizing V1 and V2 torsional terms. J. Am. Chem. Soc. 1977, 99, 8127–8134. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Chen, C.C.; Herzberg, O. Inhibition of β-lactamase by clavulanate. Trapped intermediates in cryocrystallographic studies. J. Mol. Biol. 1992, 224, 1103–1113. [Google Scholar] [CrossRef]
- Korb, O.; Stützle, T.; Exner, T.E. Empirical scoring functions for advanced protein−ligand docking with PLANTS. J. Chem. Inf. Model. 2009, 49, 84–96. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Compounds | MSSA S. aureus ATCC 25923 | MRSA S. aureus ATCC 43300 | ||
|---|---|---|---|---|
| MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | |
| GA | 200 | 400 | 400 | >400 |
| MG | 1600 | 3200 | 1600 | 3200 |
| Ampicillin | 0.125 | 0.25 | 62.5 | 250.0 |
| Amoxicillin | 0.25 | 0.5 | 62.5 | 250.0 |
| Cloxacillin | 0.25 | 0.5 | 0.5 | 1.0 |
| Penicillin G | 0.0625 | 0.125 | 62.5 | 125.0 |
| Solvent a | - | - | - | - |
| Bacterial Strains | Compounds | MIC | Combinded Compounds (μg/mL) | MIC | The Lowest FIC Indexes | |||
|---|---|---|---|---|---|---|---|---|
| Alone (μg/mL) | Combined (μg/mL) | Alone (μg/mL) | Combinded (μg/mL) | |||||
| MSSA (S. aureus ATCC 25923) | GA | 200 | 6.25 | Ampicillin | 0.125 | 0.125 | 1.03 | IND * |
| 6.25 | Amoxicillin | 0.25 | 0.25 | 1.03 | IND * | |||
| 6.25 | Cloxacillin | 0.25 | 0.25 | 1.03 | IND * | |||
| 6.25 | Penicillin G | 0.063 | 0.063 | 1.03 | IND * | |||
| MG | 1600 | 50 | Ampicillin | 0.125 | 0.125 | 1.03 | IND * | |
| 50 | Amoxicillin | 0.25 | 0.25 | 1.03 | IND * | |||
| 50 | Cloxacillin | 0.25 | 0.25 | 1.03 | IND * | |||
| 50 | Penicillin G | 0.063 | 0.063 | 1.03 | IND * | |||
| MG | 1600 | 800 | GA | 200 | 25 | 0.63 | ADD * | |
| MRSA (S. aureus ATCC 43300) | GA | 400 | 100 | Ampicillin | 62.5 | 15.625 | 0.5 | ADD * |
| 100 | Amoxicillin | 62.5 | 15.625 | 0.5 | ADD * | |||
| 200 | Cloxacillin | 0.5 | 0.25 | 1 | IND * | |||
| 200 | Penicillin G | 62.5 | 7.813 | 0.63 | ADD * | |||
| MG | 1600 | 100 | Ampicillin | 62.5 | 15.625 | 0.313 | SYN * | |
| 200 | Amoxicillin | 62.5 | 15.625 | 0.375 | SYN * | |||
| 100 | Cloxacillin | 0.5 | 0.5 | 1.06 | IND * | |||
| 200 | Penicillin G | 62.5 | 15.625 | 0.375 | SYN * | |||
| MG | 1600 | 800 | GA | 400 | 50 | 0.63 | ADD * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiamboonsri, P.; Eurtivong, C.; Wanwong, S. Assessing the Potential of Gallic Acid and Methyl Gallate to Enhance the Efficacy of β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus by Targeting β-Lactamase: In Silico and In Vitro Studies. Antibiotics 2023, 12, 1622. https://doi.org/10.3390/antibiotics12111622
Jiamboonsri P, Eurtivong C, Wanwong S. Assessing the Potential of Gallic Acid and Methyl Gallate to Enhance the Efficacy of β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus by Targeting β-Lactamase: In Silico and In Vitro Studies. Antibiotics. 2023; 12(11):1622. https://doi.org/10.3390/antibiotics12111622
Chicago/Turabian StyleJiamboonsri, Pimsumon, Chatchakorn Eurtivong, and Sompit Wanwong. 2023. "Assessing the Potential of Gallic Acid and Methyl Gallate to Enhance the Efficacy of β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus by Targeting β-Lactamase: In Silico and In Vitro Studies" Antibiotics 12, no. 11: 1622. https://doi.org/10.3390/antibiotics12111622
APA StyleJiamboonsri, P., Eurtivong, C., & Wanwong, S. (2023). Assessing the Potential of Gallic Acid and Methyl Gallate to Enhance the Efficacy of β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus by Targeting β-Lactamase: In Silico and In Vitro Studies. Antibiotics, 12(11), 1622. https://doi.org/10.3390/antibiotics12111622