
Spread of Pseudomonas aeruginosa ST274 Clone in Different Niches: Resistome, Virulome, and Phylogenetic Relationship

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
2.1. Resistance Phenotype
2.2. Serotyping and Pulsed-Field Gel Electrophoresis (PFGE)
2.3. Detection of Virulence and Quorum-Sensing Genes
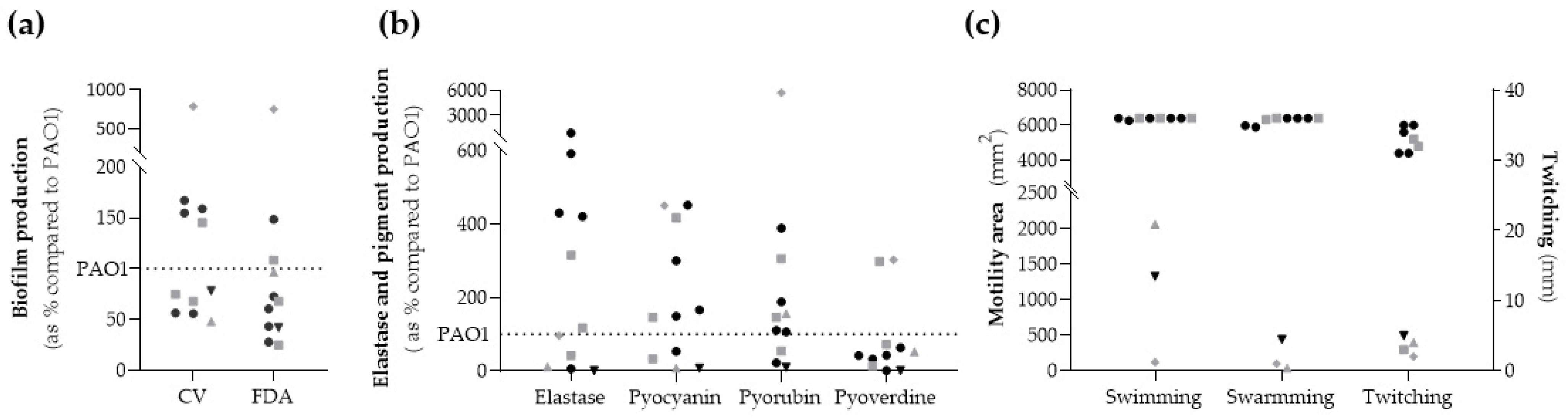
2.4. Biofilm Formation
2.5. Elastase Activity and Pigment Production
2.6. Motility
2.7. Genome Properties of ST274-P. aeruginosa Strains from Different Origins
2.7.1. Phylogenetic Analysis
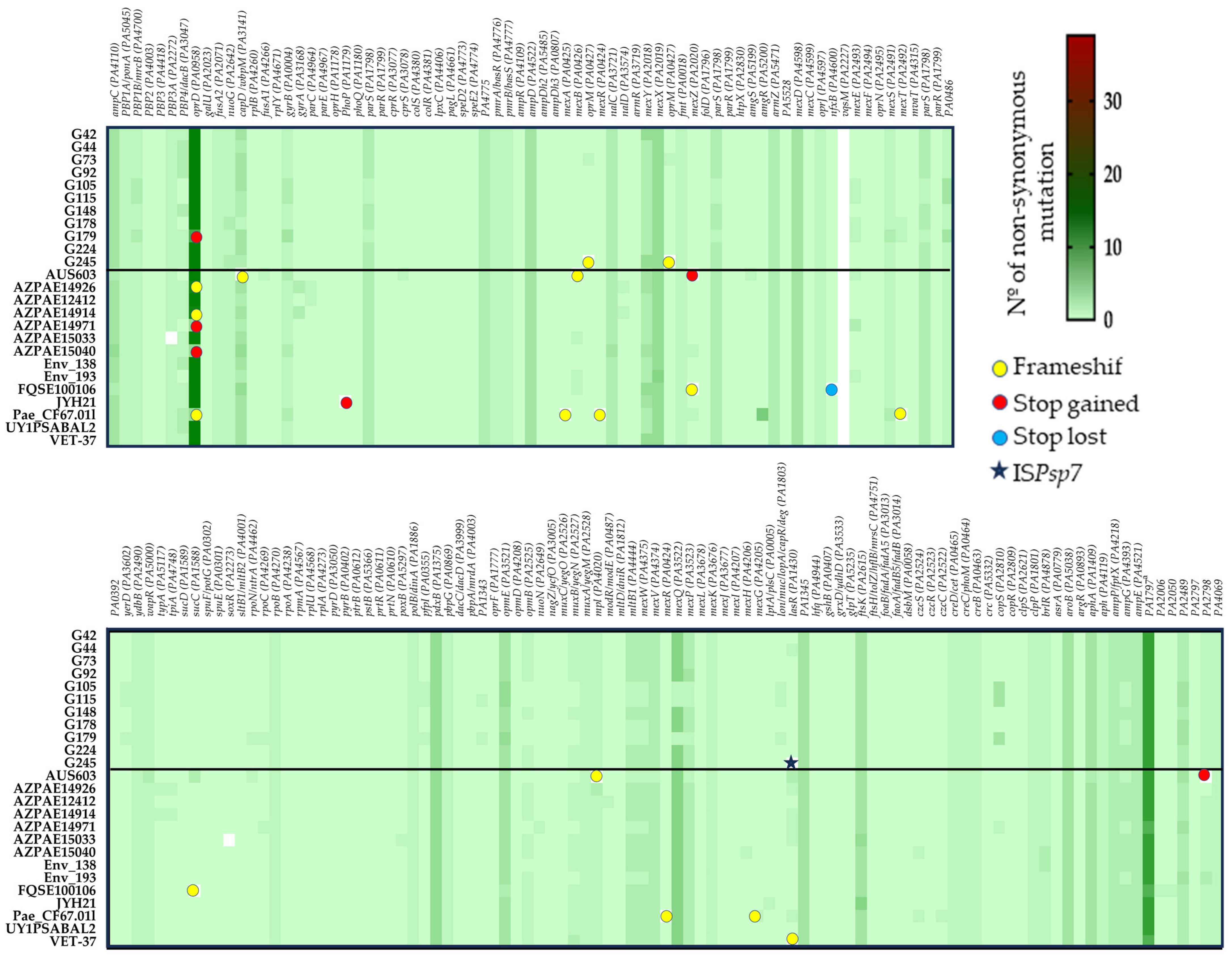
2.7.2. Resistome
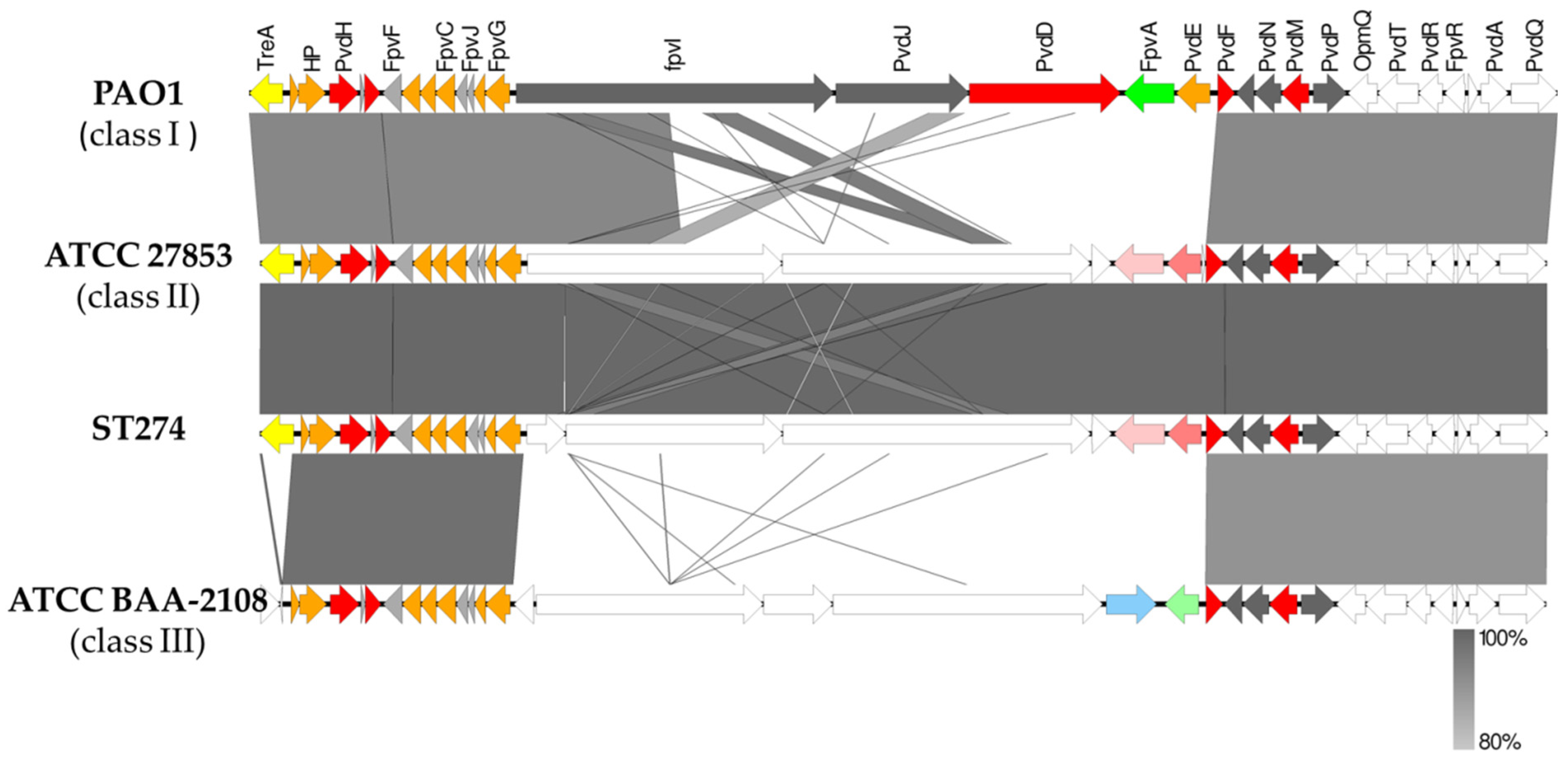
2.7.3. Detection of Virulence and Quorum-Sensing Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antimicrobial Susceptibility Testing
4.3. Serotyping
4.4. Detection of Virulence and Quorum-Sensing Genes
4.5. Clonal Relationship
4.6. Biofilm Formation
4.7. Elastase and Pigment Production
4.8. Motility
4.9. Whole Genome Sequencing (WGS)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Freschi, L.; Vincent, A.T.; Jeukens, J.; Emond-Rheault, J.G.; Kukavica-Ibrulj, I.; Dupont, M.J.; Charette, S.J.; Boyle, B.; Levesque, R.C. The Pseudomonas aeruginosa Pan-Genome Provides New Insights on Its Population Structure, Horizontal Gene Transfer, and Pathogenicity. Genome Biol. Evol. 2018, 11, 109–120. [Google Scholar] [CrossRef] [PubMed]
- López-Causapé, C.; de Dios-Caballero, J.; Cobo, M.; Escribano, A.; Asensio, Ó.; Oliver, A.; del Campo, R.; Cantón, R.; Solé, A.; Cortell, I.; et al. Antibiotic resistance and population structure of cystic fibrosis Pseudomonas aeruginosa isolates from a Spanish multi-centre study. Int. J. Antimicrob. Agents 2017, 50, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic pathogen and lab rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Subedi, D.; Vijay, A.K.; Kohli, G.S.; Rice, S.A.; Willcox, M. Comparative genomics of clinical strains of Pseudomonas aeruginosa strains isolated from different geographic sites. Sci. Rep. 2018, 8, 15668. [Google Scholar] [CrossRef]
- Malhotra, S.; Hayes, D., Jr.; Wozniak, D.J. Cystic Fibrosis and Pseudomonas aeruginosa: The Host-Microbe Interface. Clin. Microbiol. Rev. 2019, 32, e00138-18. [Google Scholar] [CrossRef]
- Jurado-Martín, I.; Sainz-Mejías, M.; McClean, S. Pseudomonas aeruginosa: An Audacious Pathogen with an Adaptable Arsenal of Virulence Factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef]
- Lozano, C.; Azcona-Gutiérrez, J.M.; Van Bambeke, F.; Sáenz, Y. Great phenotypic and genetic variation among successive chronic Pseudomonas aeruginosa from a cystic fibrosis patient. PLoS ONE 2018, 13, e0204167. [Google Scholar] [CrossRef]
- Mauch, R.M.; Jensen, P.Ø.; Moser, C.; Levy, C.E.; Høiby, N. Mechanisms of humoral immune response against Pseudomonas aeruginosa biofilm infection in cystic fibrosis. J. Cyst. Fibros. 2018, 17, 143–152. [Google Scholar] [CrossRef]
- Gabrielaite, M.; Johansen, H.K.; Molin, S.; Nielsen, F.C.; Marvig, R.L. Gene Loss and Acquisition in Lineages of Pseudomonas aeruginosa Evolving in Cystic Fibrosis Patient Airways. mBio 2020, 11, e02359-20. [Google Scholar] [CrossRef]
- Nageeb, W.; Amin, D.H.; Mohammedsaleh, Z.M.; Makharita, R.R. Novel Molecular Markers Linked to Pseudomonas aeruginosa Epidemic High-Risk Clones. Antibiotics 2021, 10, 35. [Google Scholar] [CrossRef]
- López-Causapé, C.; Rojo-Molinero, E.; Mulet, X.; Cabot, G.; Moyà, B.; Figuerola, J.; Togores, B.; Pérez, J.L.; Oliver, A. Clonal Dissemination, Emergence of Mutator Lineages and Antibiotic Resistance Evolution in Pseudomonas aeruginosa Cystic Fibrosis Chronic Lung Infection. PLoS ONE 2013, 8, e71001. [Google Scholar] [CrossRef] [PubMed]
- Ocampo-Sosa, A.A.; Fernández-Martínez, M.; Cabot, G.; Peña, C.; Tubau, F.; Oliver, A.; Martínez-Martínez, L. Draft Genome Sequence of the Quorum-Sensing and Biofilm-Producing Pseudomonas aeruginosa Strain Pae221, Belonging to the Epidemic High-Risk Clone Sequence Type 274. Genome Announc. 2015, 3, e01343-14. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Olmos, A.; García-Castillo, M.; Alba, J.M.; Morosini, M.I.; Lamas, A.; Romero, B.; Galán, J.C.; del Campo, R.; Cantón, R. Population structure and antimicrobial susceptibility of both nonpersistent and persistent Pseudomonas aeruginosa isolates recovered from cystic fibrosis patients. J. Clin. Microbiol. 2013, 51, 2761–2765. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Lara, S.; del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A.; Martínez-Martínez, L.; Bou, G.; Zamorano, L.; Sánchez-Diener, I.; Galán, F.; Gracia, I.; et al. Predicting Pseudomonas aeruginosa susceptibility phenotypes from whole genome sequence resistome analysis. Clin. Microbiol. Infect. 2021, 27, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The resistome of Pseudomonas aeruginosa in relationship to phenotypic susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef]
- Bocharova, Y.A.; Savinova, T.A.; Lyamin, A.V.; Kondratenko, O.V.; Polikarpova, S.V.; Zhilina, S.V.; Fedorova, N.I.; Semykin, S.Y.; Chaplin, A.V.; Korostin, D.O.; et al. Genome features and antibiotic resistance of Pseudomonas aeruginosa strains isolated in patients with cystic fibrosis in the Russian Federation. Russ. Clin. Lab. Diagn. 2021, 66, 629–634. [Google Scholar] [CrossRef]
- Ahmed, M.A.S.; Hadi, H.A.; Abu Jarir, S.; Khan, F.A.; Arbab, M.A.; Hamid, J.M.; Alyazidi, M.A.; Al-Maslamani, M.A.; Skariah, S.; Sultan, A.A.; et al. Prevalence and microbiological and genetic characteristics of multidrug-resistant Pseudomonas aeruginosa over three years in Qatar. Antimicrob. Steward. Healthc. Epidemiol. 2022, 2, e96. [Google Scholar] [CrossRef]
- Van Mansfeld, R.; Willems, R.; Brimicombe, R.; Heijerman, H.; Van Berkhout, F.T.; Wolfs, T.; Van der Ent, C.; Bonten, M. Pseudomonas aeruginosa genotype prevalence in Dutch cystic fibrosis patients and age dependency of colonization by various P. aeruginosa sequence types. J. Clin. Microbiol. 2009, 47, 4096–4101. [Google Scholar] [CrossRef]
- Mitchelmore, P.J.; Randall, J.; Bull, M.J.; Moore, K.A.; O’Neill, P.A.; Paszkiewicz, K.; Mahenthiralingam, E.; Scotton, C.J.; Sheldon, C.D.; Withers, N.J.; et al. Molecular epidemiology of Pseudomonas aeruginosa in an unsegregated bronchiectasis cohort sharing hospital facilities with a cystic fibrosis cohort. Thorax 2017, 73, 677–679. [Google Scholar] [CrossRef]
- Ruiz-Roldán, L.; Rojo-Bezares, B.; de Toro, M.; López, M.; Toledano, P.; Lozano, C.; Chichón, G.; Alvarez-Erviti, L.; Torres, C.; Sáenz, Y. Antimicrobial resistance and virulence of Pseudomonas spp. among healthy animals: Concern about exolysin ExlA detection. Sci. Rep. 2020, 10, 11667. [Google Scholar] [CrossRef]
- Torres, R.T.; Cunha, M.V.; Ferreira, H.; Fonseca, C.; Palmeira, J.D. A high-risk carbapenem-resistant Pseudomonas aeruginosa clone detected in red deer (Cervus elaphus) from Portugal. Sci. Total Environ. 2022, 829, 154699. [Google Scholar] [CrossRef] [PubMed]
- CLSI 2022; Performance Standards for Antimicrobial Susceptibility Testing. 32th Informational Supplement; CLSI Document M100-S32; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022.
- Kus, J.V.; Tullis, E.; Cvitkovitch, D.G.; Burrows, L.L. Significant differences in type IV pilin allele distribution among Pseudomonas aeruginosa isolates from cystic fibrosis (CF) versus non-CF patients. Microbiology 2004, 150 Pt 5, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Caballero, J.D.; Wheatley, R.M.; Kapel, N.; López-Causapé, C.; Van der Schalk, T.; Quinn, A.; Shaw, L.P.; Ogunlana, L.; Recanatini, C.; Xavier, B.B.; et al. Mixed strain pathogen populations accelerate the evolution of antibiotic resistance in patients. Nat. Commun. 2023, 14, 4083. [Google Scholar] [CrossRef] [PubMed]
- López-Causapé, C.; Cabot, G.; Del Barrio-Tofiño, E.; Oliver, A. The Versatile Mutational Resistome of Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 685. [Google Scholar] [CrossRef]
- Rojo-Bezares, B.; Estepa, V.; Cebollada, R.; de Toro, M.; Somalo, S.; Seral, C.; Castillo, F.J.; Torres, C.; Sáenz, Y. Carbapenem-resistant Pseudomonas aeruginosa strains from a Spanish hospital: Characterization of metallo-beta-lactamases, porin OprD and integrons. Int. J. Med. Microbiol. 2014, 304, 405–414. [Google Scholar] [CrossRef]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M.G.J.; Orange, N.; Dufour, A.; Cornelis, P. Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef]
- Lloyd, M.G.; Vossler, J.L.; Nomura, C.T.; Moffat, J.F. Blocking RpoN reduces virulence of Pseudomonas aeruginosa isolated from cystic fibrosis patients and increases antibiotic sensitivity in a laboratory strain. Sci. Rep. 2019, 9, 6677. [Google Scholar] [CrossRef]
- Viducic, D.; Ono, T.; Murakami, K.; Katakami, M.; Susilowati, H.; Miyake, Y. rpoN gene of Pseudomonas aeruginosa alters its susceptibility to quinolones and carbapenems. Antimicrob. Agents Chemother. 2007, 51, 1455–1462. [Google Scholar] [CrossRef]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef]
- Kadri, S.S.; Adjemian, J.; Lai, Y.L.; Spaulding, A.B.; Ricotta, E.; Prevots, D.R.; Palmore, T.N.; Rhee, C.; Klompas, M.; Dekker, J.P.; et al. Difficult-to-Treat Resistance in Gram-Negative Bacteremia at 173 US Hospitals: Retrospective Cohort Analysis of Prevalence, Predictors, and Outcome of Resistance to All First-Line Agents. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 1803–1814. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Guidance on the Treatment of Extended-Spectrum β-lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin. Infect. Dis. 2022, 75, 187–212. [Google Scholar] [CrossRef]
- Camus, L.; Vandenesch, F.; Moreau, K. From genotype to phenotype: Adaptations of Pseudomonas aeruginosa to the cystic fibrosis environment. Microb. Genom. 2021, 7, 000513. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, D.A.; Wu, M.; Hoffman, L.R.; Kulasekara, H.D.; Déziel, E.; Smith, E.E.; Nguyen, H.; Ernst, R.K.; Larson Freeman, T.J.; Spencer, D.H.; et al. Growth phenotypes of Pseudomonas aeruginosa lasR mutants adapted to the airways of cystic fibrosis patients. Mol. Microbiol. 2007, 64, 512–533. [Google Scholar] [CrossRef] [PubMed]
- Groleau, M.C.; de Oliveira Pereira, T.; Dekimpe, V.; Déziel, E. PqsE Is Essential for RhlR-Dependent Quorum Sensing Regulation in Pseudomonas aeruginosa. mSystems 2020, 5, e00194-20. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Kostylev, M.; Kim, D.Y.; Smalley, N.E.; Salukhe, I.; Greenberg, E.P.; Dandekar, A.A. Evolution of the Pseudomonas aeruginosa quorum-sensing hierarchy. Proc. Natl. Acad. Sci. USA 2019, 116, 7027–7032. [Google Scholar] [CrossRef]
- Feltner, J.B.; Wolter, D.J.; Pope, C.E.; Groleau, M.C.; Smalley, N.E.; Greenberg, E.P.; Mayer-Hamblett, N.; Burns, J.; Déziel, E.; Hoffman, L.R.; et al. LasR Variant Cystic Fibrosis Isolates Reveal an Adaptable Quorum-Sensing Hierarchy in Pseudomonas aeruginosa. mBio 2016, 7, e01513-16. [Google Scholar] [CrossRef]
- Vincent, A.T.; Freschi, L.; Jeukens, J.; Kukavica-Ibrulj, I.; Emond-Rheault, J.-G.; Leduc, A.; Boyle, B.; Jean-Pierre, F.; Groleau, M.-C.; Déziel, E.; et al. Genomic characterization of environmental Pseudomonas aeruginosa isolated from dental unit waterlines revealed the insertion sequence ISPa11 as a chaotropic element. FEMS Microbiol. Ecol. 2017, 93, fix106. [Google Scholar] [CrossRef]
- Groleau, M.C.; Taillefer, H.; Vincent, A.T.; Constant, P.; Déziel, E. Pseudomonas aeruginosa isolates defective in function of the LasR quorum sensing regulator are frequent in diverse environmental niches. Environ. Microbiol. 2021, 24, 1062–1075. [Google Scholar] [CrossRef]
- Ruiz-Roldán, L.; Rojo-Bezares, B.; Lozano, C.; López, M.; Chichón, G.; Torres, C.; Sáenz, Y. Occurrence of Pseudomonas spp. in Raw Vegetables: Molecular and Phenotypical Analysis of Their Antimicrobial Resistance and Virulence-Related Traits. Int. J. Mol. Sci. 2021, 22, 12626. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, L.; Rao, X.; Wang, J.; Yu, H.; Jiang, J.; Zhou, W.; Xiao, Y.; Li, M.; Zhang, Y.; et al. Characterization of lasR-deficient clinical isolates of Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 13344. [Google Scholar] [CrossRef]
- Scribner, M.R.; Stephens, A.C.; Huong, J.L.; Richardson, A.R.; Cooper, V.S. The Nutritional Environment Is Sufficient to Select Coexisting Biofilm and Quorum Sensing Mutants of Pseudomonas aeruginosa. J. Bacteriol. 2022, 204, e0044421. [Google Scholar] [CrossRef] [PubMed]
- Williams McMackin, E.A.; Djapgne, L.; Corley, J.M.; Yahr, T.L. Fitting Pieces into the Puzzle of Pseudomonas aeruginosa Type III Secretion System Gene Expression. J. Bacteriol. 2019, 201, e00209-19. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.J.; Vasil, A.I.; Johnson, Z.; Vasil, M.L. Genetic and biochemical analyses of a eukaryotic-like phospholipase D of Pseudomonas aeruginosa suggest horizontal acquisition and a role for persistence in a chronic pulmonary infection model. Mol. Microbiol. 2001, 39, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Sims, E.H.; Spencer, D.H.; Kaul, R.; Olson, M.V. Evidence for diversifying selection at the pyoverdine locus of Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 2138–2147. [Google Scholar] [CrossRef]
- Burrows, L.L. Pseudomonas aeruginosa twitching motility: Type IV pili in action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef]
- Nguyen, Y.; Jackson, S.G.; Aidoo, F.; Junop, M.; Burrows, L.L. Structural characterization of novel Pseudomonas aeruginosa type IV pilins. J. Mol. Biol. 2010, 395, 491–503. [Google Scholar] [CrossRef]
- Kiyaga, S.; Kyany’a, C.; Muraya, A.W.; Smith, H.J.; Mills, E.G.; Kibet, C.; Mboowa, G.; Musila, L. Genetic Diversity, Distribution, and Genomic Characterization of Antibiotic Resistance and Virulence of Clinical Pseudomonas aeruginosa Strains in Kenya. Front. Microbiol. 2022, 13, 835403. [Google Scholar] [CrossRef]
- Giltner, C.L.; Rana, N.; Lunardo, M.N.; Hussain, A.Q.; Burrows, L.L. Evolutionary and functional diversity of the Pseudomonas type IVa pilin island. Environ. Microbiol. 2010, 13, 250–264. [Google Scholar] [CrossRef]
- Asikyan, M.L.; Kus, J.V.; Burrows, L.L. Novel proteins that modulate type IV pilus retraction dynamics in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7022–7034. [Google Scholar] [CrossRef] [PubMed]
- Kus, J.V.; Kelly, J.; Tessier, L.; Harvey, H.; Cvitkovitch, D.G.; Burrows, L.L. Modification of Pseudomonas aeruginosa Pa5196 type IV Pilins at multiple sites with D-Araf by a novel GT-C family Arabinosyltransferase, TfpW. J. Bacteriol. 2008, 190, 7464–7478. [Google Scholar] [CrossRef] [PubMed]
- Kung, V.L.; Ozer, E.A.; Hauser, A.R. The accessory genome of Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2010, 74, 621–641. [Google Scholar] [CrossRef] [PubMed]
- Horna, G.; Quezada, K.; Ramos, S.; Mosqueda, N.; Rubio, M.; Guerra, H.; Ruiz, J. Specific type IV pili groups in clinical isolates of Pseudomonas aeruginosa. Int. Microbiol. 2019, 22, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Stintzi, A.; De Vos, D.; Cornelis, P.; Tappe, R.; Taraz, K.; Budzikiewicz, H. Use of siderophores to type pseudomonads: The three Pseudomonas aeruginosa pyoverdine systems. Microbiology 1997, 143, 35–43. [Google Scholar] [CrossRef]
- Cornelis, P.; Hohnadel, D.; Meyer, J.M. Evidence for different pyoverdine-mediated iron uptake systems among Pseudomonas aeruginosa strains. Infect. Immun. 1989, 57, 3491–3497. [Google Scholar] [CrossRef]
- Baysse, C.; Meyer, J.M.; Plesiat, P.; Geoffroy, V.; Michel-Briand, Y.; Cornelis, P. Uptake of pyocin S3 occurs through the outer membrane ferripyoverdine type II receptor of Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 3849–3851. [Google Scholar] [CrossRef]
- Behrens, H.M.; Lowe, E.D.; Gault, J.; Housden, N.G.; Kaminska, R.; Weber, T.M.; Thompson, C.M.A.; Mislin, G.L.A.; Schalk, I.J.; Walker, D.; et al. Pyocin S5 Import into Pseudomonas aeruginosa Reveals a Generic Mode of Bacteriocin Transport. mBio 2020, 11, e03230-19. [Google Scholar] [CrossRef]
- Mulet, X.; Cabot, G.; Ocampo-Sosa, A.A.; Domínguez, M.A.; Zamorano, L.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; Peña, C.; et al. Biological Markers of Pseudomonas aeruginosa Epidemic High-Risk Clones. Antimicrob. Agents Chemother. 2013, 57, 5527–5535. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Rojo-Bezares, B.; Cavalié, L.; Dubois, D.; Oswald, E.; Torres, C.; Sáenz, Y. Characterization of carbapenem resistance mechanisms and integrons in Pseudomonas aeruginosa strains from blood samples in a French hospital. J. Med. Microbiol. 2016, 65, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Estepa, V.; Rojo-Bezares, B.; Torres, C.; Sáenz, Y. Faecal carriage of Pseudomonas aeruginosa in healthy humans: Antimicrobial susceptibility and global genetic lineages. FEMS Microbiol. Ecol. 2014, 89, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Heras, J.; Domínguez, C.; Mata, E.; Pascual, V.; Lozano, C.; Torres, C.; Zarazaga, M. GelJ—A tool for analyzing DNA fingerprint gel images. BMC Bioinform. 2015, 16, 270. [Google Scholar] [CrossRef] [PubMed]
- Casado-García, Á.; Chichón, G.; Domínguez, C.; García-Domínguez, M.; Heras, J.; Inés, A.; López, M.; Mata, E.; Pascual, V.; Sáenz, Y. MotilityJ: An open-source tool for the classification and segmentation of bacteria on motility images. Comput. Biol. Med. 2021, 136, 104673. [Google Scholar] [CrossRef] [PubMed]
- Semmler, A.B.; Whitchurch, C.B.; Mattick, J.S. A re-examination of twitching motility in Pseudomonas aeruginosa. Microbiology 1999, 145, 2863–2873. [Google Scholar] [CrossRef]
- Cullen, L.; Weiser, R.; Olszak, T.; Maldonado, R.F.; Moreira, A.S.; Slachmuylders, L.; Brackman, G.; Paunova-Krasteva, T.S.; Zarnowiec, P.; Czerwonka, G.; et al. Phenotypic characterization of an international Pseudomonas aeruginosa reference panel: Strains of cystic fibrosis (CF) origin show less in vivo virulence than non-CF strains. Microbiology 2015, 161, 1961–1977. [Google Scholar] [CrossRef]
- Vielva, L.; de Toro, M.; Lanza, V.F.; de la Cruz, F. PLACNETw: A web-based tool for plasmid reconstruction from bacterial genomes. Bioinformatics 2017, 33, 3796–3798. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Lanza, V.F.; de Toro, M.; Garcillán-Barcia, M.P.; Mora, A.; Blanco, J.; Coque, T.M.; de la Cruz, F. Plasmid flux in Escherichia coli ST131 sublineages, analyzed by Plasmid Constellation Network (PLACNET), a new method for plasmid reconstruction from whole genome sequences. PLoS Genet. 2014, 10, e1004766. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Madaha, E.L.; Mienie, C.; Gonsu, H.K.; Bughe, R.N.; Fonkoua, M.C.; Mbacham, W.F.; Alayande, K.A.; Bezuidenhout, C.C.; Ateba, C.N. Whole-genome sequence of multi-drug resistant Pseudomonas aeruginosa strains UY1PSABAL and UY1PSABAL2 isolated from human broncho-alveolar lavage, Yaoundé, Cameroon. PLoS ONE 2020, 15, e0238390. [Google Scholar] [CrossRef]
- López-Causapé, C.; Sommer, L.M.; Cabot, G.; Rubio, R.; Ocampo-Sosa, A.A.; Johansen, H.K.; Figuerola, J.; Cantón, R.; Kidd, T.J.; Molin, S.; et al. Evolution of the Pseudomonas aeruginosa mutational resistome in an international Cystic Fibrosis clone. Sci. Rep. 2017, 7, 5555. [Google Scholar] [CrossRef]
- Díaz-Caballero, J.D.; Clark, S.T.; Coburn, B.; Zhang, Y.; Wang, P.W.; Donaldson, S.L.; Tullis, D.E.; Yau, Y.C.W.; Waters, V.J.; Hwang, D.M.; et al. Selective Sweeps and Parallel Pathoadaptation Drive Pseudomonas aeruginosa Evolution in the Cystic Fibrosis Lung. mBio 2015, 6, e00981-15. [Google Scholar] [CrossRef] [PubMed]
- Winsor, G.L.; Van Rossum, T.; Lo, R.; Khaira, B.; Whiteside, M.D.; Hancock, R.E.W.; Brinkman, F.S.L. Pseudomonas Genome Database: Facilitating user-friendly, comprehensive comparisons of microbial genomes. Nucleic Acids Res. 2008, 37, D483–D488. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Urbach, J.M.; Wu, G.; Liberati, N.T.; Feinbaum, R.L.; Miyata, S.; Diggins, L.T.; He, J.; Saucier, M.; Deziel, E.; et al. Genomic analysis reveals that Pseudomonas aeruginosa virulence is combinatorial. Genome Biol. 2006, 7, R90. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sample | Origin a | PFGE Pattern | Serotype b | Resistance Phenotype c | MIC (mg/L) c | Virulotype d | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PTZ | CAZ | FEP | IMP | MER | DOR | GEN | TOB | CIP | LEV | CT | ATM | |||||||
| G42 | Healthy volunteer | NC | P2 | O:3 | multiS | ≤8 | 4 | 4 | 2 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G44 | Healthy volunteer | NC | P4 | PoliA | modR | ≤8 | 16 | 4 | 4 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G73 | Vegetable (lettuce) | NC | P5 | O:3 | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 4 | V1 |
| G92 | Vegetable (chard) | NC | P6 | O:3 | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G105 | River water | NC | P9b | O:3 | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G115 | River water | NC | P9b | O:3 | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G148 | Blood | C | P7 | O:3 | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 4 | V1 |
| G178 | Blood | C | P8 | AutoA | multiS | ≤8 | 2 | 4 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 4 | V1 |
| G179 e | Blood | C | P9a | AutoA | XDR, DTR | >64 | >16 | >16 | >8 | 8 | >4 | 8 | ≤2 | 2 | 4 | ≤2 | >16 | V1 |
| G224 | Respiratory | C | P3 | O:3 | multiS | 16 | 2 | 2 | ≤1 | ≤1 | ≤1 | 4 | ≤2 | ≤0.5 | ≤1 | ≤2 | 8 | V1 |
| G245 e | Respiratory | C | P1 | PoliA | multiS | ≤8 | ≤1 | ≤1 | ≤1 | ≤1 | ≤1 | ≤2 | ≤2 | ≤0.5 | ≤1 | ≤2 | ≤1 | V2 |
| Strains | Reads | Assembly Parameters | Genetic Elements | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of Contigs | Contig Maximum Length (bp) | Total Bases in Contigs > 1 kb | N50 (bp) | Genome Size (bp) | GC Content (%) | CDS | Genes | rRNA | tRNA | tmRNA | ||
| G42 | 5,890,730 | 87 | 726,697 | 6,748,134 | 353,893 | 6,757,744 | 66.04 | 6166 | 6241 | 3 | 71 | 1 |
| G44 | 5,143,091 | 76 | 739,379 | 6,550,288 | 612,825 | 6,556,612 | 66.29 | 6000 | 6076 | 3 | 72 | 1 |
| G73 | 4,765,846 | 66 | 1,027,638 | 6,349,927 | 704,215 | 6,355,219 | 66.47 | 5786 | 5861 | 3 | 71 | 1 |
| G92 | 6,001,411 | 68 | 1,029,741 | 6,402,390 | 726,878 | 6,410,342 | 66.42 | 5840 | 5913 | 3 | 69 | 1 |
| G105 | 5,647,144 | 46 | 768,785 | 6,241,289 | 621,817 | 6,241,882 | 66.54 | 5659 | 5734 | 3 | 71 | 1 |
| G115 | 5,060,730 | 62 | 768,777 | 6,302,900 | 621,730 | 6,309,215 | 66.43 | 5741 | 5816 | 3 | 71 | 1 |
| G148 | 5,581,661 | 69 | 931,567 | 6,423,973 | 695,550 | 6,430,217 | 66.35 | 5856 | 5934 | 6 | 71 | 1 |
| G178 | 17,116,583 | 59 | 1,439,870 | 6,404,614 | 606,103 | 6,412,019 | 66.43 | 5825 | 5901 | 4 | 71 | 1 |
| G179 | 7,309,267 | 78 | 729,747 | 6,244,365 | 427,443 | 6,251,772 | 66.53 | 5662 | 5739 | 3 | 73 | 1 |
| G224 | 5,613,901 | 61 | 1,454,566 | 6,350,601 | 804,729 | 6,356,169 | 66.48 | 5821 | 5896 | 3 | 71 | 1 |
| G245 | 11,589,531 | 109 | 726,712 | 6,794,368 | 409,397 | 6,805,979 | 66.07 | 6192 | 6267 | 3 | 71 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chichón, G.; López, M.; de Toro, M.; Ruiz-Roldán, L.; Rojo-Bezares, B.; Sáenz, Y. Spread of Pseudomonas aeruginosa ST274 Clone in Different Niches: Resistome, Virulome, and Phylogenetic Relationship. Antibiotics 2023, 12, 1561. https://doi.org/10.3390/antibiotics12111561
Chichón G, López M, de Toro M, Ruiz-Roldán L, Rojo-Bezares B, Sáenz Y. Spread of Pseudomonas aeruginosa ST274 Clone in Different Niches: Resistome, Virulome, and Phylogenetic Relationship. Antibiotics. 2023; 12(11):1561. https://doi.org/10.3390/antibiotics12111561
Chicago/Turabian StyleChichón, Gabriela, María López, María de Toro, Lidia Ruiz-Roldán, Beatriz Rojo-Bezares, and Yolanda Sáenz. 2023. "Spread of Pseudomonas aeruginosa ST274 Clone in Different Niches: Resistome, Virulome, and Phylogenetic Relationship" Antibiotics 12, no. 11: 1561. https://doi.org/10.3390/antibiotics12111561
APA StyleChichón, G., López, M., de Toro, M., Ruiz-Roldán, L., Rojo-Bezares, B., & Sáenz, Y. (2023). Spread of Pseudomonas aeruginosa ST274 Clone in Different Niches: Resistome, Virulome, and Phylogenetic Relationship. Antibiotics, 12(11), 1561. https://doi.org/10.3390/antibiotics12111561