

Semisynthetic Amides of Amphotericin B and Nystatin A1: A Comparative Study of In Vitro Activity/Toxicity Ratio in Relation to Selectivity to Ergosterol Membranes
 , , ,
, , ,
Abstract
1. Introduction
2. Results
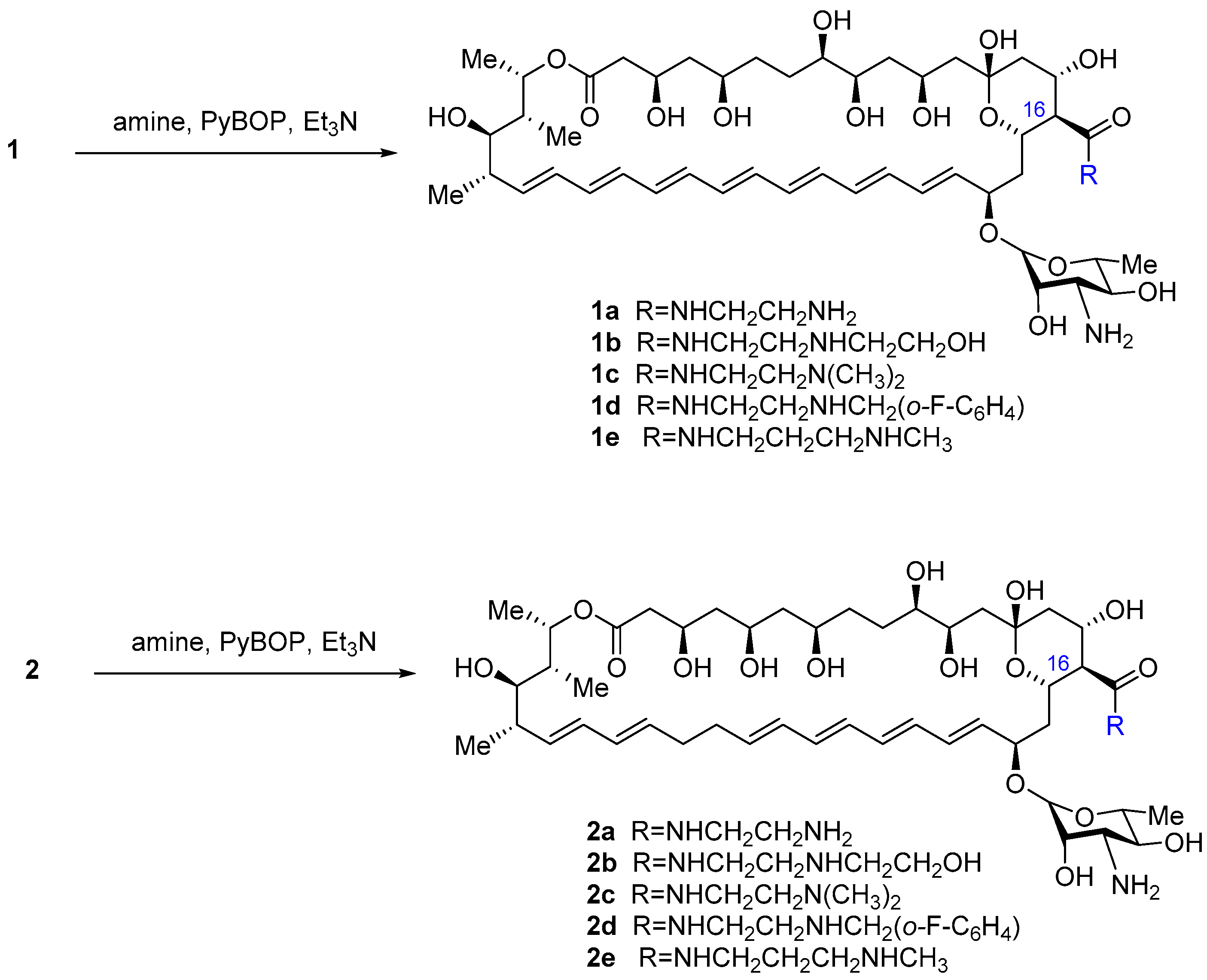
2.1. Synthesis of Polyene Amides
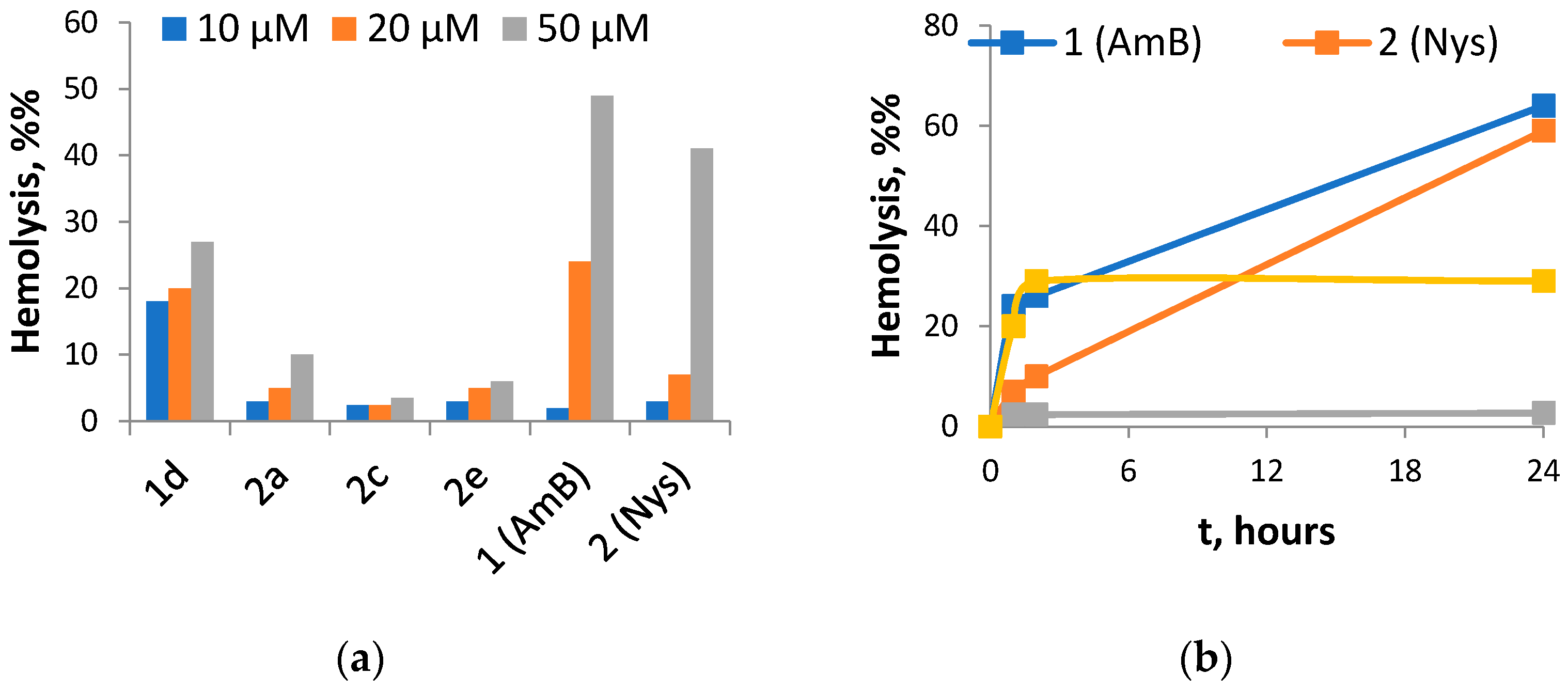
2.2. In Vitro Antifungal Activity and Toxicity toward Mammalian Cells
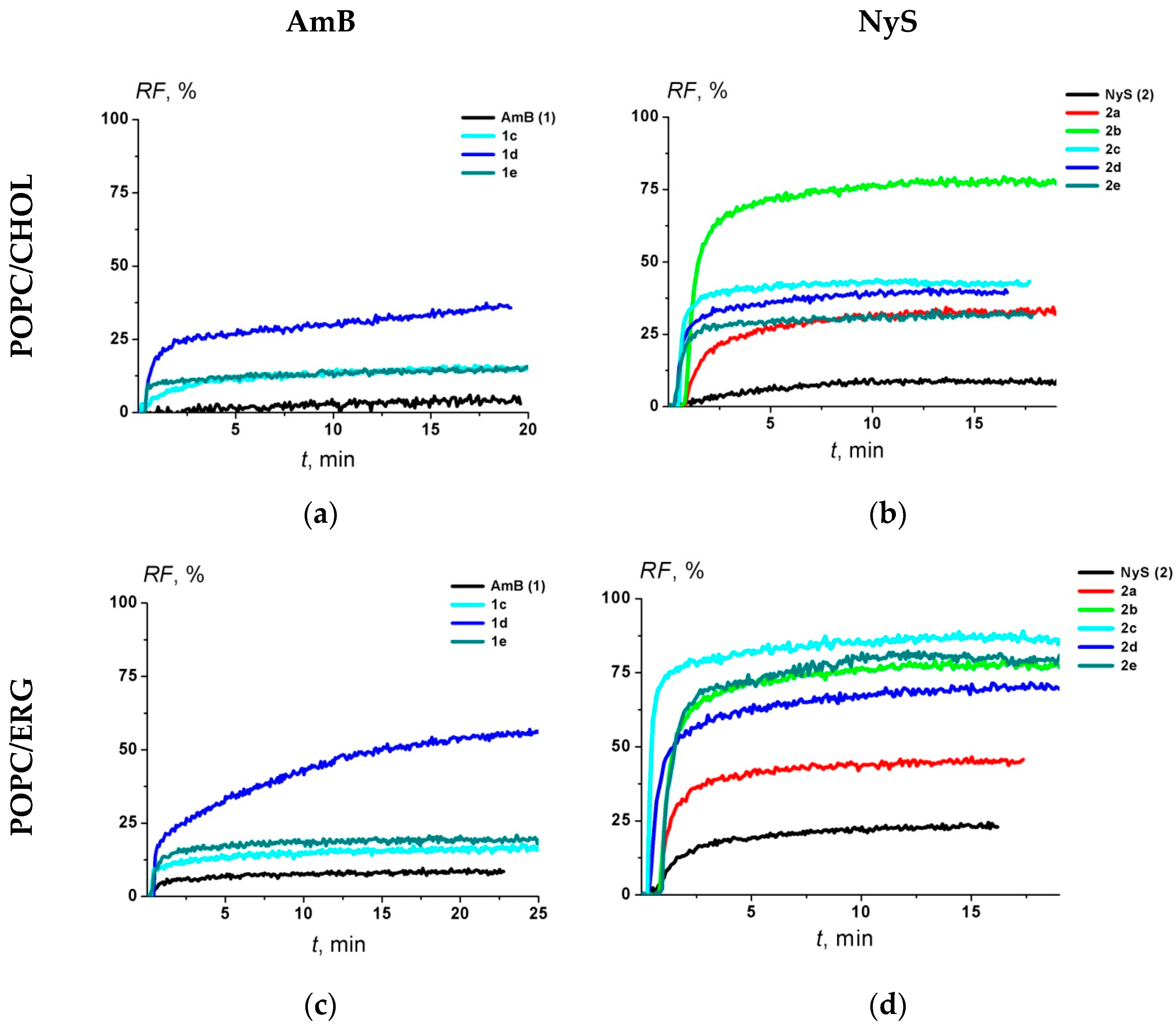
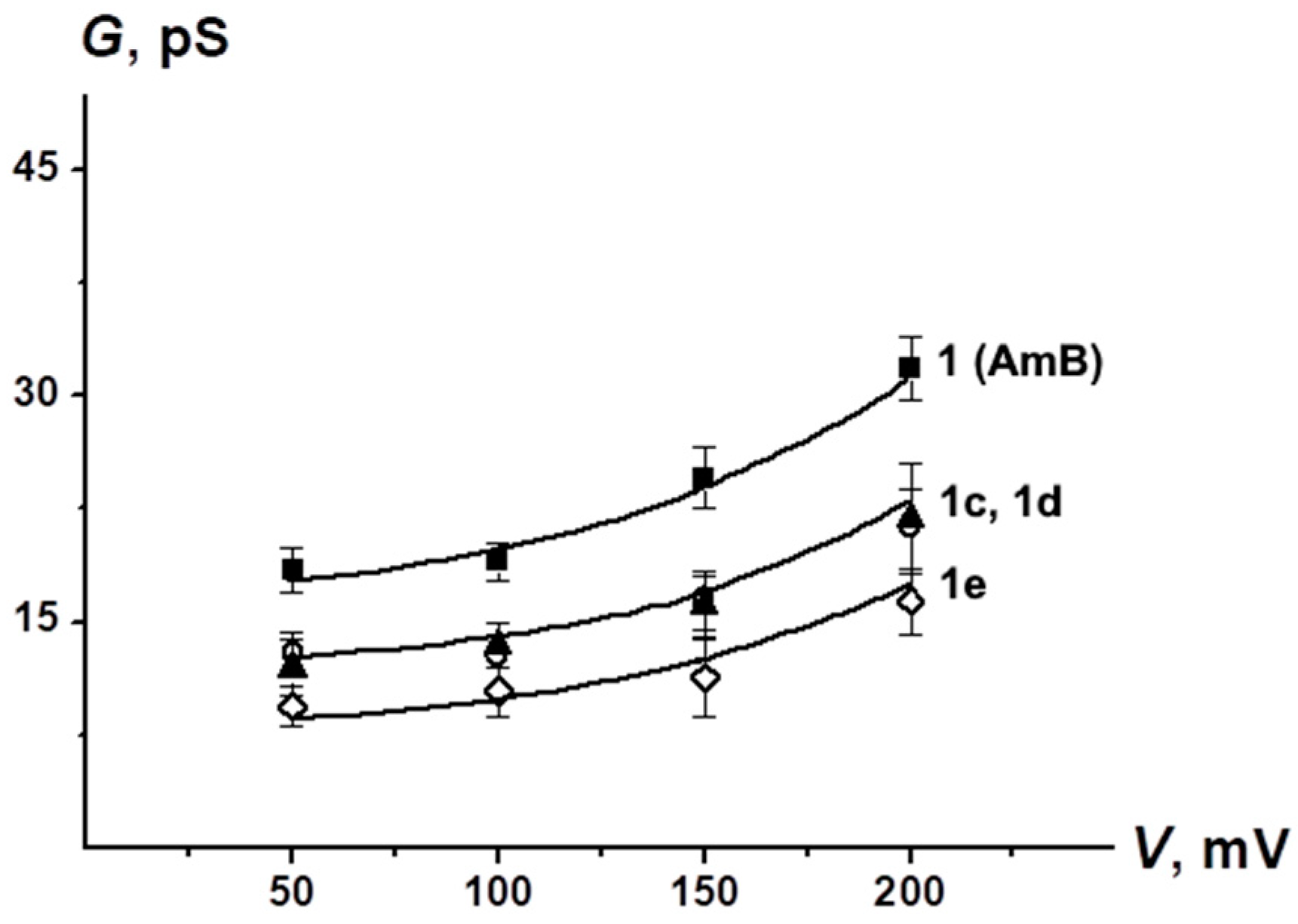
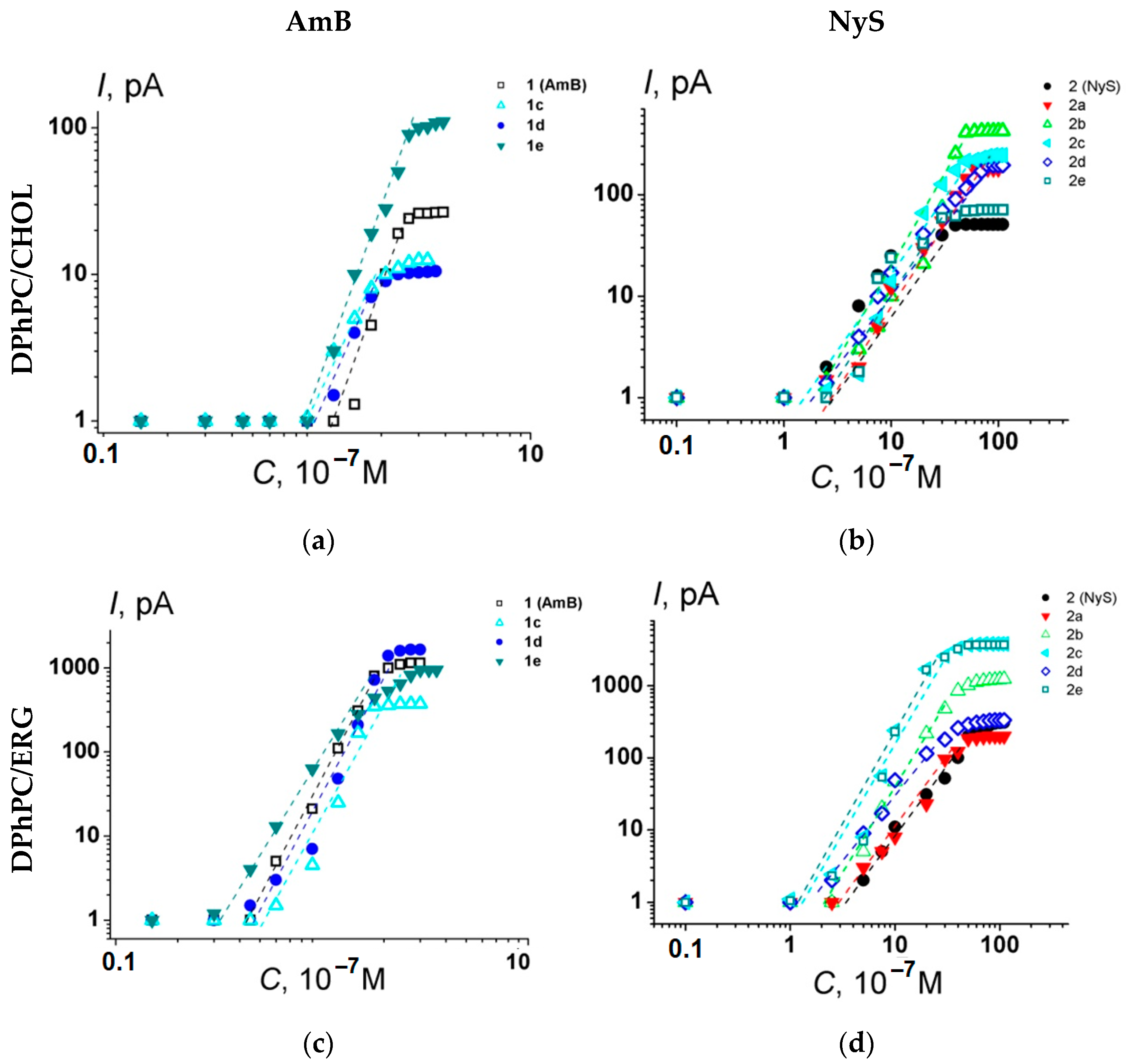
2.3. Electrophysiological Experiments
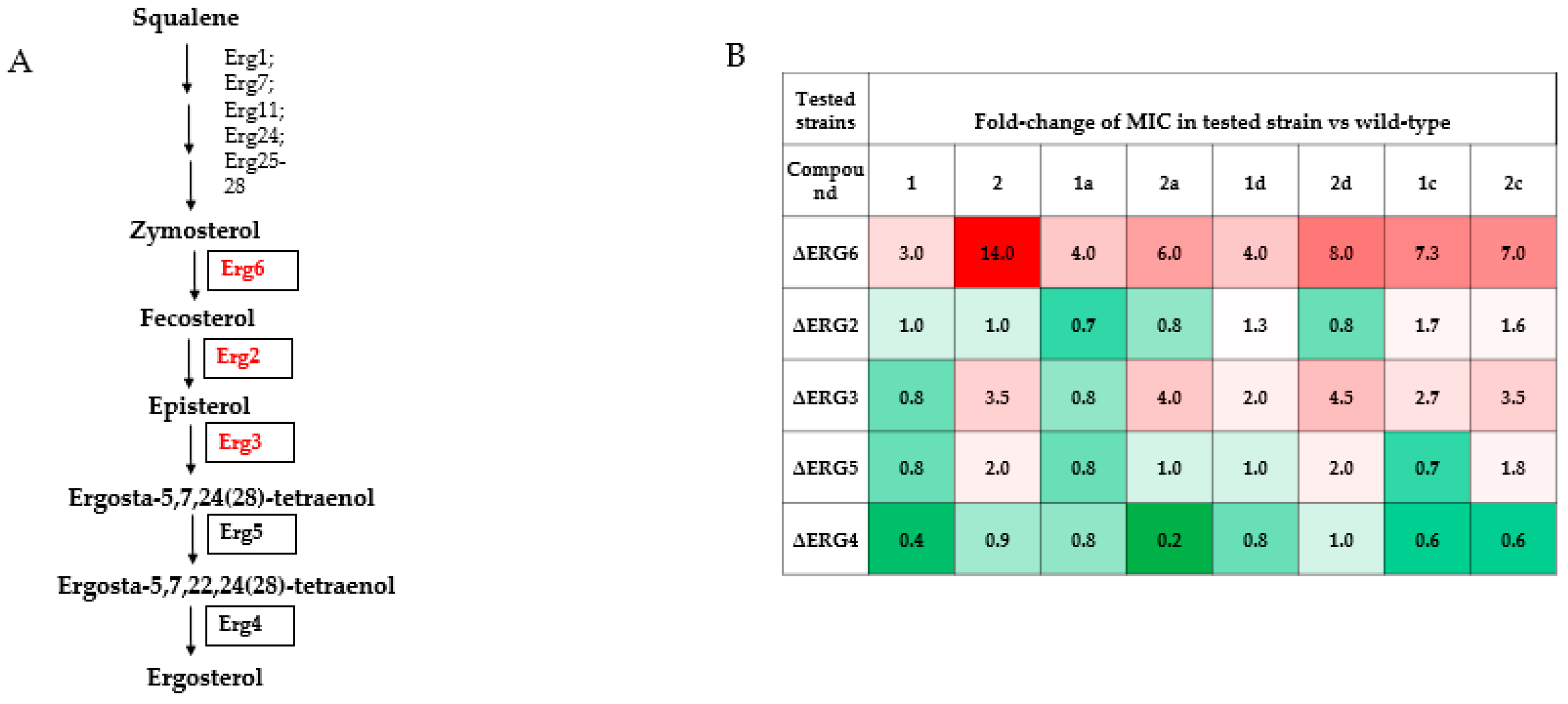
2.4. Comparison of Susceptibility of Ergosterol-Pathway Mutants to Polyene Derivatives
3. Discussion
4. Materials and Methods
4.1. General
4.2. Carboxamides of Amphotericin B and Nystatin (1c,d,e, 2a–e) (General Method)
4.3. Solubility Testing
4.4. Antifungal Susceptibility Testing
4.4.1. Organisms
4.4.2. Preparation of Chemical Samples (for Pathogen Exposure)
4.5. Cell Culture and Antiproliferative Activity
4.6. Testing of Hemolysis Activity
4.7. Calcein Release from Large Unilamellar Vesicles
4.8. Registration of Ion Channels in Planar Lipid Bilayers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida Albicans and Emerging Non-Albicans Candida Species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Parra, J.; Muiño-Miguez, A.; Bendala-Estrada, A.D.; Ramos-Martínez, A.; Muñez-Rubio, E.; Fernández Carracedo, E.; Tejada Montes, J.; Rubio-Rivas, M.; Arnalich-Fernandez, F.; Beato Pérez, J.L.; et al. Inappropriate Antibiotic Use in the COVID-19 Era: Factors Associated with Inappropriate Prescribing and Secondary Complications. Analysis of the Registry SEMI-COVID. PLoS ONE 2021, 16, e0251340. [Google Scholar] [CrossRef] [PubMed]
- Adebisi, Y.A.; Alaran, A.J.; Okereke, M.; Oke, G.I.; Amos, O.A.; Olaoye, O.C.; Oladunjoye, I.; Olanrewaju, A.Y.; Ukor, N.A.; Don Eliseo Lucero-Prisno, I. COVID-19 and Antimicrobial Resistance: A Review. Infect. Dis. Res. Treat. 2021, 14, 11786337211033870. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Rossolini, G.M.; Schultsz, C.; Tacconelli, E.; Murthy, S.; Ohmagari, N.; Holmes, A.; Bachmann, T.; Goossens, H.; Canton, R.; et al. Antimicrobial Resistance Research in a Post-Pandemic World: Insights on Antimicrobial Resistance Research in the COVID-19 Pandemic. J. Glob. Antimicrob. Resist. 2021, 25, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Nieuwlaat, R.; Mbuagbaw, L.; Mertz, D.; Burrows, L.L.; Bowdish, D.M.E.; Moja, L.; Wright, G.D.; Schünemann, H.J. Coronavirus Disease 2019 and Antimicrobial Resistance: Parallel and Interacting Health Emergencies. Clin. Infect. Dis. 2021, 72, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Cavassin, F.B.; Baú-Carneiro, J.L.; Vilas-Boas, R.R.; Queiroz-Telles, F. Sixty Years of Amphotericin B: An Overview of the Main Antifungal Agent Used to Treat Invasive Fungal Infections. Infect. Dis. Ther. 2021, 10, 115–147. [Google Scholar] [CrossRef] [PubMed]
- Steinbrink, J.M.; Miceli, M.H. Mucormycosis. Infect. Dis. Clin. N. Am. 2021, 35, 435–452. [Google Scholar] [CrossRef]
- Pal, R.; Singh, B.; Bhadada, S.K.; Banerjee, M.; Bhogal, R.S.; Hage, N.; Kumar, A. COVID-19-Associated Mucormycosis: An Updated Systematic Review of Literature. Mycoses 2021, 64, 1452–1459. [Google Scholar] [CrossRef]
- Haro-Reyes, T.; Díaz-Peralta, L.; Galván-Hernández, A.; Rodríguez-López, A.; Rodríguez-Fragoso, L.; Ortega-Blake, I. Polyene Antibiotics Physical Chemistry and Their Effect on Lipid Membranes; Impacting Biological Processes and Medical Applications. Membranes 2022, 12, 681. [Google Scholar] [CrossRef] [PubMed]
- Ermishkin, L.N.; Kasumov, K.M.; Potzeluyev, V.M. Single Ionic Channels Induced in Lipid Bilayers by Polyene Antibiotics Amphotericin B and Nystatine. Nature 1976, 262, 698–699. [Google Scholar] [CrossRef]
- Andreoli, T.E.; Monahan, M. The Interaction of Polyene Antibiotics with Thin Lipid Membranes. J. Gen. Physiol. 1968, 52, 300–325. [Google Scholar] [CrossRef]
- Holz, R.; Finkelstein, A. The Water and Nonelectrolyte Permeability Induced in Thin Lipid Membranes by the Polyene Antibiotics Nystatin and Amphotericin B. J. Gen. Physiol. 1970, 56, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Dennis, V.W.; Stead, N.W.; Andreoli, T.E. Molecular Aspects of Polyene- and Sterol-Dependent Pore Formation in Thin Lipid Membranes. J. Gen. Physiol. 1970, 55, 375–400. [Google Scholar] [CrossRef]
- De Kruijff, B.; Demel, R.A. Polyene Antibiotic-Sterol Interactions in Membranes of Acholeplasma Laidlawii Cells and Lecithin Liposomes. III. Molecular Structure of the Polyene Antibiotic-Cholesterol Complexes. Biochim. Et Biophys. Acta (BBA) Biomembr. 1974, 339, 57–70. [Google Scholar] [CrossRef]
- Volmer, A.A.; Szpilman, A.M.; Carreira, E.M. Synthesis and Biological Evaluation of Amphotericin B Derivatives. Nat. Prod. Rep. 2010, 27, 1329–1349. [Google Scholar] [CrossRef]
- Solovieva, S.E.; Olsufyeva, E.N.; Preobrazhenskaya, M.N. Chemical Modification of Antifungal Polyene Macrolide Antibiotics. Russ. Chem. Rev. 2011, 80, 103–126. [Google Scholar] [CrossRef]
- Omelchuk, O.A.; Tevyashova, A.N.; Shchekotikhin, A.E. Recent advances in antifungal drug discovery based on polyene macrolide antibiotics. Russ. Chem. Rev. 2018, 87, 1206. [Google Scholar] [CrossRef]
- Welscher, Y.M.; ten Napel, H.H.; Balagué, M.M.; Souza, C.M.; Riezman, H.; de Kruijff, B.; Breukink, E. Natamycin Blocks Fungal Growth by Binding Specifically to Ergosterol without Permeabilizing the Membrane*. J. Biol. Chem. 2008, 283, 6393–6401. [Google Scholar] [CrossRef]
- te Welscher, Y.M.; van Leeuwen, M.R.; de Kruijff, B.; Dijksterhuis, J.; Breukink, E. Polyene Antibiotic That Inhibits Membrane Transport Proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 11156–11159. [Google Scholar] [CrossRef]
- Gray, K.C.; Palacios, D.S.; Dailey, I.; Endo, M.M.; Uno, B.E.; Wilcock, B.C.; Burke, M.D. Amphotericin Primarily Kills Yeast by Simply Binding Ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.M.; Clay, M.C.; Cioffi, A.G.; Diaz, K.A.; Hisao, G.S.; Tuttle, M.D.; Nieuwkoop, A.J.; Comellas, G.; Maryum, N.; Wang, S.; et al. Amphotericin Forms an Extramembranous and Fungicidal Sterol Sponge. Nat. Chem. Biol. 2014, 10, 400–406. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, J.; Li, X.; Xiao, E.; Lange, J.D.; Rienstra, C.M.; Burke, M.D.; Mitchell, D.A. Sterol Sponge Mechanism Is Conserved for Glycosylated Polyene Macrolides. ACS Cent. Sci. 2021, 7, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Endo, M.M.; Cioffi, A.G.; Burke, M.D. Our Path to Less Toxic Amphotericins. Synlett 2016, 27, 337–354. [Google Scholar]
- Delhom, R.; Nelson, A.; Laux, V.; Haertlein, M.; Knecht, W.; Fragneto, G.; Wacklin-Knecht, H.P. The Antifungal Mechanism of Amphotericin B Elucidated in Ergosterol and Cholesterol-Containing Membranes Using Neutron Reflectometry. Nanomaterials 2020, 10, 2439. [Google Scholar] [CrossRef]
- Kamiński, D.M. Recent Progress in the Study of the Interactions of Amphotericin B with Cholesterol and Ergosterol in Lipid Environments. Eur. Biophys. J. 2014, 43, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Umegawa, Y.; Yamagami, M.; Suzuki, T.; Tsuchikawa, H.; Hanashima, S.; Matsumori, N.; Murata, M. The Perpendicular Orientation of Amphotericin B Methyl Ester in Hydrated Lipid Bilayers Supports the Barrel-Stave Model. Biochemistry 2019, 58, 2282–2291. [Google Scholar] [CrossRef] [PubMed]
- Umegawa, Y.; Yamamoto, T.; Dixit, M.; Funahashi, K.; Seo, S.; Nakagawa, Y.; Suzuki, T.; Matsuoka, S.; Tsuchikawa, H.; Hanashima, S.; et al. Amphotericin B Assembles into Seven-Molecule Ion Channels: An NMR and Molecular Dynamics Study. Sci. Adv. 2022, 8, eabo2658. [Google Scholar] [CrossRef]
- Ferreira, G.F.; de Baltazar, L.M.; Santos, J.R.A.; Monteiro, A.S.; de Fraga, L.A.O.; Resende-Stoianoff, M.A.; Santos, D.A. The Role of Oxidative and Nitrosative Bursts Caused by Azoles and Amphotericin B against the Fungal Pathogen Cryptococcus Gattii. J. Antimicrob. Chemother. 2013, 68, 1801–1811. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Trevijano-Contador, N.; Román, E.; Sánchez-Fresneda, R.; Casas, C.; Herrero, E.; Argüelles, J.C.; Pla, J.; Cuenca-Estrella, M.; Zaragoza, O. The Production of Reactive Oxygen Species Is a Universal Action Mechanism of Amphotericin B against Pathogenic Yeasts and Contributes to the Fungicidal Effect of This Drug. Antimicrob. Agents Chemother. 2014, 58, 6627–6638. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, Z.; Han, X.; Tang, J.; Wang, J.; Dong, S.; Wang, E. Ion Channel Behavior of Amphotericin B in Sterol-Free and Cholesterol- or Ergosterol-Containing Supported Phosphatidylcholine Bilayer Model Membranes Investigated by Electrochemistry and Spectroscopy. Biophys. J. 2002, 83, 3245–3255. [Google Scholar] [CrossRef]
- Barwicz, J.; Christian, S.; Gruda, I. Effects of the Aggregation State of Amphotericin B on Its Toxicity to Mice. Antimicrob. Agents Chemother. 1992, 36, 2310–2315. [Google Scholar] [CrossRef]
- Antillón, A.; de Vries, A.H.; Espinosa-Caballero, M.; Falcón-González, J.M.; Flores Romero, D.; González–Damián, J.; Jiménez-Montejo, F.E.; León-Buitimea, A.; López-Ortiz, M.; Magaña, R.; et al. An Amphotericin B Derivative Equally Potent to Amphotericin B and with Increased Safety. PLoS ONE 2016, 11, e0162171. [Google Scholar] [CrossRef]
- Tevyashova, A.N.; Bychkova, E.N.; Solovieva, S.E.; Zatonsky, G.V.; Grammatikova, N.E.; Isakova, E.B.; Mirchink, E.P.; Treshchalin, I.D.; Pereverzeva, E.R.; Bykov, E.E.; et al. Discovery of Amphamide, a Drug Candidate for the Second Generation of Polyene Antibiotics. ACS Infect. Dis. 2020, 6, 2029–2044. [Google Scholar] [CrossRef] [PubMed]
- Janout, V.; Schell, W.A.; Thévenin, D.; Yu, Y.; Perfect, J.R.; Regen, S.L. Taming Amphotericin B. Bioconjugate Chem. 2015, 26, 2021–2024. [Google Scholar] [CrossRef] [PubMed]
- Hervé, M.; Debouzy, J.C.; Borowski, E.; Cybulska, B.; Gary-Bobo, C.M. The Role of the Carboxyl and Amino Groups of Polyene Macrolides in Their Interactions with Sterols and Their Selective Toxicity. A 31P-NMR Study. Biochim. Biophys. Acta (BBA) Biomembr. 1989, 980, 261–272. [Google Scholar] [CrossRef]
- Amir, A.; Levin-Khalifa, M.; Dvash, T. Water-Soluble Nystatin and Derivative. ACS Med. Chem. Lett. 2022, 13, 182–187. [Google Scholar] [CrossRef]
- Hąc-Wydro, K.; Dynarowicz-Łątka, P.; Grzybowska, J.; Borowski, E. N-(1-Piperidinepropionyl)Amphotericin B Methyl Ester (PAME)—A New Derivative of the Antifungal Antibiotic Amphotericin B: Searching for the Mechanism of Its Reduced Toxicity. J. Colloid Interface Sci. 2005, 287, 476–484. [Google Scholar] [CrossRef]
- Tevyashova, A.N.; Efimova, S.S.; Alexandrov, A.I.; Ghazy, E.S.M.O.; Bychkova, E.N.; Solovieva, S.E.; Zatonsky, G.B.; Grammatikova, N.E.; Dezhenkova, L.G.; Pereverzeva, E.R.; et al. Semisynthetic Amides of Polyene Antibiotic Natamycin. ACS Infect. Dis. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Lee, M.-J.; Kong, D.; Han, K.; Sherman, D.H.; Bai, L.; Deng, Z.; Lin, S.; Kim, E.-S. Structural Analysis and Biosynthetic Engineering of a Solubility-Improved and Less-Hemolytic Nystatin-like Polyene in Pseudonocardia Autotrophica. Appl. Microbiol. Biotechnol. 2012, 95, 157–168. [Google Scholar] [CrossRef]
- Van Arnam, E.B.; Ruzzini, A.C.; Sit, C.S.; Horn, H.; Pinto-Tomás, A.A.; Currie, C.R.; Clardy, J. Selvamicin, an Atypical Antifungal Polyene from Two Alternative Genomic Contexts. Proc. Natl. Acad. Sci. USA 2016, 113, 12940–12945. [Google Scholar] [CrossRef] [PubMed]
- Yalkowsky, R.M.; Dannenfleser, S.H. Aquasol Database of Aqueous Solubility, Version 5; University of Arizona, College of Pharmacy: Tuscon, Arizona, 1992.
- EUCAST Definitive Document E.DEF 7.3.2 Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7.3.2_Yeast_testing_definitive_revised_2020.pdf (accessed on 9 January 2023).
- EUCAST Definitive Document E.DEF 9.3.2 Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Conidia Forming Moulds. Available online: https://www.aspergillus.org.uk/wp-content/uploads/2016/03/EUCAST_E_Def_9_3_Mould_testing_definitive_0.pdf (accessed on 9 January 2023).
- Andreoli, T.E. The structure and function of amphotericin b-cholesterol pores in lipid bilayer membranes*. Ann. New York Acad. Sci. USA 1974, 235, 448–468. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Toshima, Y.; Minamikawa, H.; Hato, M.; Suzuki, K.; Kamo, N. Formation and Characterization of Planar Lipid Bilayer Membranes from Synthetic Phytanyl-Chained Glycolipids. Biochim. Et Biophys. Acta (BBA) Biomembr. 1999, 1421, 91–102. [Google Scholar] [CrossRef]
- Kara, S.; Afonin, S.; Babii, O.; Tkachenko, A.N.; Komarov, I.V.; Ulrich, A.S. Diphytanoyl Lipids as Model Systems for Studying Membrane-Active Peptides. Biochim. Et Biophys. Acta (BBA) Biomembr. 2017, 1859, 1828–1837. [Google Scholar] [CrossRef]
- Efimova, S.S.; Tevyashova, A.N.; Olsufyeva, E.N.; Bykov, E.E.; Ostroumova, O.S. Pore-Forming Activity of New Conjugate Antibiotics Based on Amphotericin B. PLoS ONE 2017, 12, e0188573. [Google Scholar] [CrossRef] [PubMed]
- Kleinberg, M.E.; Finkelstein, A. Single-Length and Double-Length Channels Formed by Nystatin in Lipid Bilayer Membranes. J. Membr. Biol. 1984, 80, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Chulkov, E.G.; Ostroumova, O.S. Phloretin Modulates the Rate of Channel Formation by Polyenes. Biochim. Et Biophys. Acta (BBA) Biomembr. 2016, 1858, 289–294. [Google Scholar] [CrossRef]
- Vandeputte, P.; Tronchin, G.; Larcher, G.; Ernoult, E.; Bergès, T.; Chabasse, D.; Bouchara, J.P. A Nonsense Mutation in the ERG6 Gene Leads to Reduced Susceptibility to Polyenes in a Clinical Isolate of Candida Glabrata. Antimicrob. Agents Chemother. 2008, 52, 3701–3709. [Google Scholar] [CrossRef]
- Hull, C.M.; Bader, O.; Parker, J.E.; Weig, M.; Gross, U.; Warrilow, A.G.; Kelly, D.E.; Kelly, S.L. Two Clinical Isolates of Candida Glabrata Exhibiting Reduced Sensitivity to Amphotericin B Both Harbor Mutations in ERG2. Antimicrob. Agents Chemother. 2012, 56, 6417–6421. [Google Scholar] [CrossRef]
- Jensen, R.H.; Astvad, K.M.T.; Silva, L.V.; Sanglard, D.; Jørgensen, R.; Nielsen, K.F.; Mathiasen, E.G.; Doroudian, G.; Perlin, D.S.; Arendrup, M.C. Stepwise Emergence of Azole, Echinocandin and Amphotericin B Multidrug Resistance in Vivo in Candida Albicans Orchestrated by Multiple Genetic Alterations. J. Antimicrob. Chemother. 2015, 70, 2551–2555. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.; Rolley, N.; Kelly, D.E.; Kelly, S.L. Identification and Characterization of Four Azole-Resistant Erg3 Mutants of Candida Albicans. Antimicrob. Agents Chemother. 2010, 54, 4527–4533. [Google Scholar] [CrossRef]
- Wilcock, B.C.; Endo, M.M.; Uno, B.E.; Burke, M.D. C2′-OH of Amphotericin B Plays an Important Role in Binding the Primary Sterol of Human Cells but Not Yeast Cells. J. Am. Chem. Soc. 2013, 135, 8488–8491. [Google Scholar] [CrossRef]
- Li, J.; Grillo, A.S.; Burke, M.D. From Synthesis to Function via Iterative Assembly of N-Methyliminodiacetic Acid Boronate Building Blocks. Acc. Chem. Res. 2015, 48, 2297–2307. [Google Scholar] [CrossRef]
- Boros-Majewska, J.; Salewska, N.; Borowski, E.; Milewski, S.; Malic, S.; Wei, X.-Q.; Hayes, A.J.; Wilson, M.J.; Williams, D.W. Novel Nystatin A1 Derivatives Exhibiting Low Host Cell Toxicity and Antifungal Activity in an in Vitro Model of Oral Candidosis. Med. Microbiol. Immunol. 2014, 203, 341–355. [Google Scholar] [CrossRef]
- Davis, S.A.; Vincent, B.M.; Endo, M.M.; Whitesell, L.; Marchillo, K.; Andes, D.R.; Lindquist, S.; Burke, M.D. Nontoxic Antimicrobials That Evade Drug Resistance. Nat. Chem. Biol. 2015, 11, 481–487. [Google Scholar] [CrossRef]
- Czub, J.; Borowski, E.; Baginski, M. Interactions of Amphotericin B Derivatives with Lipid Membranes—A Molecular Dynamics Study. Biochim. Et Biophys. Acta (BBA) Biomembr. 2007, 1768, 2616–2626. [Google Scholar] [CrossRef]
- Ostroumova, O.S.; Efimova, S.S.; Mikhailova, E.V.; Schagina, L.V. The Interaction of Dipole Modifiers with Amphotericin-Ergosterol Complexes. Effects of Phospholipid and Sphingolipid Membrane Composition. Eur. Biophys. J. 2014, 43, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Vincent, B.M.; Lancaster, A.K.; Scherz-Shouval, R.; Whitesell, L.; Lindquist, S. Fitness Trade-Offs Restrict the Evolution of Resistance to Amphotericin B. PLoS Biol. 2013, 11, e1001692. [Google Scholar] [CrossRef] [PubMed]
- Frías-De-León, M.G.; Hernández-Castro, R.; Vite-Garín, T.; Arenas, R.; Bonifaz, A.; Castañón-Olivares, L.; Acosta-Altamirano, G.; Martínez-Herrera, E. Antifungal Resistance in Candida Auris: Molecular Determinants. Antibiotics 2020, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Montal, M.; Mueller, P. Formation of Bimolecular Membranes from Lipid Monolayers and a Study of Their Electrical Properties. Proc. Natl. Acad. Sci. USA 1972, 69, 3561–3566. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minimum Inhibitory Concentration (MIC, µg/mL) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tested Strains | Compound | |||||||||||
| 1a 1 | 2a | 1b 1 | 2b | 1c | 2c | 1d | 2d | 1e | 2e | 1 | 2 | |
| C. parapsilosis ATCC 22019 | 0.03/0.06 2 | 0.5/1 | 0.06/0.125 | 2/4 | 1/1 | 1/2 | 2/2 | 2/4 | 1/2 | 1/2 | 1/2 | 2/2 |
| C. albicans ATCC 10231 | n/t | 1/2 | n/t | 2/4 | 1/1 | 2/4 | 2/2 | 2/4 | 1/1 | 4/8 | 0.5/0.5 | 1/2 |
| A. fumigatus ATCC 46645 | n/t | 2 | n/t | 4 | 1 | 2 | 4 | 2 | 2 | 8 | 2 | 4 |
| A. niger 37a | 0.125 | 8 | 0.5 | 16 | 4 | 8 | 1 | 32 | 2 | 16 | 2 | 8 |
| T. rubrum 2002 | 0.5 | 8 | 0.25 | 32 | 2 | 8 | 1 | 8 | 2 | 8 | 1 | 8 |
| C. krusei 432M | 0.25/0.5 | 4/4 | 0.25/0.5 | 8/8 | 2/2 | 16/16 | 2/2 | 8/8 | 2/2 | 8/16 | 0.5/1.0 | 4/4 |
| C. albicans 604M | 0.05/0.1 | 1/1 | n/t | 2/4 | 1/1 | 2/2 | 2/2 | 2/2 | 1/1 | 2/4 | 1/1 | 1/2 |
| C. albicans 8R | 0.05/0.1 | 1/2 | n/t | 4/4 | 1/1 | 2/4 | 2/2 | 4/4 | 1/1 | 2/4 | 1/1 | 1/2 |
| C. glabrata 61L | 0.03/0.06 | 1/1 | n/t | 2/2 | 1/1 | 1/2 | 2/4 | 2/2 | 2/2 | 2/2 | 1/2 | 1/2 |
| C. tropicalis 3010 | n/t | 1/1 | n/t | 2/2 | 0.5/1 | 2/2 | 2/2 | 2/2 | 1/1 | 2/4 | 0.5/1 | 1/2 |
| C. parapsilosis 58L | n/t | 1/2 | n/t | 2/4 | 1/1 | 1/2 | 2/2 | 2/4 | 1/1 | 2/2 | 0.5/1 | 1/2 |
| Compound | IC50, μM (HEK293) |
|---|---|
| 1a | 35.0 ± 4.0 |
| 1b | 4.0 ± 0.4 |
| 1c | 3.9 ± 0.5 |
| 1d | 20.0 ± 2.4 |
| 1e | 6.0 ± 0.7 |
| 2a | 47.0 ± 5.2 |
| 2b | 47.0 ± 4.7 |
| 2c | >50.0 |
| 2d | 15.0 ± 1.7 |
| 2e | >50.0 |
| 1 (AmB) | 18.0 ± 4.0 |
| 2 (Nys) | 15.0 ± 3.0 |
| Doxorubicin | 0.20 ± 0.02 |
| POPC/CHOL (67/33 mol %) | POPC/ERG (67/33 mol %) | EI 3 | |||||
|---|---|---|---|---|---|---|---|
| Polyene | RFmax 1, % | t1 2, min | t2 2, min | RFmax, % | t1, min | t2, min | |
| AmB (1) | 6 ± 1 | 0.6 ± 0.3 | 12.4 ± 8.1 | 9 ± 1 | 0.3 ± 0.1 | 5.8 ± 1.1 | 1.5 ± 0.4 |
| 1c | 13 ± 1 | 0.6 ± 0.1 | 7.9 ± 0.4 | 16 ± 2 | 0.5 ± 0.2 | 6.9 ± 1.1 | 1.2 ± 0.2 |
| 1d | 39 ± 3 | 0.5 ± 0.1 | 13.1 ± 4.8 | 57 ± 3 | 0.4 ± 0.1 | 11.9 ± 1.2 | 1.6 ± 0.2 |
| 1e | 16 ± 1 | 0.3 ± 0.1 | 4.7 ± 2.6 | 21 ± 3 | 0.1 ± 0.1 | 9.7 ± 2.4 | 1.3 ± 0.2 |
| Nys (2) | 10 ± 2 | 0.9 ± 0.3 | 5.7 ± 1.9 | 19 ± 3 | 0.7 ± 0.1 | 5.4 ± 0.5 | 1.9 ± 0.8 |
| 2a | 33 ± 5 | 0.5 ± 0.2 | 4.2 ± 0.2 | 44 ± 2 | 0.5 ± 0.1 | 4.8 ± 0.6 | 1.3 ± 0.3 |
| 2b | 71 ± 7 | 0.5 ± 0.1 | 4.6 ± 0.3 | 82 ± 5 | 0.5 ± 0.1 | 4.3 ± 0.4 | 1.3 ± 0.3 |
| 2c | 37 ± 9 | 0.2 ± 0.1 | 2.4 ± 0.2 | 86 ± 8 | 0.3 ± 0.1 | 6.6 ± 2.4 | 2.3 ± 0.8 |
| 2d | 39 ± 9 | 0.3 ± 0.1 | 3.9 ± 0.3 | 70 ± 6 | 0.5 ± 0.1 | 7.9 ± 2.4 | 1.8 ± 0.6 |
| 2e | 22 ± 4 | 0.9 ± 0.4 | 6.7 ± 0.4 | 75 ± 7 | 0.4 ± 0.1 | 2.9 ± 0.7 | 3.4 ± 0.9 |
| Polyene | G200 mV 1, pS | τ 2, ms | Pop 3 |
|---|---|---|---|
| 1 (AmB) 4 | 31.7 ± 2.1 | 23 ± 4 | 0.49 ± 0.09 |
| 1a4 | 20.8 ± 1.8 | 6 ± 2 | 0.23 ± 0.07 |
| 1c | 21.3 ± 3.2 | 10 ± 1 | 0.24 ± 0.11 |
| 1d | 21.2 ± 2.6 | 11 ± 1 | 0.14 ± 0.08 |
| 1e | 16.4 ± 2.2 | 17 ± 2 | 0.29 ± 0.12 |
| Polyene | CCHOL 1, 10−7 M | mCHOL 2 | CERG 1, 10−7 M | mERG 2 | CCHOL/CERG |
|---|---|---|---|---|---|
| AmB (1) | 0.98 ± 0.17 | 5–6 | 0.67 ± 0.16 | 6–6 | 1.5 ± 0.6 |
| 1c | 0.88 ± 0.08 | 5–5 | 0.98 ± 0.07 | 4–5 | 0.9 ± 0.1 |
| 1d | 0.94 ± 0.11 | 4–5 | 0.51 ± 0.09 | 5–6 | 1.8 ± 0.5 |
| 1e | 1.05 ± 0.05 | 4–5 | 0.92 ± 0.09 | 5–6 | 1.1 ± 0.2 |
| Nys (2) | 1.9 ± 0.2 | 2–3 | 1.8 ± 0.1 | 2–3 | 1.1 ± 0.2 |
| 2a | 2.2 ± 0.3 | 2–2 | 2.1 ± 0.6 | 2–2 | 1.0 ± 0.4 |
| 2b | 1.7 ± 0.1 | 3–3 | 1.6 ± 0.2 | 2–3 | 1.1 ± 0.2 |
| 2c | 3.1 ± 0.2 | 2–2 | 2.0 ± 0.4 | 2–3 | 1.6 ± 0.4 |
| 2d | 2.9 ± 0.5 | 2–3 | 2.7 ± 0.8 | 2–2 | 1.0 ± 0.5 |
| 2e | 2.4 ± 0.2 | 2–3 | 1.6 ± 0.3 | 2–3 | 1.5 ± 0.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tevyashova, A.; Efimova, S.; Alexandrov, A.; Omelchuk, O.; Ghazy, E.; Bychkova, E.; Zatonsky, G.; Grammatikova, N.; Dezhenkova, L.; Solovieva, S.; et al. Semisynthetic Amides of Amphotericin B and Nystatin A1: A Comparative Study of In Vitro Activity/Toxicity Ratio in Relation to Selectivity to Ergosterol Membranes. Antibiotics 2023, 12, 151. https://doi.org/10.3390/antibiotics12010151
Tevyashova A, Efimova S, Alexandrov A, Omelchuk O, Ghazy E, Bychkova E, Zatonsky G, Grammatikova N, Dezhenkova L, Solovieva S, et al. Semisynthetic Amides of Amphotericin B and Nystatin A1: A Comparative Study of In Vitro Activity/Toxicity Ratio in Relation to Selectivity to Ergosterol Membranes. Antibiotics. 2023; 12(1):151. https://doi.org/10.3390/antibiotics12010151
Chicago/Turabian StyleTevyashova, Anna, Svetlana Efimova, Alexander Alexandrov, Olga Omelchuk, Eslam Ghazy, Elena Bychkova, Georgy Zatonsky, Natalia Grammatikova, Lyubov Dezhenkova, Svetlana Solovieva, and et al. 2023. "Semisynthetic Amides of Amphotericin B and Nystatin A1: A Comparative Study of In Vitro Activity/Toxicity Ratio in Relation to Selectivity to Ergosterol Membranes" Antibiotics 12, no. 1: 151. https://doi.org/10.3390/antibiotics12010151
APA StyleTevyashova, A., Efimova, S., Alexandrov, A., Omelchuk, O., Ghazy, E., Bychkova, E., Zatonsky, G., Grammatikova, N., Dezhenkova, L., Solovieva, S., Ostroumova, O., & Shchekotikhin, A. (2023). Semisynthetic Amides of Amphotericin B and Nystatin A1: A Comparative Study of In Vitro Activity/Toxicity Ratio in Relation to Selectivity to Ergosterol Membranes. Antibiotics, 12(1), 151. https://doi.org/10.3390/antibiotics12010151