
Analysis of Extended Spectrum Beta Lactamase (ESBL) Genes of Non-Invasive ESBL Enterobacterales in Southeast Austria in 2017

 ,
,  and
and
Abstract
1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Collection of Isolates
5.2. Antibiotic Susceptibility Test
5.3. Strain Cultivation
5.4. MALDI-TOF
5.5. DNA Isolation
5.6. Determination of ESBL Genes
5.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adler, A.; Katz, D.E.; Marchaim, D. The Continuing Plague of Extended-Spectrum β-Lactamase Producing Enterbacterales Infections: An Update. Infect. Dis. Clin. N. Am. 2020, 34, 677–708. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Jacoby, G.A.; Medeiros, A.A. A Functional Classification Scheme for Beta-Lactamases and Its Correlation with Molecular Structure. Antimicrob. Agents Chemother. 1995, 39, 1211–1233. [Google Scholar] [CrossRef] [PubMed]
- Drawz, S.M.; Bonomo, R.A. Three Decades of Beta-Lactamase Inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Fourteen Years in Resistance. Int. J. Antimicrob. Agents 2012, 39, 283–294. [Google Scholar] [CrossRef]
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the Face of ESBLs in Europe. J. Antimicrob. Chemother. 2006, 59, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of Plasmids by PCR-Based Replicon Typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Woodford, N.; Carattoli, A.; Karisik, E.; Underwood, A.; Ellington, M.J.; Livermore, D.M. Complete Nucleotide Sequences of Plasmids PEK204, PEK499, and PEK516, Encoding CTX-M Enzymes in Three Major Escherichia Coli Lineages from the United Kingdom, All Belonging to the International O25:H4-ST131 Clone. Antimicrob. Agents Chemother. 2009, 53, 4472–4482. [Google Scholar] [CrossRef]
- Carattoli, A. Animal Reservoirs for Extended Spectrum Beta-Lactamase Producers. Clin. Microbiol. Infect. 2008, 14 (Suppl. S1), 117–123. [Google Scholar] [CrossRef]
- Garcia-Migura, L.; Hendriksen, R.S.; Fraile, L.; Aarestrup, F.M. Antimicrobial Resistance of Zoonotic and Commensal Bacteria in Europe: The Missing Link between Consumption and Resistance in Veterinary Medicine. Vet. Microbiol. 2014, 170, 1–9. [Google Scholar] [CrossRef]
- Hasman, H.; Mevius, D.; Veldman, K.; Olesen, I.; Aarestrup, F.M. β-Lactamases among Extended-Spectrum β-Lactamase (ESBL)-Resistant Salmonella from Poultry, Poultry Products and Human Patients in The Netherlands. J. Antimicrob. Chemother. 2005, 56, 115–121. [Google Scholar] [CrossRef]
- Hooban, B.; Joyce, A.; Fitzhenry, K.; Chique, C.; Morris, D. The Role of the Natural Aquatic Environment in the Dissemination of Extended Spectrum Beta-Lactamase and Carbapenemase Encoding Genes: A Scoping Review. Water Res. 2020, 180, 115880. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Silva, V.; Dapkevicius, M..d.L.E.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia Coli as Commensal and Pathogenic Bacteria Among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, E2239. [Google Scholar] [CrossRef] [PubMed]
- Barišić, I.; Mitteregger, D.; Hirschl, A.M.; Noehammer, C.; Wiesinger-Mayr, H. High Diversity of Beta-Lactamases in the General Hospital Vienna Verified by Whole Genome Sequencing and Statistical Analysis. Infect. Genet. Evol. 2014, 27, 408–417. [Google Scholar] [CrossRef]
- Franiek, N.; Orth, D.; Grif, K.; Ewers, C.; Wieler, L.H.; Thalhammer, J.G.; Würzner, R. ESBL-producing E. coli and EHEC in dogs and cats in the Tyrol as possible source of human infection. Berl. Munch. Tierarztl. Wochenschr. 2012, 125, 469–475. [Google Scholar] [PubMed]
- Huemer, H.P.; Eigentler, A.; Aschbacher, R.; Larcher, C. Dominance of CTX-M Group 1 Beta-Lactamase Enzymes in ESBL Producing E. Coli from Outpatient Urines in Neighboring Regions of Austria and Italy. Wien. Klin. Wochenschr. 2011, 123, 41–44. [Google Scholar] [CrossRef]
- Prelog, M.; Fille, M.; Prodinger, W.; Grif, K.; Brunner, A.; Würzner, R.; Zimmerhackl, L.B. CTX-M-1-Related Extended-Spectrum Beta-Lactamases Producing Escherichia Coli: So Far a Sporadic Event in Western Austria. Infection 2008, 36, 362–367. [Google Scholar] [CrossRef]
- Zarfel, G.; Galler, H.; Feierl, G.; Haas, D.; Kittinger, C.; Leitner, E.; Grisold, A.J.; Mascher, F.; Posch, J.; Pertschy, B.; et al. Comparison of Extended-Spectrum-β-Lactamase (ESBL) Carrying Escherichia Coli from Sewage Sludge and Human Urinary Tract Infection. Environ. Pollut. 2013, 173, 192–199. [Google Scholar] [CrossRef]
- Zarfel, G.; Lipp, M.; Gürtl, E.; Folli, B.; Baumert, R.; Kittinger, C. Troubled Water under the Bridge: Screening of River Mur Water Reveals Dominance of CTX-M Harboring Escherichia Coli and for the First Time an Environmental VIM-1 Producer in Austria. Sci. Total Environ. 2017, 593–594, 399–405. [Google Scholar] [CrossRef]
- Bujdáková, H.; Klimáková, J.; Allerberger, F.; Moravcíková, M.; Bagová, M.; Hanzen, J.; Michálková-Papajová, D.; Dierich, M.P.; Kettner, M. Spectrum and Transferability of Beta-Lactam Resistance in Hospital Strains of Enterobacter Isolated in Bratislava and Innsbruck. Int. J. Antimicrob. Agents 2000, 16, 31–36. [Google Scholar] [CrossRef]
- Eisner, A.; Fagan, E.J.; Feierl, G.; Kessler, H.H.; Marth, E.; Livermore, D.M.; Woodford, N. Emergence of Enterobacteriaceae Isolates Producing CTX-M Extended-Spectrum Beta-Lactamase in Austria. Antimicrob. Agents Chemother. 2006, 50, 785–787. [Google Scholar] [CrossRef] [PubMed]
- Apfalter, P.; Assadian, O.; Daxböck, F.; Hirschl, A.M.; Rotter, M.L.; Makristathis, A. Extended Double Disc Synergy Testing Reveals a Low Prevalence of Extended-Spectrum Beta-Lactamases in Enterobacter Spp. in Vienna, Austria. J. Antimicrob. Chemother. 2007, 59, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Loncaric, I.; Stalder, G.L.; Mehinagic, K.; Rosengarten, R.; Hoelzl, F.; Knauer, F.; Walzer, C. Comparison of ESBL--and AmpC Producing Enterobacteriaceae and Methicillin-Resistant Staphylococcus Aureus (MRSA) Isolated from Migratory and Resident Population of Rooks (Corvus Frugilegus) in Austria. PLoS ONE 2013, 8, e84048. [Google Scholar] [CrossRef] [PubMed]
- Desvars-Larrive, A.; Ruppitsch, W.; Lepuschitz, S.; Szostak, M.P.; Spergser, J.; Feßler, A.T.; Schwarz, S.; Monecke, S.; Ehricht, R.; Walzer, C.; et al. Urban Brown Rats (Rattus Norvegicus) as Possible Source of Multidrug-Resistant Enterobacteriaceae and Meticillin-Resistant Staphylococcus Spp., Vienna, Austria, 2016 and 2017. Euro Surveill. 2019, 24, 1900149. [Google Scholar] [CrossRef] [PubMed]
- Zarfel, G.; Galler, H.; Luxner, J.; Petternel, C.; Reinthaler, F.F.; Haas, D.; Kittinger, C.; Grisold, A.J.; Pless, P.; Feierl, G. Multiresistant Bacteria Isolated from Chicken Meat in Austria. Int. J. Environ. Res. Public Health 2014, 11, 12582–12593. [Google Scholar] [CrossRef] [PubMed]
- Eberhart, M.; Grisold, A.; Lavorato, M.; Resch, E.; Trobisch, A.; Resch, B. Extended-Spectrum Beta-Lactamase (ESBL) Producing Enterobacterales in Stool Surveillance Cultures of Preterm Infants Are No Risk Factor for Necrotizing Enterocolitis: A Retrospective Case-Control Study over 12 Years. Infection 2020, 48, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Bundesministeriums für Arbeit, Soziales, Gesundheit und Konsumentenschutz. Resistenzbericht Österreich AURES 2017; AURES Druckerei des Bundesministeriums für Arbeit, Soziales, Gesundheit und Konsumentenschutz: Wien, Austria, 2018; ISBN 978-3-85010-515-6. [Google Scholar]
- Rohde, A.M.; Zweigner, J.; Wiese-Posselt, M.; Schwab, F.; Behnke, M.; Kola, A.; Obermann, B.; Knobloch, J.K.-M.; Feihl, S.; Querbach, C.; et al. Incidence of Infections Due to Third Generation Cephalosporin-Resistant Enterobacteriaceae—A Prospective Multicentre Cohort Study in Six German University Hospitals. Antimicrob. Resist. Infect. Control 2018, 7, 159. [Google Scholar] [CrossRef]
- Fils, P.E.L.; Cholley, P.; Gbaguidi-Haore, H.; Hocquet, D.; Sauget, M.; Bertrand, X. ESBL-Producing Klebsiella Pneumoniae in a University Hospital: Molecular Features, Diffusion of Epidemic Clones and Evaluation of Cross-Transmission. PLoS ONE 2021, 16, e0247875. [Google Scholar] [CrossRef]
- Meini, S.; Tascini, C.; Cei, M.; Sozio, E.; Rossolini, G.M. AmpC β-Lactamase-Producing Enterobacterales: What a Clinician Should Know. Infection 2019, 47, 363–375. [Google Scholar] [CrossRef]
- Fiolić, Z.; Bosnjak, Z.; Bedenić, B.; Budimir, A.; Mareković, I.; Cetkovic, H.; Kalenić, S. Nationwide Survey of Klebsiella Pneumoniae Strains Producing CTX-M Extended-Spectrum β-Lactamases in Croatia. Coll. Antropol. 2015, 39, 947–951. [Google Scholar]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-Spectrum β-Lactamases: An Update on Their Characteristics, Epidemiology and Detection. JAC Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef]
- Giufrè, M.; Mazzolini, E.; Cerquetti, M.; Brusaferro, S. CCM2015 One-Health ESBL-producing Escherichia coli Study Group Extended-Spectrum β-Lactamase-Producing Escherichia Coli from Extraintestinal Infections in Humans and from Food-Producing Animals in Italy: A “One Health” Study. Int. J. Antimicrob. Agents 2021, 58, 106433. [Google Scholar] [CrossRef]
- Valentin, T.; Feierl, G.; Masoud-Landgraf, L.; Kohek, P.; Luxner, J.; Zarfel, G. Proteus Mirabilis Harboring Carbapenemase NDM-5 and ESBL VEB-6 Detected in Austria. Diagn. Microbiol. Infect. Dis. 2018, 91, 284–286. [Google Scholar] [CrossRef]
- Nicolas-Chanoine, M.-H.; Bertrand, X.; Madec, J.-Y. Escherichia Coli ST131, an Intriguing Clonal Group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef]
- Mazzariol, A.; Bazaj, A.; Cornaglia, G. Multi-Drug-Resistant Gram-Negative Bacteria Causing Urinary Tract Infections: A Review. J. Chemother. 2017, 29, 2–9. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Zummo, S.; Gerace, E.; Scappatura, G.; Biondo, C. Extended-Spectrum β-Lactamase & Carbapenemase-Producing Fermentative Gram-Negative Bacilli in Clinical Isolates from a University Hospital in Southern Italy. New Microbiol. 2021, 44, 227–233. [Google Scholar] [PubMed]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.; Shimoda, S.; Shimono, N. Current Epidemiology, Genetic Evolution and Clinical Impact of Extended-Spectrum β-Lactamase-Producing Escherichia Coli and Klebsiella Pneumoniae. Infect. Genet. Evol. 2018, 61, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Poulou, A.; Grivakou, E.; Vrioni, G.; Koumaki, V.; Pittaras, T.; Pournaras, S.; Tsakris, A. Modified CLSI Extended-Spectrum β-Lactamase (ESBL) Confirmatory Test for Phenotypic Detection of ESBLs among Enterobacteriaceae Producing Various β-Lactamases. J. Clin. Microbiol. 2014, 52, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 7.1. 2017. Available online: http://www.eucast.org (accessed on 31 October 2022).
- Stürenburg, E.; Kühn, A.; Mack, D.; Laufs, R. A Novel Extended-Spectrum Beta-Lactamase CTX-M-23 with a P167T Substitution in the Active-Site Omega Loop Associated with Ceftazidime Resistance. J. Antimicrob. Chemother. 2004, 54, 406–409. [Google Scholar] [CrossRef][Green Version]
- Asai, T.; Masani, K.; Sato, C.; Hiki, M.; Usui, M.; Baba, K.; Ozawa, M.; Harada, K.; Aoki, H.; Sawada, T. Phylogenetic Groups and Cephalosporin Resistance Genes of Escherichia Coli from Diseased Food-Producing Animals in Japan. Acta Vet. Scand. 2011, 53, 52. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, V.; Vijaykumar, G.S.; Sudeepa Kumar, M.; Prashanth, H.V.; Prakash, R.; Nagaraj, E.R. Phenotypic and Genotypic Methods for Detection of Extended Spectrum β Lactamase Producing Escherichia Coli and Klebsiella Pneumoniae Isolated from Ventilator Associated Pneumonia. J. Clin. Diagn. Res. 2013, 7, 1975–1978. [Google Scholar] [CrossRef] [PubMed]
- Vourli, S.; Giakkoupi, P.; Miriagou, V.; Tzelepi, E.; Vatopoulos, A.C.; Tzouvelekis, L.S. Novel GES/IBC Extended-Spectrum Beta-Lactamase Variants with Carbapenemase Activity in Clinical Enterobacteria. FEMS Microbiol. Lett. 2004, 234, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Naas, T.; Poirel, L.; Karim, A.; Nordmann, P. Molecular Characterization of In50, a Class 1 Integron Encoding the Gene for the Extended-Spectrum Beta-Lactamase VEB-1 in Pseudomonas Aeruginosa. FEMS Microbiol. Lett. 1999, 176, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Pasterán, F.; Rapoport, M.; Petroni, A.; Faccone, D.; Corso, A.; Galas, M.; Vázquez, M.; Procopio, A.; Tokumoto, M.; Cagnoni, V. Emergence of PER-2 and VEB-1a in Acinetobacter Baumannii Strains in the Americas. Antimicrob. Agents Chemother. 2006, 50, 3222–3224. [Google Scholar] [CrossRef] [PubMed][Green Version]



{kind=link}
{kind=link}
| Isolates/Body Site (%) | ||||||
|---|---|---|---|---|---|---|
| Species (n) | GUS | Feces | Wound | Skin | URT | LRT |
| E. coli (392) | 309 (78.8%) | 21 (5.4%) | 27 (6.9%) | 22 (5.6%) | 9 (2.3%) | 4 (1.02%) |
| K. pneumoniae (54) | 28 (51.9%) | 9 (16.4%) | 2 (3.7%) | 7 (13%) | 5 (9.26%) | 3 (5.56%) |
| P. mirabilis (8) | 3 (37.5%) | 1 (12.5%) | 3 (37.5%) | - | 1 (12.5%) | - |
| E. cloacae (5) | 1 (20%) | 2 (40%) | 1 (20%) | - | - | 1(20%) |
| K. oxytoca (5) | - | 5 (100%) | - | - | - | - |
| C. braakii (1) | 1 (100%) | - | - | - | - | - |
| C. freundii (1) | - | - | 1 (100%) | - | - | - |
| C. koseri (1) | 1 (100%) | - | - | - | - | - |
| Salmonella spp. * (1) | - | 1 (100%) | - | - | - | - |
| sum (468) | 343 (73.3%) | 39 (8.3%) | 34 (7.3%) | 29 (6.2%) | 15 (3.2%) | 8 (1.7%) |
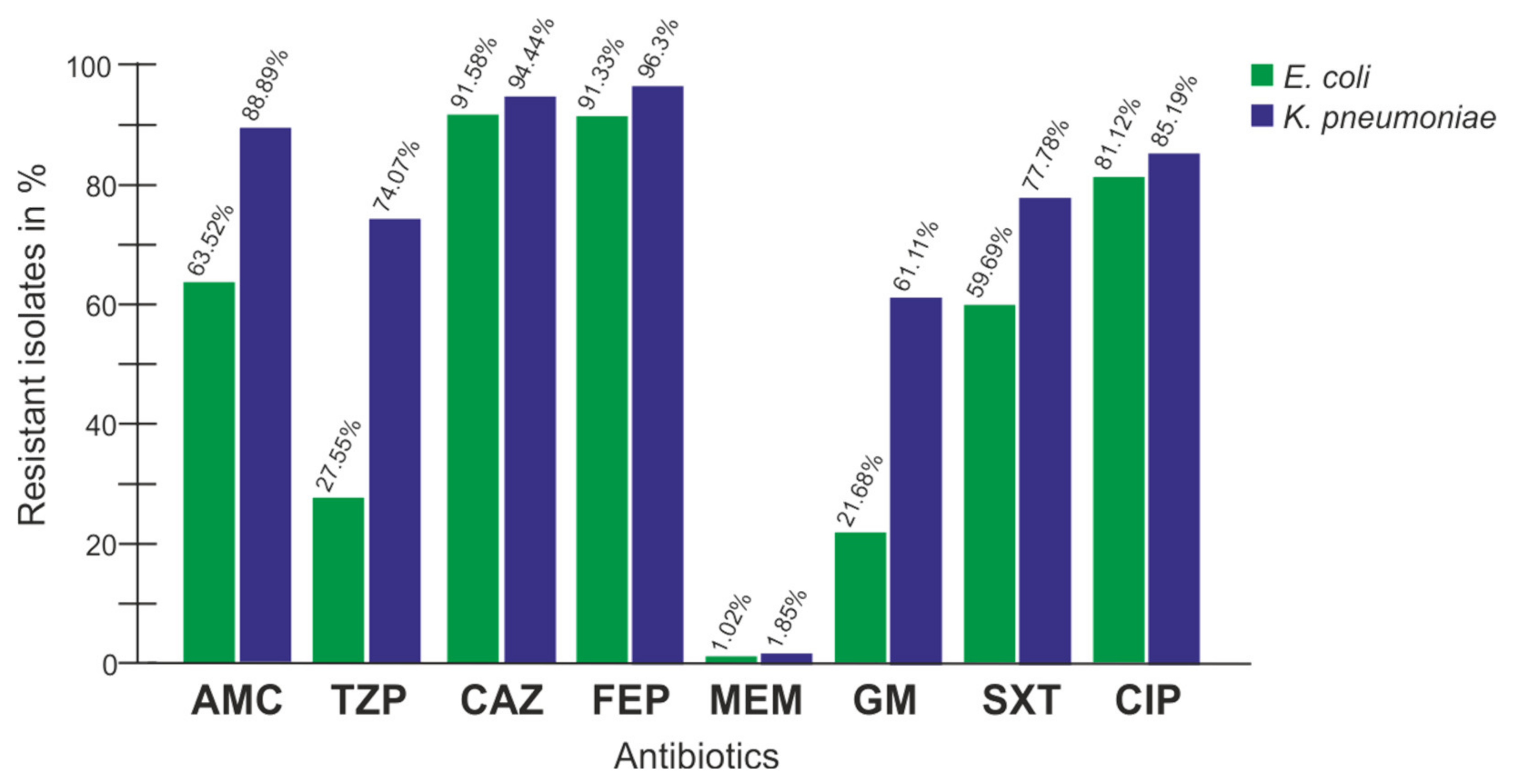
| AM | AMC | TZP | CXM | CTX | CAZ | FEP | MEM | GM | SXT | CIP | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli | 100 | 63.5 | 27.6 | 100 | 100 | 91.6 | 91.3 | 1.02 | 21.7 | 59.7 | 81.1 |
| K. pneumoniae | 100 | 88.9 | 74.1 | 100 | 100 | 94 | 96.3 | 1.85 | 61 | 77.8 | 85.19 |
| P. mirabilis | 100 | 37.5 | 12.5 | 100 | 100 | 87.5 | 100 | 12.5 | 62.5 | 75 | 87.5 |
| E. cloacae | 100 | 100 | 20 | 100 | 100 | 100 | 100 | 100 | 80 | 80 | 80 |
| K. oxytoca | 100 | 80 | 0 | 100 | 100 | 20 | 100 | 0 | 100 | 80 | 0 |
| C. braakii | 100 | 100 | 100 | 100 | 100 | 100 | 0 | 0 | 100 | 0 | 100 |
| C. freundii | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 0 | 100 | 0 | 0 |
| C. koseri | 100 | 0 | 0 | 100 | 100 | 100 | 100 | 0 | 0 | 100 | 0 |
| Salmonella spp. | 100 | 100 | 0 | 100 | 100 | 100 | 0 | 0 | 100 | 0 | 100 |
| sum | 100 | 66.7 | 32.5 | 100 | 100 | 91.2 | 91.9 | 1.28 | 28.9 | 62.2 | 80.6 |
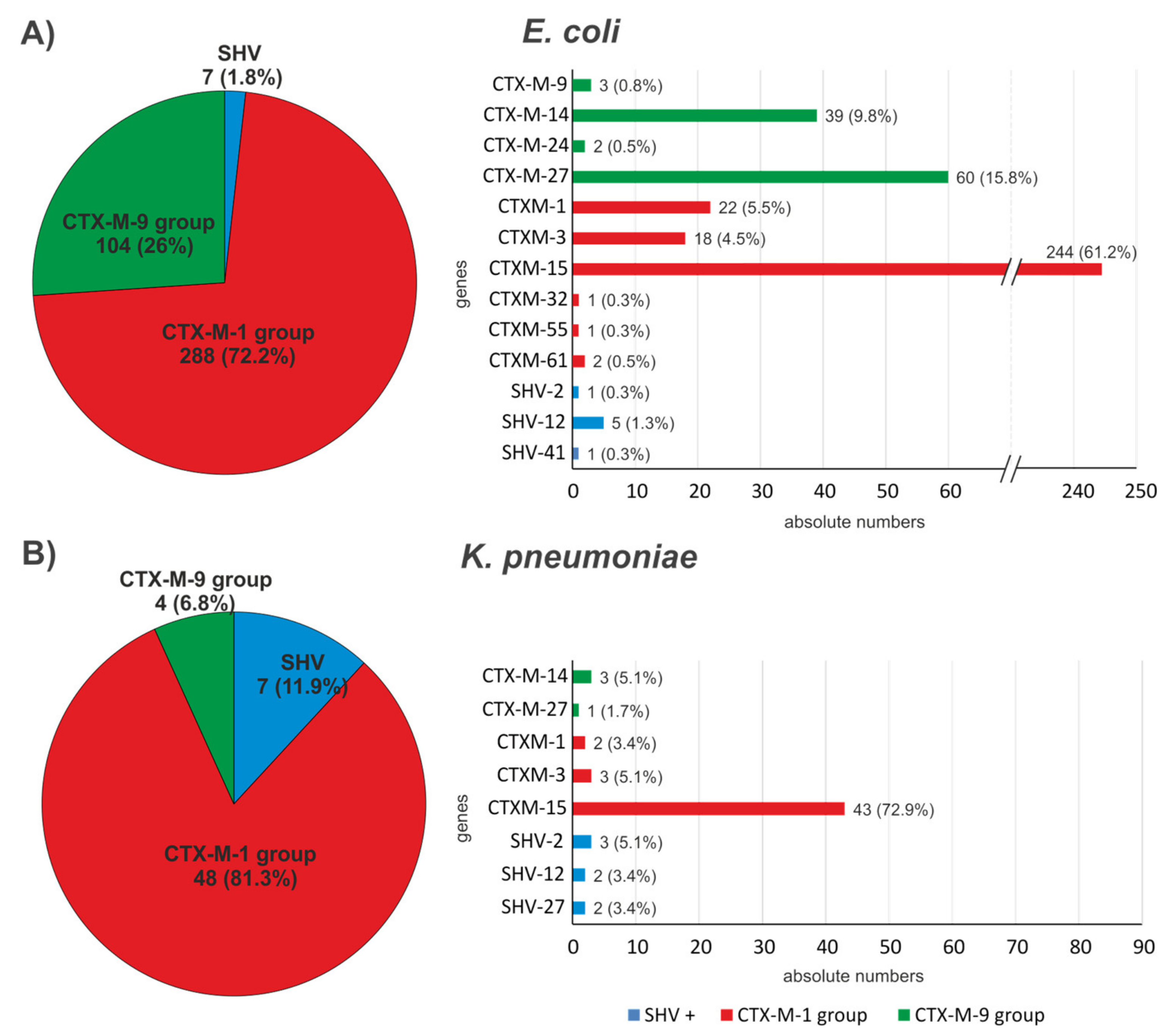
| Species | CTX-M-1 Group | CTX-M-2 Group | CTX-M-9 Group | SHV | VEB |
|---|---|---|---|---|---|
| E. coli * | blaCTX-M-1 (22) blaCTX-M-3 (18) blaCTX-M-15 (244) blaCTX-M-32 (1) blaCTX-M-55 (1) blaCTX-M-61 (2) | - | blaCTX-M-9 (3) blaCTX-M-14 (39) blaCTX-M-24 (2) blaCTX-M-27 (60) | blaSHV-2 (1) blaSHV-12 (5) blaSHV-41 (1) | - |
| K. pneumoniae ** | blaCTX-M-1 (2) blaCTX-M-3 (3) blaCTX-M-15 (43) | - | blaCTX-M-14 (3) blaCTX-M-27 (1) | blaSHV-2 (3) blaSHV-12 (2) blaSHV-27 (2) | - |
| P. mirabilis | blaCTX-M-15 (3) | blaCTX-M-2 (1) | blaCTX-M-27 (1) | - | blaVEB-6 (3) |
| E. cloacae *** | blaCTX-M-15 (4) | - | blaCTX-M-9 (1) | blaSHV-12 (1) | - |
| K. oxytoca | blaCTX-M-3 (5) | - | - | - | - |
| C. braakii | - | - | - | blaSHV-12 (1) | - |
| C. freundii | blaCTX-M-15 (1) | - | - | - | - |
| C. koseri | blaCTX-M-15 (1) | - | - | - | - |
| Salmonella spp. | - | - | - | blaSHV-12 (1) | - |
| Gene Families | Primer | Sequence 5’ → 3’ | Reference |
|---|---|---|---|
| blaCTX-M-1-group | f | TCTTCCAGAATAAGGAATCCC | [42] |
| r | CCGTTTCCGCTATTACAAAC | ||
| blaCTX-M-2-group | f | ATGATGACTCAGAGCATTCG | [43] |
| r | TGGGTTACGATTTTCGCCGC | ||
| blaCTX-M-9-group | f | ATGGTGACAAAGAGAGTGCA | [44] |
| r | CCCTTCGGCGATGATTCTC | ||
| blaGES | f | ATGCGCTTCATTCACGC C | [45] |
| r | CTATTTGTCCGTGCTCAGG | ||
| blaSHV | f | TGGTTATGCGTTATATTCGCC | [46] |
| r | GGTTAGCGTTGCCAGTGCT | ||
| blaTEM | f | TCCGCTCATGAGACAATAACC | [42] |
| r | TTGGTCTGACAGTTACCAATGC | ||
| blaVEB | f | GATAGGAGTACAGACATATG | [47] |
| r | TTTATTCAAATAGTAATTCCACG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulitsch-Fuchs, A.H.; Melchior, N.; Haitzmann, T.; Fingerhut, T.; Feierl, G.; Baumert, R.; Kittinger, C.; Zarfel, G. Analysis of Extended Spectrum Beta Lactamase (ESBL) Genes of Non-Invasive ESBL Enterobacterales in Southeast Austria in 2017. Antibiotics 2023, 12, 1. https://doi.org/10.3390/antibiotics12010001
Paulitsch-Fuchs AH, Melchior N, Haitzmann T, Fingerhut T, Feierl G, Baumert R, Kittinger C, Zarfel G. Analysis of Extended Spectrum Beta Lactamase (ESBL) Genes of Non-Invasive ESBL Enterobacterales in Southeast Austria in 2017. Antibiotics. 2023; 12(1):1. https://doi.org/10.3390/antibiotics12010001
Chicago/Turabian StylePaulitsch-Fuchs, Astrid H., Nadine Melchior, Theresa Haitzmann, Theres Fingerhut, Gebhard Feierl, Rita Baumert, Clemens Kittinger, and Gernot Zarfel. 2023. "Analysis of Extended Spectrum Beta Lactamase (ESBL) Genes of Non-Invasive ESBL Enterobacterales in Southeast Austria in 2017" Antibiotics 12, no. 1: 1. https://doi.org/10.3390/antibiotics12010001
APA StylePaulitsch-Fuchs, A. H., Melchior, N., Haitzmann, T., Fingerhut, T., Feierl, G., Baumert, R., Kittinger, C., & Zarfel, G. (2023). Analysis of Extended Spectrum Beta Lactamase (ESBL) Genes of Non-Invasive ESBL Enterobacterales in Southeast Austria in 2017. Antibiotics, 12(1), 1. https://doi.org/10.3390/antibiotics12010001