
Phage Cocktails with Daptomycin and Ampicillin Eradicates Biofilm-Embedded Multidrug-Resistant Enterococcus faecium with Preserved Phage Susceptibility

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolates
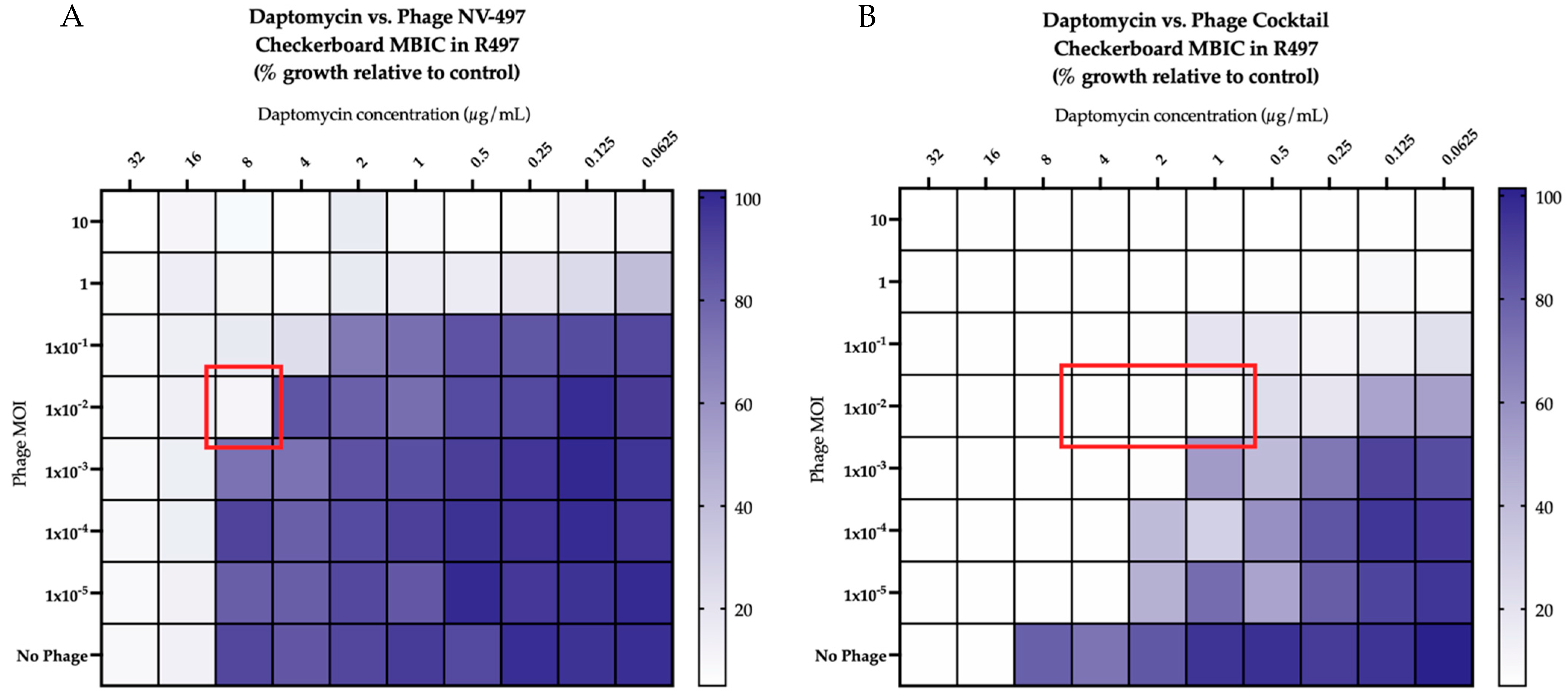
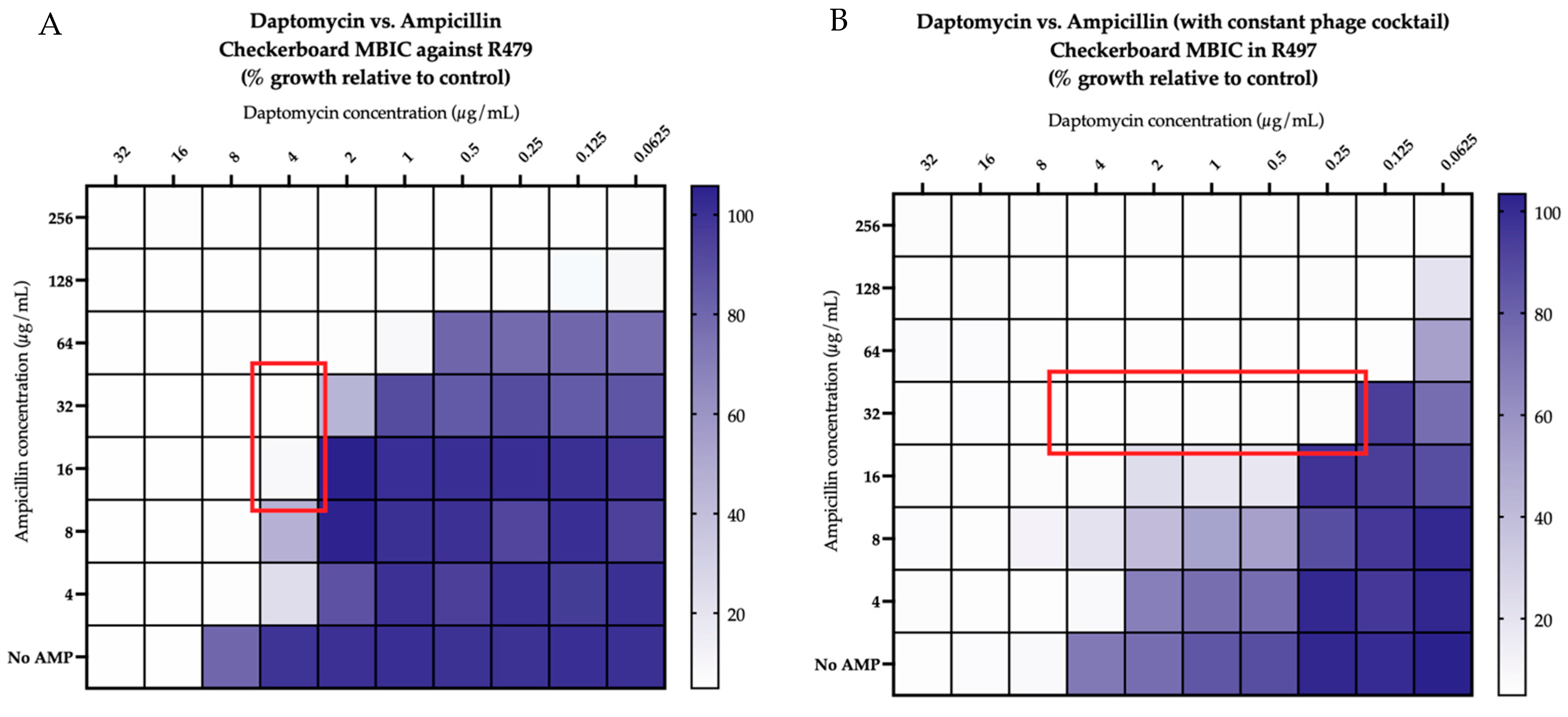
2.2. Checkerboard Analyses
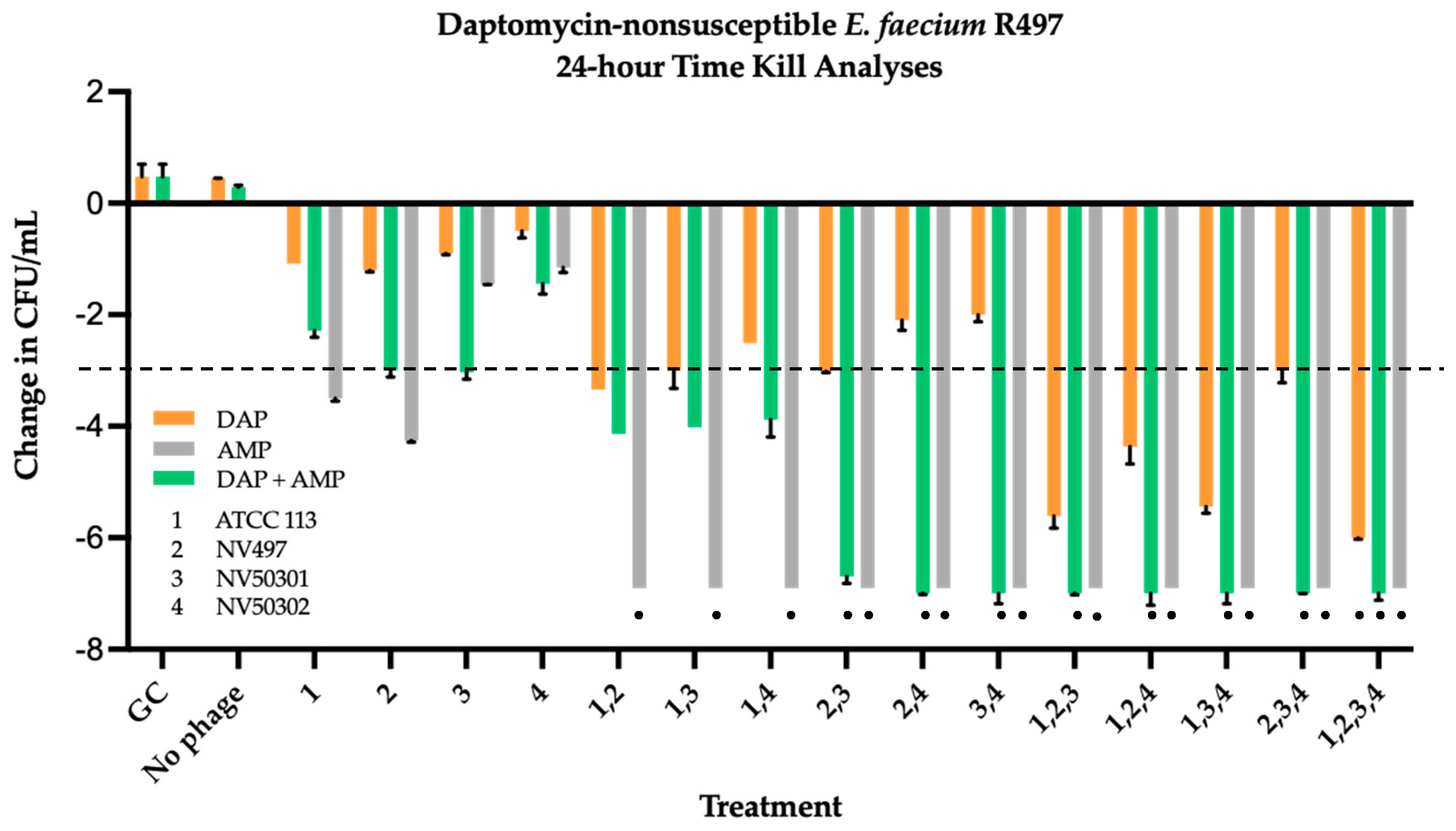
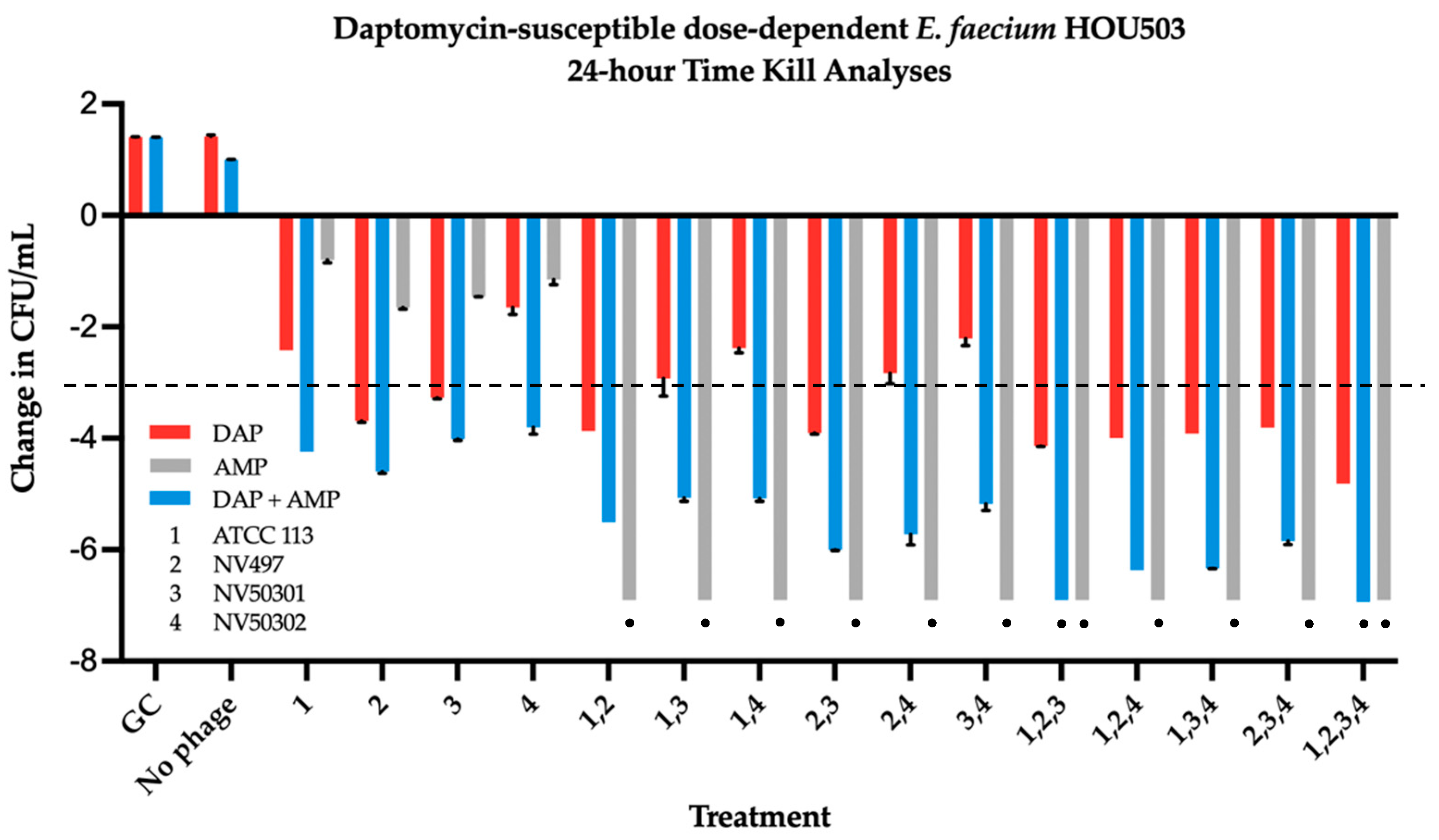
2.3. Time Kill Analyses
2.4. Bacteriophage and Antibiotic Resistance Testing
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Agents and Media
4.3. Bacteriophage Source and Propagation
4.4. Biofilm Quantification Assay
4.5. Phage Sensitivity Assay
4.6. Antibiotic Susceptibility Testing
4.7. Modified Checkerboard for Antibiotic and Bacteriophage Synergy Screening
4.8. Time Kill Analyses
4.9. Resistance Testing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Author Disclaimer
Support Statement
Copyright Statement
References
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Nigo, M.; Munita, J.M.; Arias, C.A.; Murray, B.E. What’s New in the Treatment of Enterococcal Endocarditis? Curr. Infect. Dis. Rep. 2014, 16, 431. [Google Scholar] [CrossRef]
- Chuang, Y.-C.; Lin, H.-Y.; Chen, P.-Y.; Lin, C.-Y.; Wang, J.-T.; Chen, Y.-C.; Chang, S.-C. Effect of Daptomycin Dose on the Outcome of Vancomycin-Resistant, Daptomycin-Susceptible Enterococcus faecium Bacteremia. Clin. Infect. Dis. 2017, 64, 1026–1034. [Google Scholar] [CrossRef]
- Zasowski, E.J.; Claeys, K.C.; Lagnf, A.M.; Davis, S.L.; Rybak, M.J. Time Is of the Essence: The Impact of Delayed Antibiotic Therapy on Patient Outcomes in Hospital-Onset Enterococcal Bloodstream Infections. Clin. Infect. Dis. 2016, 62, 1242–1250. [Google Scholar] [CrossRef]
- Cole, K.A.; Kenney, R.M.; Perri, M.B.; Dumkow, L.E.; Samuel, L.P.; Zervos, M.J.; Davis, S.L. Outcomes of Aminopenicillin Therapy for Vancomycin-Resistant Enterococcal Urinary Tract Infections. Antimicrob. Agents Chemother. 2015, 59, 7362–7366. [Google Scholar] [CrossRef]
- Lee, R.A.; Vo, D.T.; Zurko, J.C.; Griffin, R.L.; Rodriguez, J.M.; Camins, B.C. Infectious Diseases Consultation Is Associated with Decreased Mortality in Enterococcal Bloodstream Infections. Open Forum Infect. Dis. 2020, 7, ofaa064. [Google Scholar] [CrossRef]
- Higuita, N.I.; Huycke, M.M. Enterococcal Disease, Epidemiology, and Implications for Treatment. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection [Internet]; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Hidron, A.I.; Edwards, J.R.; Patel, J.; Horan, T.C.; Sievert, D.M.; Pollock, D.A.; Fridkin, S.K. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Annual Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef]
- Crank, C.; O’Driscoll, T. Vancomycin-resistant enterococcal infections: Epidemiology, clinical manifestations, and optimal management. Infect. Drug Resist. 2015, 8, 217. [Google Scholar] [CrossRef]
- Jahanbakhsh, S.; Singh, N.B.; Yim, J.; Kebriaei, R.; Smith, J.R.; Lev, K.; Tran, T.T.; Rose, W.E.; Arias, C.A.; Rybak, M.J. Impact of Daptomycin Dose Exposure Alone or in Combination with β-Lactams or Rifampin against Vancomycin-Resistant Enterococci in an In Vitro Biofilm Model. Antimicrob. Agents Chemother. 2020, 64, e02074-19. [Google Scholar] [CrossRef]
- Benamu, E.; Deresinski, S. Vancomycin-resistant enterococcus infection in the hematopoietic stem cell transplant recipient: An overview of epidemiology, management, and prevention. F1000Research 2018, 7, 3. [Google Scholar] [CrossRef]
- Stewart, P.S.; Davison, W.M.; Steenbergen, J.N. Daptomycin Rapidly Penetrates a Staphylococcus epidermidis Biofilm. Antimicrob. Agents Chemother. 2009, 53, 3505–3507. [Google Scholar] [CrossRef]
- Lellek, H.; Franke, G.C.; Ruckert, C.; Wolters, M.; Wolschke, C.; Christner, M.; Büttner, H.; Alawi, M.; Kröger, N.; Rohde, H. Emergence of daptomycin non-susceptibility in colonizing vancomycin-resistant Enterococcus faecium isolates during daptomycin therapy. Int. J. Med. Microbiol. 2015, 305, 902–909. [Google Scholar] [CrossRef]
- Kamboj, M.; Cohen, N.; Gilhuley, K.; Babady, N.E.; Seo, S.K.; Sepkowitz, K.A. Emergence of Daptomycin-Resistant VRE: Experience of a Single Institution. Infect. Control Hosp. Epidemiol. 2011, 32, 391–394. [Google Scholar] [CrossRef]
- Munita, J.M.; Murray, B.E.; Arias, C.A. Daptomycin for the treatment of bacteraemia due to vancomycin-resistant enterococci. Int. J. Antimicrob. Agents 2014, 44, 387–395. [Google Scholar] [CrossRef]
- El Haddad, L.; Hanson, B.M.; Arias, C.A.; Ghantoji, S.S.; Harb, C.P.; Stibich, M.; Chemaly, R.F. Emergence and Transmission of Daptomycin and Vancomycin-Resistant Enterococci Between Patients and Hospital Rooms. Clin. Infect. Dis. 2021, 73, 2306–2313. [Google Scholar] [CrossRef]
- Wudhikarn, K.; Gingrich, R.D.; de Magalhaes Silverman, M. Daptomycin nonsusceptible enterococci in hematologic malignancy and hematopoietic stem cell transplant patients: An emerging threat. Ann. Hematol. 2013, 92, 129–131. [Google Scholar] [CrossRef]
- Kebriaei, R.; Rice, S.A.; Singh, K.V.; Stamper, K.C.; Dinh, A.Q.; Rios, R.; Diaz, L.; Murray, B.E.; Munita, J.M.; Tran, T.T.; et al. Influence of Inoculum Effect on the Efficacy of Daptomycin Monotherapy and in Combination with β-Lactams against Daptomycin-Susceptible Enterococcus faecium Harboring LiaSR Substitutions. Antimicrob. Agents Chemother. 2018, 62, e00315-18. [Google Scholar] [CrossRef]
- Hall, A.D.; Steed, M.E.; Arias, C.A.; Murray, B.E.; Rybak, M.J. Evaluation of Standard- and High-Dose Daptomycin versus Linezolid against Vancomycin-Resistant Enterococcus Isolates in an In Vitro Pharmacokinetic/Pharmacodynamic Model with Simulated Endocardial Vegetations. Antimicrob. Agents Chemother. 2012, 56, 3174–3180. [Google Scholar] [CrossRef]
- Kebriaei, R.; Stamper, K.C.; Singh, K.V.; Khan, A.; Rice, S.A.; Dinh, A.Q.; Tran, T.T.; Murray, B.E.; Arias, C.A.; Rybak, M.J. Mechanistic Insights into the Differential Efficacy of Daptomycin Plus β-Lactam Combinations Against Daptomycin-Resistant Enterococcus faecium. J. Infect. Dis. 2020, 222, 1531–1539. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef]
- Morrisette, T.; Kebriaei, R.; Lev, K.L.; Morales, S.; Rybak, M.J. Bacteriophage Therapeutics: A Primer for Clinicians on Phage-Antibiotic Combinations. Pharmacotherapy 2020, 40, 153–168. [Google Scholar] [CrossRef]
- Tkhilaishvili, T.; Lombardi, L.; Klatt, A.-B.; Trampuz, A.; Luca, M.D. Bacteriophage Sb-1 enhances antibiotic activity against biofilm, degrades exopolysaccharide matrix and targets persisters of Staphylococcus aureus. Int. J. Antimicrob. Agents 2018, 52, 842–853. [Google Scholar] [CrossRef]
- Harper, D.R.; Parracho, H.M.R.T.; Walker, J.; Sharp, R.J.; Hughes, G.; Werthén, M.; Lehman, S.M.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270–284. [Google Scholar] [CrossRef]
- Parasion, S.; Kwiatek, M.; Gryko, R.; Mizak, L.; Malm, A. Bacteriophages as an alternative strategy for fighting biofilm development. Pol. J. Microbiol. 2014, 63, 137–145. [Google Scholar] [CrossRef]
- Kumaran, D.; Taha, M.; Yi, Q.; Ramirez-Arcos, S.; Diallo, J.-S.; Carli, A.V.; Abdelbary, H. Does Treatment Order Matter? Investigating the Ability of Bacteriophage to Augment Antibiotic Activity against Staphylococcus aureus Biofilms. Front. Microbiol. 2018, 9, 127. [Google Scholar] [CrossRef]
- Morrisette, T.; Lev, K.L.; Kebriaei, R.; Abdul-Mutakabbir, J.C.; Stamper, K.C.; Morales, S.; Lehman, S.M.; Canfield, G.S.; Duerkop, B.A.; Arias, C.A.; et al. Bacteriophage-Antibiotic Combinations for Enterococcus faecium with Varying Bacteriophage and Daptomycin Susceptibilities. Antimicrob. Agents Chemother. 2020, 64, e00993-20. [Google Scholar] [CrossRef]
- Torres-Barceló, C. Phage Therapy Faces Evolutionary Challenges. Viruses 2018, 10, 323. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef]
- Lev, K.; Kunz Coyne, A.J.; Kebriaei, R.; Morrisette, T.; Stamper, K.; Holger, D.J.; Canfield, G.S.; Duerkop, B.A.; Arias, C.A.; Rybak, M.J. Evaluation of Bacteriophage-Antibiotic Combination Therapy for Biofilm-Embedded MDR Enterococcus faecium. Antibiotics 2022, 11, 392. [Google Scholar] [CrossRef]
- Kebriaei, R.; Lev, K.; Morrisette, T.; Stamper, K.C.; Abdul-Mutakabbir, J.C.; Lehman, S.M.; Morales, S.; Rybak, M.J. Bacteriophage-Antibiotic Combination Strategy: An Alternative against Methicillin-Resistant Phenotypes of Staphylococcus aureus. Antimicrob. Agents Chemother. 2020, 64, e00461-20. [Google Scholar] [CrossRef]
- Tran, T.T.; Panesso, D.; Gao, H.; Roh, J.H.; Munita, J.M.; Reyes, J.; Diaz, L.; Lobos, E.A.; Shamoo, Y.; Mishra, N.N.; et al. Whole-Genome Analysis of a Daptomycin-Susceptible Enterococcus faecium Strain and Its Daptomycin-Resistant Variant Arising during Therapy. Antimicrob. Agents Chemother. 2012, 57, 261–268. [Google Scholar] [CrossRef]
- Diaz, L.; Tran, T.T.; Munita, J.M.; Miller, W.R.; Rincon, S.; Carvajal, L.P.; Wollam, A.; Reyes, J.; Panesso, D.; Rojas, N.L.; et al. Whole-Genome Analyses of Enterococcus faecium Isolates with Diverse Daptomycin MICs. Antimicrob. Agents Chemother. 2014, 58, 4527–4534. [Google Scholar] [CrossRef]
- Reyes, J.; Panesso, D.; Tran, T.T.; Mishra, N.N.; Cruz, M.R.; Munita, J.M.; Singh, K.V.; Yeaman, M.R.; Murray, B.E.; Shamoo, Y.; et al. A liaR Deletion Restores Susceptibility to Daptomycin and Antimicrobial Peptides in Multidrug-Resistant Enterococcus faecalis. J. Infect. Dis. 2015, 211, 1317–1325. [Google Scholar] [CrossRef]
- Munita, J.M.; Tran, T.T.; Diaz, L.; Panesso, D.; Reyes, J.; Murray, B.E.; Arias, C.A. A liaF Codon Deletion Abolishes Daptomycin Bactericidal Activity against Vancomycin-Resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 2013, 57, 2831–2833. [Google Scholar] [CrossRef]
- Bonilla, N.; Rojas, M.I.; Netto Flores Cruz, G.; Hung, S.-H.; Rohwer, F.; Barr, J.J. Phage on tap–a quick and efficient protocol for the preparation of bacteriophage laboratory stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef]
- Canfield, G.S.; Chatterjee, A.; Espinosa, J.; Mangalea, M.R.; Sheriff, E.K.; Keidan, M.; McBride, S.W.; McCollister, B.D.; Hang, H.C.; Duerkop, B.A. Lytic Bacteriophages Facilitate Antibiotic Sensitization of Enterococcus faecium. Antimicrob. Agents Chemother. 2021, 65, e00143-21. [Google Scholar] [CrossRef]
- Panesso, D.; Reyes, J.; Gaston, E.P.; Deal, M.; Londoño, A.; Nigo, M.; Munita, J.M.; Miller, W.R.; Shamoo, Y.; Tran, T.T.; et al. Deletion of liaR Reverses Daptomycin Resistance in Enterococcus faecium Independent of the Genetic Background. Antimicrob. Agents Chemother. 2015, 59, 7327–7334. [Google Scholar] [CrossRef] [Green Version]
- Grein, F.; Müller, A.; Scherer, K.M.; Liu, X.; Ludwig, K.C.; Klöckner, A.; Strach, M.; Sahl, H.-G.; Kubitscheck, U.; Schneider, T. Ca2+-Daptomycin targets cell wall biosynthesis by forming a tripartite complex with undecaprenyl-coupled intermediates and membrane lipids. Nat. Commun. 2020, 11, 1455. [Google Scholar] [CrossRef] [PubMed]
- Morrisette, T.; Lev, K.L.; Canfield, G.S.; Duerkop, B.A.; Kebriaei, R.; Stamper, K.C.; Holger, D.; Lehman, S.M.; Willcox, S.; Arias, C.A.; et al. Evaluation of Bacteriophage Cocktails Alone and in Combination with Daptomycin against Daptomycin-Nonsusceptible Enterococcus faecium. Antimicrob. Agents Chemother. 2022, 66, e0162321. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.M.; Mearns, G.; Rankin, D.; Cole, R.A.; Smrekar, F.; Branston, S.D.; Morales, S. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus aureus Infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.L.; Lehman, M.K.; Fey, P.D.; Bayles, K.W. Contribution of the Staphylococcus aureus Atl AM and GL Murein Hydrolase Activities in Cell Division, Autolysis, and Biofilm Formation. PLoS ONE 2012, 7, e42244. [Google Scholar] [CrossRef]
- Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of Bacteriophages Using the Small Drop Plaque Assay System. In Bacteriophages; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 501, pp. 81–85. [Google Scholar]
- Ceri, H.; Olson, M.; Morck, D.; Storey, D.; Read, R.; Buret, A.; Olson, B. The MBEC assay system: Multiple equivalent biofilms for antibiotic and biocide susceptibility testing. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2001; Volume 337, pp. 377–385. [Google Scholar]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef]
- Ali, L.; Khambaty, F.; Diachenko, G. Investigating the suitability of the Calgary Biofilm Device for assessing the antimicrobial efficacy of new agents. Bioresour. Technol. 2006, 97, 1887–1893. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically: M07-A10. In Approved Standard, 10th ed.; Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2015. [Google Scholar]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; on behalf of the CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B.; Carias, L.L.; Rudin, S.; Hutton, R.; Marshall, S.; Hassan, M.; Josseaume, N.; Dubost, L.; Marie, A.; Arthur, M. Role of Class A Penicillin-Binding Proteins in the Expression of β-Lactam Resistance in Enterococcus faecium. J. Bacteriol. 2009, 191, 3649–3656. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, R.A.; Leung, C.Y.; Chan, B.K.; Turner, P.E.; Weitz, J.S. Quantitative Models of Phage-Antibiotic Combination Therapy. mSystems 2020, 5, e00756-19. [Google Scholar] [CrossRef]
- Zhvania, P.; Hoyle, N.S.; Nadareishvili, L.; Nizharadze, D.; Kutateladze, M. Phage Therapy in a 16-Year-Old Boy with Netherton Syndrome. Front. Med. 2017, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Barber, K.E.; Werth, B.J.; McRoberts, J.P.; Rybak, M.J. A Novel Approach Utilizing Biofilm Time-Kill Curves To Assess the Bactericidal Activity of Ceftaroline Combinations against Biofilm-Producing Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 2989–2992. [Google Scholar] [CrossRef] [PubMed]
- Barber, K.E.; Shammout, Z.; Smith, J.R.; Kebriaei, R.; Morrisette, T.; Rybak, M.J. Biofilm Time-Kill Curves to Assess the Bactericidal Activity of Daptomycin Combinations against Biofilm-Producing Vancomycin-Resistant Enterococcus faecium and faecalis. Antibiotics 2021, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Rybak, M.J.; Hershberger, E.; Moldovan, T.; Grucz, R.G. In Vitro Activities of Daptomycin, Vancomycin, Linezolid, and Quinupristin-Dalfopristin against Staphylococci and Enterococci, Including Vancomycin- Intermediate and -Resistant Strains. Antimicrob. Agents Chemother. 2000, 44, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- O’Flynn, G.; Ross, R.P.; Fitzgerald, G.F.; Coffey, A. Evaluation of a Cocktail of Three Bacteriophages for Biocontrol of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2004, 70, 3417–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enterococcus Faecium Clinical Isolates | DAP | VAN | Biofilm Quantification | Phage Susceptibility | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ATCC Phage 113 | NV-497 | NV-503-01 | NV-503-02 | NV-S447-01 | NV-S447-02 | 9181 | 9183 | 9184 | ||||
| R497 | R | R | Medium | |||||||||
| HOU503 | SDD | R | High | |||||||||
| 5938 | R | R | Low | |||||||||
| S447 | SDD | R | Low | |||||||||
| S80849 | SDD | R | Low | |||||||||
| SF11499 | SDD | R | Low | |||||||||
| SF12047 | SDD | R | Low | |||||||||
| 12311 | SDD | R | Low | |||||||||
| Single Phage | 2-Phage Cocktails | 3-Phage Cocktails | 4-Phage Cocktail | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 1,2 | 1,3 | 1,4 | 2,3 | 2,4 | 3,4 | 1,2,3 | 1,2,4 | 1,3,4 | 2,3,4 | 1,2,3,4 | |
| R497 | |||||||||||||||
| DAP | |||||||||||||||
| AMP | |||||||||||||||
| DAP + AMP | |||||||||||||||
| HOU503 | |||||||||||||||
| DAP | |||||||||||||||
| AMP | |||||||||||||||
| DAP + AMP | |||||||||||||||
 = resistant
= resistant  = susceptible
= susceptiblePublisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunz Coyne, A.J.; Stamper, K.; Kebriaei, R.; Holger, D.J.; El Ghali, A.; Morrisette, T.; Biswas, B.; Wilson, M.; Deschenes, M.V.; Canfield, G.S.; et al. Phage Cocktails with Daptomycin and Ampicillin Eradicates Biofilm-Embedded Multidrug-Resistant Enterococcus faecium with Preserved Phage Susceptibility. Antibiotics 2022, 11, 1175. https://doi.org/10.3390/antibiotics11091175
Kunz Coyne AJ, Stamper K, Kebriaei R, Holger DJ, El Ghali A, Morrisette T, Biswas B, Wilson M, Deschenes MV, Canfield GS, et al. Phage Cocktails with Daptomycin and Ampicillin Eradicates Biofilm-Embedded Multidrug-Resistant Enterococcus faecium with Preserved Phage Susceptibility. Antibiotics. 2022; 11(9):1175. https://doi.org/10.3390/antibiotics11091175
Chicago/Turabian StyleKunz Coyne, Ashlan J., Kyle Stamper, Razieh Kebriaei, Dana J. Holger, Amer El Ghali, Taylor Morrisette, Biswajit Biswas, Melanie Wilson, Michael V. Deschenes, Gregory S. Canfield, and et al. 2022. "Phage Cocktails with Daptomycin and Ampicillin Eradicates Biofilm-Embedded Multidrug-Resistant Enterococcus faecium with Preserved Phage Susceptibility" Antibiotics 11, no. 9: 1175. https://doi.org/10.3390/antibiotics11091175
APA StyleKunz Coyne, A. J., Stamper, K., Kebriaei, R., Holger, D. J., El Ghali, A., Morrisette, T., Biswas, B., Wilson, M., Deschenes, M. V., Canfield, G. S., Duerkop, B. A., Arias, C. A., & Rybak, M. J. (2022). Phage Cocktails with Daptomycin and Ampicillin Eradicates Biofilm-Embedded Multidrug-Resistant Enterococcus faecium with Preserved Phage Susceptibility. Antibiotics, 11(9), 1175. https://doi.org/10.3390/antibiotics11091175