
Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing Enterobacterales in Healthy Community Dogs in Israel

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Characterization of Study Population and Prevalence of ESBL-PE Gut Colonization
2.2. Antibiotic Resistance Profiles of the ESBL-PE Isolates
2.3. Distribution of ESBL-PE Species and Whole Genome Sequencing Taxonomic Classification
2.4. ESBL-PE Resistome
2.5. Risk Factor Analysis for ESBL-PE Gut Colonization
3. Discussion
4. Materials and Methods
4.1. Study Design and Sampling Methods
4.2. Bacterial Isolation and Species Identification
4.3. Sample Size and Statistical Analysis
4.4. Whole Genome Sequencing
4.5. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sabtu, N.; Enoch, D.A.; Brown, N.M. Antibiotic Resistance: What, Why, Where, When and How? Br. Med. Bull. 2015, 116, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.A.; Hasan, F.; Ahmed, S.; Hameed, A. Extended-Spectrum β-Lactamases (ESBLs): Characterization, Epidemiology and Detection. Crit. Rev. Microbiol. 2004, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L. Resistance in Gram-Negative Bacteria: Enterobacteriaceae. Am. J. Med. 2006, 119, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Morales-López, S.; Yepes, J.A.; Prada-Herrera, J.C.; Torres-Jiménez, A. Enterobacteria in the 21st Century: A Review Focused on Taxonomic Changes. J. Infect. Dev. Ctries. 2019, 13, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Donskey, C.J. The Role of the Intestinal Tract as a Reservoir and Source for Transmission of Nosocomial Pathogens. Clin. Infect. Dis. 2004, 39, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Biehl, L.M.; Schmidt-Hieber, M.; Liss, B.; Cornely, O.A.; Vehreschild, M.J.G.T. Colonization and Infection with Extended Spectrum Beta-Lactamase Producing Enterobacteriaceae in High-Risk Patients–Review of the Literature from a Clinical Perspective. Crit. Rev. Microbiol. 2016, 42, 1–16. [Google Scholar] [CrossRef]
- Schwaber, M.J.; Carmeli, Y. Mortality and Delay in Effective Therapy Associated with Extended-Spectrum Beta-Lactamase Production in Enterobacteriaceae Bacteraemia: A Systematic Review and Meta-Analysis. J. Antimicrob. Chemother. 2007, 60, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Berge, A.C.B.; Atwill, E.R.; Sischo, W.M. Animal and Farm Influences on the Dynamics of Antibiotic Resistance in Faecal Escherichia Coli in Young Dairy Calves. Prev. Vet. Med. 2005, 69, 25–38. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.G.A.; Gonggrijp, M.A.; Hage, J.J.; Heuvelink, A.E.; Velthuis, A.; Lam, T.J.G.M.; van Schaik, G. Prevalence and Risk Factors for Extended-Spectrum β-Lactamase or AmpC-Producing Escherichia Coli in Organic Dairy Herds in the Netherlands. J. Dairy Sci. 2017, 100, 562–571. [Google Scholar] [CrossRef]
- Adler, A.; Sturlesi, N.; Fallach, N.; Zilberman-Barzilai, D.; Hussein, O.; Blum, S.E.; Klement, E.; Schwaber, M.J.; Carmeli, Y. Prevalence, Risk Factors, and Transmission Dynamics of Extended-Spectrum-β-Lactamase-Producing Enterobacteriaceae: A National Survey of Cattle Farms in Israel in 2013. J. Clin. Microbiol. 2015, 53, 3515–3521. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Poeta, P.; Sáenz, Y.; Vinué, L.; Rojo-Bezares, B.; Jouini, A.; Zarazaga, M.; Rodrigues, J.; Torres, C. Detection of Escherichia Coli Harbouring Extended-Spectrum β-Lactamases of the CTX-M, TEM and SHV Classes in Faecal Samples of Wild Animals in Portugal. J. Antimicrob. Chemother. 2006, 58, 1311–1312. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Semmler, T.; Stubbe, A.; Stubbe, M.; Wieler, L.H.; Schaufler, K. Chromosomally Encoded ESBL Genes in Escherichia Coli of ST38 from Mongolian Wild Birds. J. Antimicrob. Chemother. 2017, 72, 1310–1313. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Raza, S.; Schaufler, K.; Roschanski, N.; Sarwar, F.; Semmler, T.; Schierack, P.; Guenther, S. High Prevalence of CTX-M-15-Type ESBL-Producing E. Coli from Migratory Avian Species in Pakistan. Front. Microbiol. 2017, 8, 2476. [Google Scholar] [CrossRef]
- Baede, V.O.; Wagenaar, J.A.; Broens, E.M.; Duim, B.; Dohmen, W.; Nijsse, R.; Timmerman, A.J.; Hordijk, J. Longitudinal Study of Extended-Spectrum-β-Lactamase- and AmpC-Producing Enterobacteriaceae in Household Dogs. Antimicrob. Agents Chemother. 2015, 59, 3117–3124. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, J.; Schoormans, A.; Kwakernaak, M.; Duim, B.; Broens, E.; Dierikx, C.; Mevius, D.; Wagenaar, J.A. High Prevalence of Fecal Carriage of Extended Spectrum β-Lactamase/AmpC-Producing Enterobacteriaceae in Cats and Dogs. Front. Microbiol. 2013, 4, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, E.K.; Pearl, D.L.; Janecko, N.; Finley, R.L.; Reid-Smith, R.J.; Weese, J.S.; Peregrine, A.S. Risk Factors for Carriage of Antimicrobial-Resistant Salmonella Spp and Escherichia Coli in Pet Dogs from Volunteer Households in Ontario, Canada, in 2005 and 2006. Am. J. Vet. Res. 2015, 76, 959–968. [Google Scholar] [CrossRef]
- van den Bunt, G.; Fluit, A.C.; Spaninks, M.P.; Timmerman, A.J.; Geurts, Y.; Kant, A.; Scharringa, J.; Mevius, D.; Wagenaar, J.A.; Bonten, M.J.M.; et al. Faecal Carriage, Risk Factors, Acquisition and Persistence of ESBL-Producing Enterobacteriaceae in Dogs and Cats and Co-Carriage with Humans Belonging to the Same Household. J. Antimicrob. Chemother. 2020, 75, 342–350. [Google Scholar] [CrossRef]
- Toombs-Ruane, L.J.; Benschop, J.; French, N.P.; Biggs, P.J.; Midwinter, A.C.; Marshall, J.C.; Chan, M.; Drinković, D.; Fayaz, A.; Baker, M.G.; et al. Carriage of Extended-Spectrum-Beta-Lactamase- and AmpC Beta-Lactamase-Producing Escherichia Coli Strains from Humans and Pets in the Same Households. Appl. Environ. Microbiol. 2020, 86, e01613-20. [Google Scholar] [CrossRef]
- Schmidt, V.M.; Pinchbeck, G.L.; Nuttall, T.; McEwan, N.; Dawson, S.; Williams, N.J. Antimicrobial Resistance Risk Factors and Characterisation of Faecal E. Coli Isolated from Healthy Labrador Retrievers in the United Kingdom. Prev. Vet. Med. 2015, 119, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Salgado-Caxito, M.; Benavides, J.A.; Adell, A.D.; Paes, A.C.; Moreno-Switt, A.I. Global Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing-Escherichia Coli in Dogs and Cats–A Scoping Review and Meta-Analysis. One Health 2021, 12, 100236. [Google Scholar] [CrossRef]
- Abdel-Moein, K.A.; Samir, A. Occurrence of Extended Spectrum β-Lactamase-Producing Enterobacteriaceae among Pet Dogs and Cats: An Emerging Public Health Threat Outside Health Care Facilities. Am. J. Infect. Control. 2014, 42, 796–798. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Betts, J.; La Ragione, R.; van Essen-Zandbergen, A.; Ceccarelli, D.; Petinaki, E.; Koutinas, C.K.; Mevius, D.J. Occurrence and Characterization of Extended-Spectrum Cephalosporin-Resistant Enterobacteriaceae in Healthy Household Dogs in Greece. J. Med. Microbiol. 2018, 67, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Aslantaş, Ö.; Yilmaz, E.Ş. Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase (ESBL) and Plasmidic AmpC β-Lactamase (PAmpC) Producing Escherichia Coli in Dogs. J. Vet. Med. Sci 2017, 79, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shnaiderman-Torban, A.; Navon-Venezia, S.; Dor, Z.; Paitan, Y.; Arielly, H.; Ahmad, W.A.; Kelmer, G.; Fulde, M.; Steinman, A. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae Shedding in Farm Horses Versus Hospitalized Horses: Prevalence and Risk Factors. Animals 2020, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shnaiderman-Torban, A.; Steinman, A.; Meidan, G.; Paitan, Y.; Abu Ahmad, W.; Navon-Venezia, S. Petting Zoo Animals as an Emerging Reservoir of Extended-Spectrum β-Lactamase and AmpC-Producing Enterobacteriaceae. Front. Microbiol. 2019, 10, 2488. [Google Scholar] [CrossRef] [Green Version]
- Shnaiderman-Torban, A.; Navon-Venezia, S.; Kelmer, E.; Cohen, A.; Paitan, Y.; Arielly, H.; Steinman, A. Extended-Spectrum β-Lactamase-Producing Enterobacterales Shedding by Dogs and Cats Hospitalized in an Emergency and Critical Care Department of a Veterinary Teaching Hospital. Antibiotics 2020, 9, 545. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids. Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of Mobile Genetic Elements Associated with Antibiotic Resistance in Salmonella Enterica Using a Newly Developed Web Tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder – an Open Online Re-source for Identification of Antimicrobial Resistance Genes in next-Generation Sequencing Data and Predic-tion of Phenotypes from Genotypes. Microbial Genomics 2022, 8. [Google Scholar] [CrossRef]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The Link between Phylogeny and Virulence InEscherichia Coli Extraintestinal Infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Ruppé, É.; Woerther, P.-L.; Barbier, F. Mechanisms of Antimicrobial Resistance in Gram-Negative Bacilli. Ann. Intensive Care 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, K.S.; Gold, H.S.; Schwaber, M.J.; Venkataraman, L.; Qi, Y.; De Girolami, P.C.; Samore, M.H.; Anderson, G.; Rasheed, J.K.; Tenover, F.C. Variety of β-Lactamases Produced by Amoxicillin-Clavulanate-Resistant Escherichia Coli Isolated in the Northeastern United States. Antimicrob. Agents Chemother. 2004, 48, 1520–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Conza, J.A.; Badaracco, A.; Ayala, J.; Rodríguez, C.; Famiglietti, A.; Gutkind, G.O. β-Lactamases Produced by Amoxicillin-Clavulanate-Resistant Enterobacteria Isolated in Buenos Aires, Argentina: A New BlaTEM Gene. Rev. Argent Microbiol. 2014, 46, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global Phylogenetic Analysis of Escherichia Coli and Plasmids Carrying the Mcr-1 Gene Indicates Bacterial Diversity but Plasmid Restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.F.; Ali, M.M.M.; Mohamed, Z.K.; Moussa, T.A.; Klena, J.D. Fecal Carriage of Extended-Spectrum β-Lactamases and AmpC-Producing Escherichia Coli in a Libyan Community. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Day, M.J.; Rodríguez, I.; van Essen-Zandbergen, A.; Dierikx, C.; Kadlec, K.; Schink, A.-K.; Wu, G.; Chattaway, M.A.; DoNascimento, V.; Wain, J.; et al. Diversity of STs, Plasmids and ESBL Genes among Escherichia Coli from Humans, Animals and Food in Germany, the Netherlands and the UK. J. Antimicrob. Chemother. 2016, 71, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Hapuarachchi, I. Genetic Evaluation of ESBL E. Coli Urinary Isolates in Otago. Master’s Thesis, University of Otago, Dunedin, New Zealand, 2020. [Google Scholar]
- Kim, S.; Kim, H.; Kim, Y.; Kim, M.; Kwak, H.; Ryu, S. Whole-Genome Sequencing-Based Characteristics in Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli Isolated from Retail Meats in Korea. Microorganisms 2020, 8, 508. [Google Scholar] [CrossRef] [Green Version]
- Apostolakos, I.; Franz, E.; van Hoek, A.H.A.M.; Florijn, A.; Veenman, C.; Sloet-van Oldruitenborgh-Oosterbaan, M.M.; Dierikx, C.; van Duijkeren, E. Occurrence and Molecular Characteristics of ESBL/AmpC-Producing Escherichia Coli in Faecal Samples from Horses in an Equine Clinic. J. Antimicrob. Chemother. 2017, 72, 1915–1921. [Google Scholar] [CrossRef] [Green Version]
- Walther, B.; Lübke-Becker, A.; Stamm, I.; Gehlen, H.; Barton, A.K.; Janssen, T.; Wieler, L.H.; Guenther, S. Suspected Nosocomial Infections with Multi-Drug Resistant, E. Coli, Including Extended-Spectrum Beta-Lactamase (ESBL)-Producing Strains, in an Equine Clinic. Berl. Munch. Tierarztl. Wochenschr. 2014, 127, 421–427. [Google Scholar]
- Ewers, C.; Franiek, N.; Grif, K.; Orth, D.; Thalhammer, J.; Wieler, L.; Würzner, R. ESBL-produzierende E. coli und EHEC bei Hunden und Katzen in Tirol als mögliche Quelle für humane Infektionen. Berl. Munch. Tierarztl. Wochenschr. 2012, 125, 469–475. [Google Scholar]
- Wedley, A.L.; Maddox, T.W.; Westgarth, C.; Coyne, K.P.; Pinchbeck, G.L.; Williams, N.J.; Dawson, S. Prevalence of Antimicrobial-Resistant Escherichia Coli in Dogs in a Cross-Sectional, Community-Based Study. Vet. Rec. 2011, 168, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, C.; Wray, A. Salmonella in Domestic Animals; CABI: Wallingford, UK, 2000; ISBN 978-0-85199-916-6. [Google Scholar]
- Nijsse, R.; Mughini-Gras, L.; Wagenaar, J.A.; Ploeger, H.W. Coprophagy in Dogs Interferes in the Diagnosis of Parasitic Infections by Faecal Examination. Vet. Parasitol. 2014, 204, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Fahrion, A.S.; Schnyder, M.; Wichert, B.; Deplazes, P. Toxocara Eggs Shed by Dogs and Cats and Their Molecular and Morphometric Species-Specific Identification: Is the Finding of T. Cati Eggs Shed by Dogs of Epidemiological Relevance? Vet. Parasitol. 2011, 177, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Leite-Martins, L.R.; Mahú, M.I.M.; Costa, A.L.; Mendes, Â.; Lopes, E.; Mendonça, D.M.V.; Niza-Ribeiro, J.J.R.; de Matos, A.J.F.; da Costa, P.M. Prevalence of Antimicrobial Resistance in Enteric Escherichia Coli from Domestic Pets and Assessment of Associated Risk Markers Using a Generalized Linear Mixed Model. Prev. Vet. Med. 2014, 117, 28–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belas, A.; Salazar, A.S.; da Gama, L.T.; Couto, N.; Pomba, C. Risk Factors for Faecal Colonisation with Escherichia Coli Producing Extended-Spectrum and Plasmid-Mediated AmpC β-Lactamases in Dogs. Vet. Rec 2014, 175, 202. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the All-Bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-Lactamase Database (BLDB)-Structure and Function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef]
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 26 July 2022).
- The Comprehensive Antibiotic Resistance Database. Available online: https://card.mcmaster.ca/ (accessed on 26 July 2022).
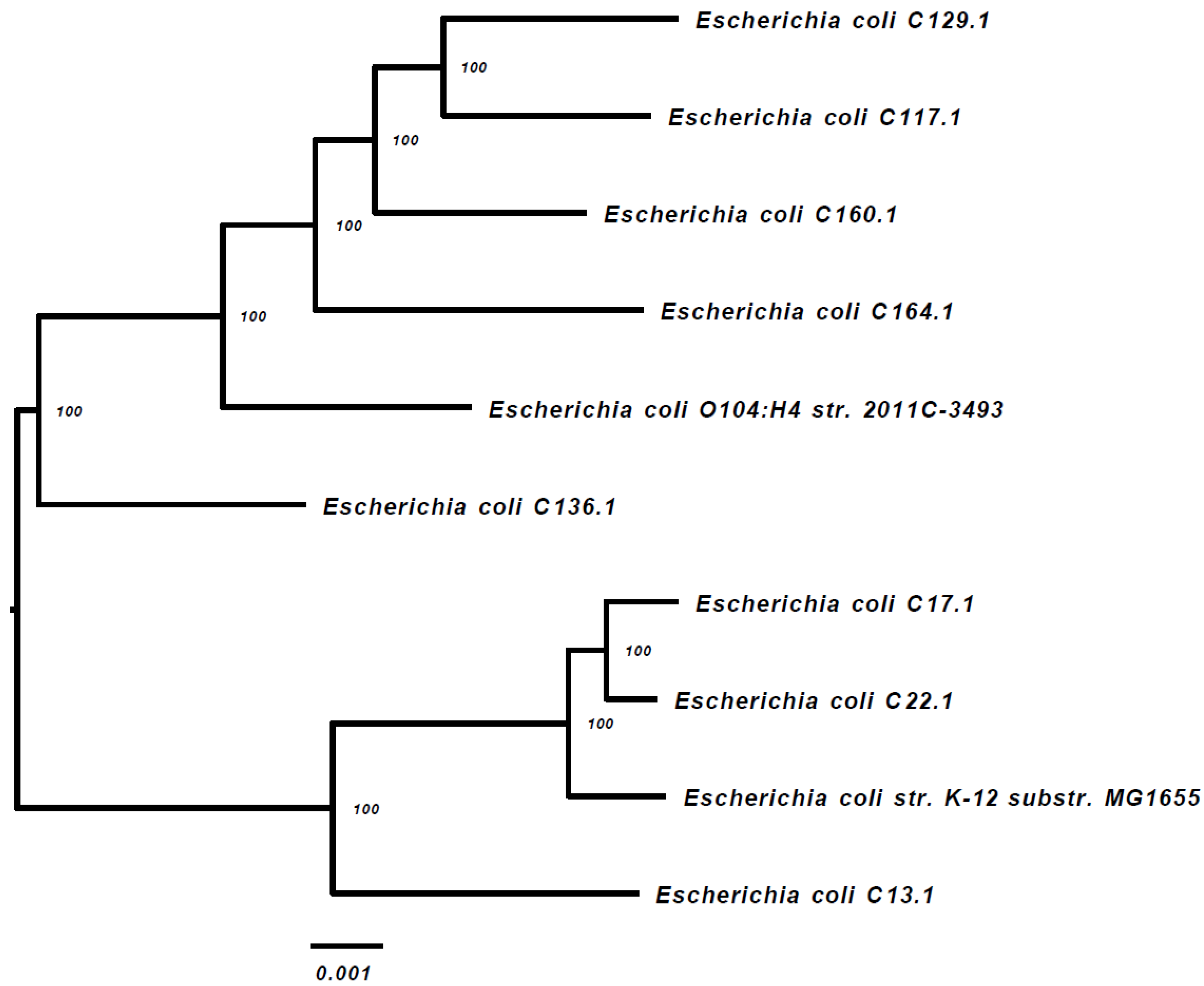

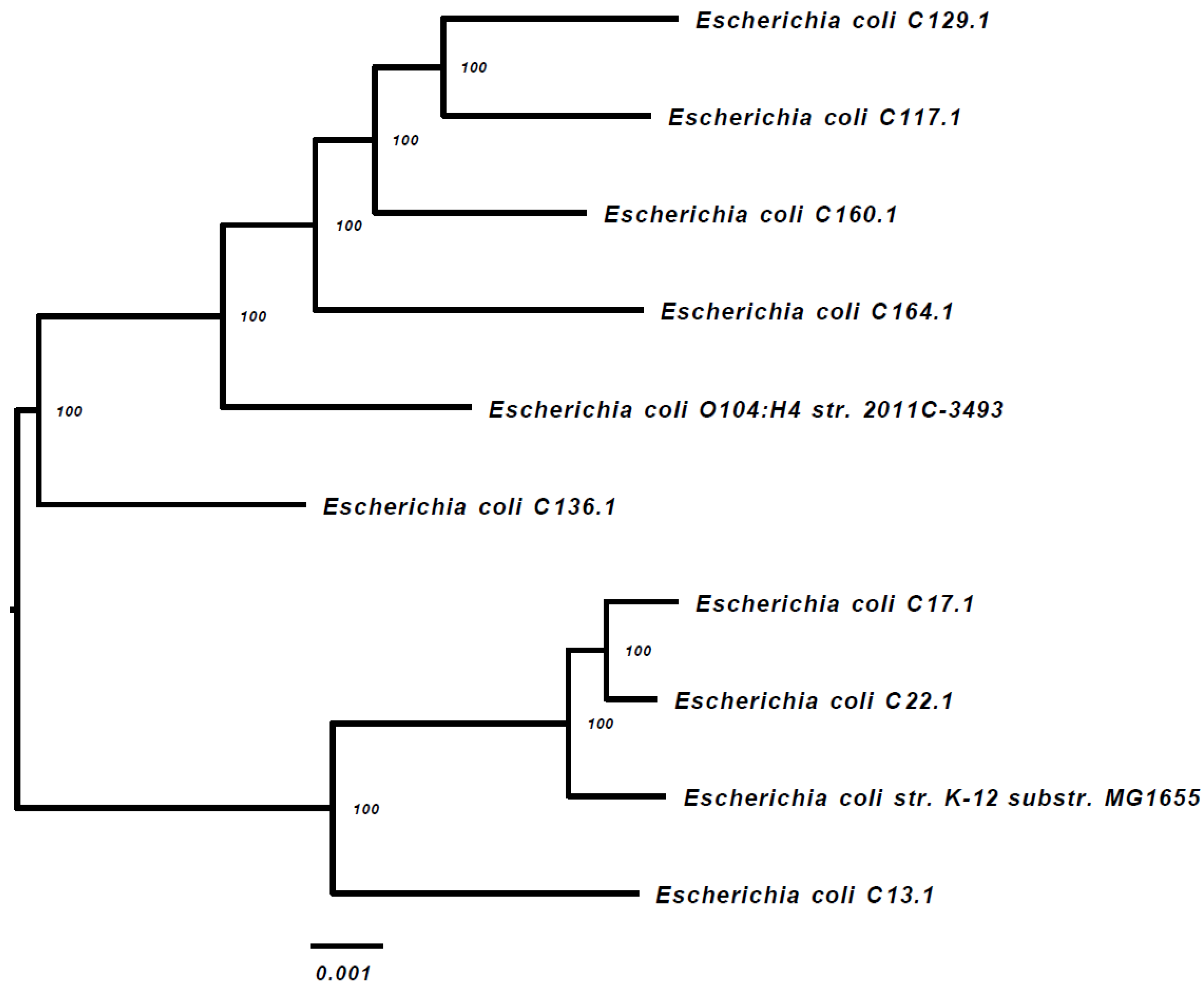{kind=link}
| ESBL-PE Isolate | Bacterial Species | E. coli Phylogenetic Group | Sequence Type | Antimicrobial Resistance 1,2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AMC | AMK | GENT | NIT | CIP | TMS | IMP | ||||
| C13.1 | E. coli | A | 46 | |||||||
| C17.1 | E. coli | A | 10 | |||||||
| C22.1 | E. coli | A | 10 | |||||||
| C23.2 | Citrobacter portucalensis | 165 | ||||||||
| C117.1 | E. coli | B1 | 10 | |||||||
| C129.1 | E. coli | B1 | 297 | |||||||
| C129.2 | Citrobacter braakii | 109 | ||||||||
| C136.1 | E. coli | C | 88 | |||||||
| C160.1 | E. coli | B1 | 602 | |||||||
| C164.1 | E. coli | B1 | 1586 | |||||||
| ESBL-P Isolate | Bla Genes | Quinolone | Tetracycline | Aminoglycoside | Macrolides | Sulfonamides | Trimethoprim | Chloramphenicol | Plasmid Replicon(s) | Plasmid/ Chromosome (Best Plasmid Homology by Blastn Search) | Notes |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C13.1 | PSE-1 CMY-171 TEM-156 | Tet(A) | AadA2, CmlA1, AadA1 | Mef(B) | Sul3 | IncI1, IncR, ColpVC, Col(MG828), IncFIA(HI1) | pMRSN346355_67.9 | ||||
| C17.1 | blaCTX-M-3 AmpC | IncFIB(AP001918, IncFII(pSE11), Col(MG828) | pZ0117KP0004-1 | Probably horizontal transfer from Klebsiella pneumoniae 1 | |||||||
| C22.1 | blaCTX-M-3 blaTEM-1 AmpC | QnrS1 | Tet(A), Tet(B) | APH (3″)-Ib, APH (6)-Id | Sul2 | DfrA14 | IncFII(pHN7A8), IncY | pCFSAN061768 | |||
| C23.2 | blaCMY-34 | QnrB9 | Chromosome | ||||||||
| C117.1 | CTX-M-55 blaTEM-234 blaEC | Tet(A) | AAC (3)-Iia, APH (3′)-Ia, APH (3″)-Ib, APH(6)-Id | Mef(B) | Sul2 | DfrA14 | CatII | IncFIB(AP001918), IncFIC(FII) | pTREC1 | ||
| C129.1 | blaCTX-M-100 TEM-234 blaTEM-156 | QnrVC1 | Tet(B), Tet(C) | AadA5, APH (3′)-Ia, AadA6 | Mph(E) | Sul1, Sul2 | DfrA17 | CatA1 | IncA/C2, IncY | pS44 related | Probably novel plasmid |
| C129.2 | blaCTX-M-39 blaTEM-237 blaCMY-93 | QnrVC1 | Tet(B), Tet(C) | AadA6, APH(3″)-Ib, APH(6)-Id, APH(3′)-Ia | Mph(E) | Sul1 | CatA1 | IncA/C2 | pS44 related | Probably novel plasmid | |
| C136.1 | SHV-12 AmpC | QnrB | Tet(A) | IncFIB(AP001918), IncFIC(FII), Col(MG828), IncX3, IncI1 | pEC-147 related | ||||||
| C160.1 | blaCTX-M-55 blaEC blaTEM-243 | Tet(A) | AadA1, APH (3″)-I, APH (6)-Id, Sul2 | Sul2 | DfrA1 | FloR | IncFIB(AP001918), IncFIC(FII), Col(MG828), IncN | Chromosome, plasmid pAH01-3-related | bla genes are located on a plasmid | ||
| C164.1 | blaCTX-M-3 blaTEM-237 blaEC-2199 | APH (3″), AAC (3)-Ii, APH(6)-Id | Mph(E) | Sul1 | DfrA12 | IncL/M, IncFIB(pHCM2) | pA1-3 | Probably horizontal transfer from Klebsiella pneumoniae 2 |
| Variable | Classification | No. (Valid%) | p-Value | OR (95% CI) |
|---|---|---|---|---|
| Age (years), median (range) | 2.6 (one month—12.5) | 0.179 | ||
| Sex | Female | 86 (63.2) | >0.999 | 0.851 (0.203–3.563) |
| Male | 50 (36.8) | |||
| Vaccination | Yes/No | 143 (99.3) | >0.999 | 0.993 (0.978–1.007) |
| Deworming | Yes/No | 115 (89.8) | 0.568 | 0.778 (0.088–6.87) |
| Ownership | Private household | 112 (77.2) | 0.21 | 0.757 (0.689–0.833) |
| Breeding farm | 33 (22.8) | |||
| Supervised walks by the owners | Yes/No | 132 (91.7) | 0.165 | 0.28 (0.051–1.53) |
| Commercial feeding only | Yes/No | 107 (73.8) | >0.999 | 1.26 (0.25–6.348) |
| Raw food | Yes/No | 9 (6.3) | >0.999 | 0.933 (0.891–0.976) |
| Hospitalization within the previous year | Yes/No | 20 (14.3) | >0.999 | 0.85 (0.099–7.301) |
| Antibiotic treatment within the previous year | Yes/No | 40 (29.2) | 0.416 | 1.885 (0.402–8.834) |
| Participation in shows | Yes/No | 5 (6.8) | >0.999 | 0.925 (0.865–0.99) |
| Coprophagia | Yes/No | 44 (30.5) | 0.024 * | 5.105 (1.215–21.457) |
| The presence of additional animals within the same household | Yes/No | 113 (78.5) | 0.405 | 0.523 (0.123–2.224) |
| Variable | p-Value | OR (95% CI) |
|---|---|---|
| Age | 0.149 | 0.762 (0.527–1.102) |
| Supervised walks | 0.137 | 0.243 (0.038–1.569) |
| Coprophagia | 0.048 * | 4.408 (1.014–19.169) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shnaiderman-Torban, A.; Navon-Venezia, S.; Baron, H.; Abu-Ahmad, W.; Arielly, H.; Zizelski Valenci, G.; Nissan, I.; Paitan, Y.; Steinman, A. Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing Enterobacterales in Healthy Community Dogs in Israel. Antibiotics 2022, 11, 1069. https://doi.org/10.3390/antibiotics11081069
Shnaiderman-Torban A, Navon-Venezia S, Baron H, Abu-Ahmad W, Arielly H, Zizelski Valenci G, Nissan I, Paitan Y, Steinman A. Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing Enterobacterales in Healthy Community Dogs in Israel. Antibiotics. 2022; 11(8):1069. https://doi.org/10.3390/antibiotics11081069
Chicago/Turabian StyleShnaiderman-Torban, Anat, Shiri Navon-Venezia, Hadar Baron, Wiessam Abu-Ahmad, Haya Arielly, Gal Zizelski Valenci, Israel Nissan, Yossi Paitan, and Amir Steinman. 2022. "Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing Enterobacterales in Healthy Community Dogs in Israel" Antibiotics 11, no. 8: 1069. https://doi.org/10.3390/antibiotics11081069
APA StyleShnaiderman-Torban, A., Navon-Venezia, S., Baron, H., Abu-Ahmad, W., Arielly, H., Zizelski Valenci, G., Nissan, I., Paitan, Y., & Steinman, A. (2022). Prevalence and Molecular Characterization of Extended-Spectrum β-Lactamase Producing Enterobacterales in Healthy Community Dogs in Israel. Antibiotics, 11(8), 1069. https://doi.org/10.3390/antibiotics11081069