
Synthetic Antimicrobial Immunomodulatory Peptides: Ongoing Studies and Clinical Trials

Abstract
:1. Introduction
2. Peptides Derived from Naturally Occurring Molecules
2.1. cNK-2
2.2. Bac2A
2.3. Clavanin-MO
2.4. Ac-ApoE (133–149)-NH2
3. De Novo Designed Antimicrobial Immunomodulatory Peptides
3.1. KSLW
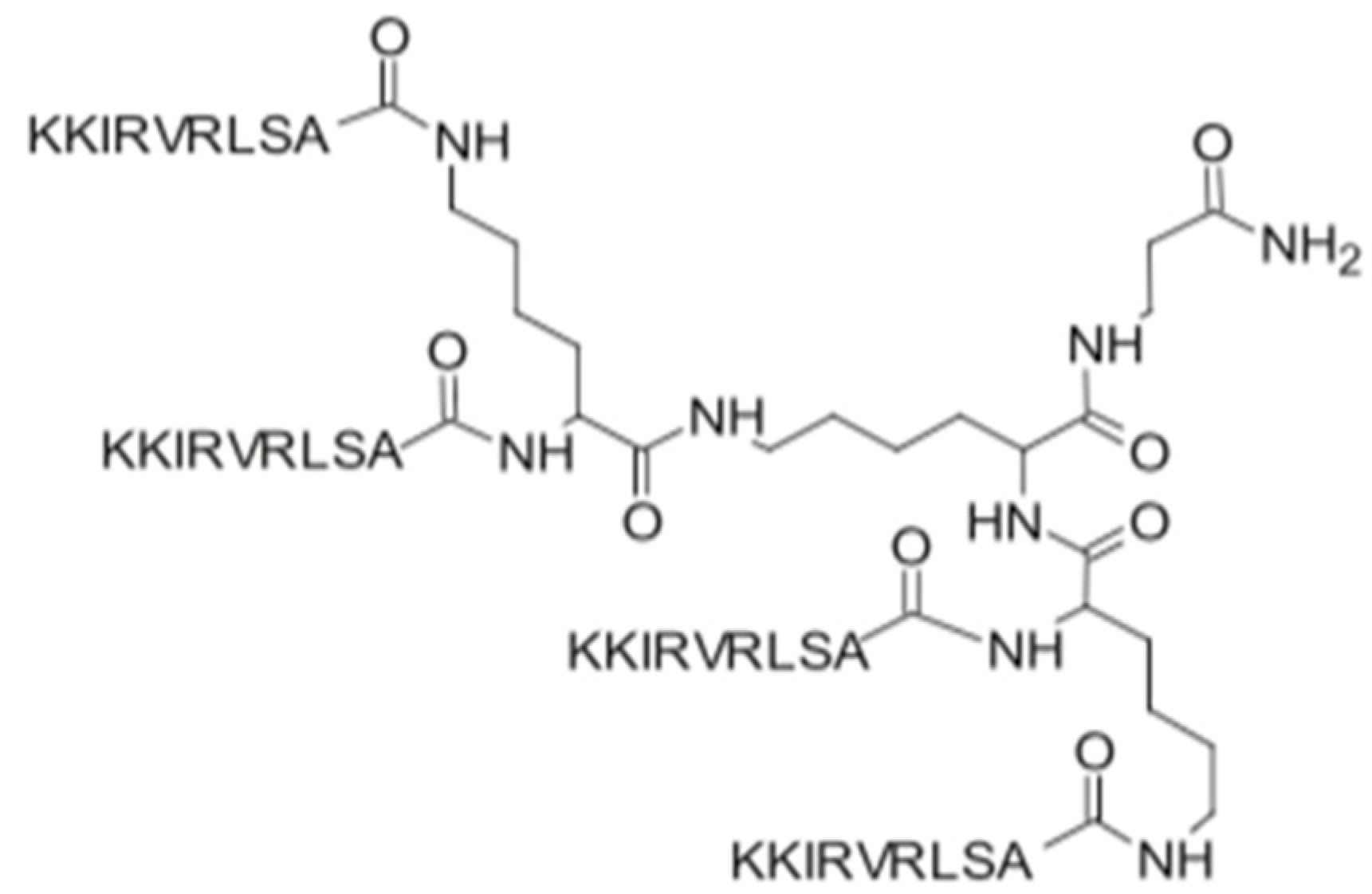
3.2. SET-M33
3.3. GW-A2
3.4. AWRK6
3.5. Synthetic Antimicrobial Immunomodulatory Peptides under Clinical Trials

{kind=link}
| De Novo Designed Synthetic Peptides | |||
|---|---|---|---|
| Peptide | Sequence | Activity | References |
| KSLW | KKVVFWVKFK-NH2 | Chemoattractant activity for neutrophils; induces actin polymerization in neutrophils; exerts an anti-inflammatory effect on phorbol myristate acetate (PMA)- and LPS-stimulated neutrophils. | [17] |
| mAb HuA VLCDR3 | GQTTVTKIDEDY | Induces a significant upregulation of IL-6 production. | [18] |
| mAb MoA VHCDR3 | GQYGNLWFAY | Induces an increased production of both IL-6 and TNF-α. | [18] |
| SET-M33 | KKIRVRLSA | Inhibition of the inflammatory cytokines TNF-α, IL1-β, MIP1, MIP2, IL-6, GM-CSF, KC, and IP10; decreases the expression of iNOS and COX-2; inhibitory effects on inflammatory cytokine expression in IB3–1 bronchial cells; in vivo neutralization of LPS; promotes keratinocyte migration. | [10] |
| GW-A2 | GAKYAKIIYNYLKKIANALW | Inhibition of the expression levels of nitric oxide (NO), inducible NO synthase (iNOS), cyclooxygenase-2 (COX-2), tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6) in lipopolysaccharide (LPS)-activated macrophages. | [11,12] |
| Dusquetide (SGX942) | RIVPA | Modulates the innate immune system at key convergence points in intracellular signaling pathways, and demonstrates activity in both reducing inflammation and increasing clearance of bacterial infection. | [19] |
| Pep19-2.5 Pep19-4LF | GCKKYRRFRWKFKGKFWFWG GKKYRRFRWKFKGKLFLFG | Anti-inflammatory activity of Pep19-2.5 is associated not only with neutralization of cell-free bacterial toxins, but also with a direct binding of the peptide to the outer leaflet of the bacterial outer membrane. | [20] |
| Synthetic Peptides Based on Naturally Occurring Molecules | |||
| Peptide | Sequence | Activity | References |
| cNK-2 | RRQRSICKQLLKKLRQQLSDALQNNDD | Upregulates the expression of CCL4, CCL5, and interleukin (IL)-1β in chicken macrophages; suppresses the expression of the pro-inflammatory cytokine IL-1β induced by LPS in HD11 cells and in primary monocytes; reduces the production of NO induced by LPS in HD11 cells and monocytes. | [4] |
| Bac2a | RLARIVVIRVAR-NH2 | Anti-endotoxic activity; induces chemotaxis of undifferentiated THP-1 cells. | [21] |
| clavanin-MO | FLPIIVFQFLGKIIHHVGNFVHGFSHVF-NH2 | Increases the production of IL-10 and reduces the expression of pro-inflammatory IL-12 and TNF-α; stimulates the migration of leukocytes; increases the expression of GM-CSF, IFN-γ, and MCP-1 in the early infection phase. | [6] |
| Ac-ApoE (133–149)-NH2 | Ac-LRVRLASHLRKLRKRLL-NH2 | Anti-inflammatory/neuroprotective activity and antimicrobial activity; binding of lipoprotein receptor-related protein (LRP) by apoE (133–149) results in inhibition of the N-methyl-D-aspartate (NMDA) receptor (NMDAR). | [7] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Review on Antimicrobial Resistance, chaired by Jim O’Neill; HM Government and Wellcome Trust: London, UK, 2016. [Google Scholar]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heuer, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.Y. Antimicrobial peptides: New hope in the war against multidrug resistance. Zool. Res. 2019, 40, 488–505. [Google Scholar] [CrossRef]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide cNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef] [Green Version]
- Bowdish, D.M.; Davidson, D.J.; Scott, M.G.; Hancock, R.E. Immunomodulatory activities of small host defense pep-tides. Antimicrob. Agents Chemother. 2005, 49, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Silva, O.N.; de la Fuente-Núñez, C.; Haney, E.F.; Fensterseifer, I.C.; Ribeiro, S.M.; Porto, W.F.; Brown, P.; Faria-Junior, C.; Rezende, T.M.; Moreno, S.E.; et al. An anti-infective synthetic peptide with dual antimicrobial and immunomodulatory activities. Sci. Rep. 2016, 6, 35465. [Google Scholar] [CrossRef] [Green Version]
- Pane, K.; Sgambati, V.; Zanfardino, A.; Smaldone, G.; Cafaro, V.; Angrisano, T.; Pedone, E.; Di Gaetano, S.; Capasso, D.; Haney, E.F.; et al. A new cryptic cationic antimicrobial peptide from human apolipoprotein E with antibacterial activity and immunomodulatory effects on human cells. FEBS J. 2016, 283, 2115–2131. [Google Scholar] [CrossRef] [Green Version]
- Na, D.H.; Faraj, J.; Capan, Y.; Leung, K.P.; DeLuca, P.P. Stability of antimicrobial decapeptide (KSL) and its analogues for delivery in the oral cavity. Pharm. Res. 2007, 24, 1544–1550. [Google Scholar] [CrossRef]
- Lee, W.; Park, E.J.; Min, G.; Choi, J.; Na, D.H.; Bae, J.S. Dual Functioned Pegylated Phospholipid Micelles Containing Cationic Antimicrobial Decapeptide for Treating Sepsis. Theranostics 2017, 7, 3759–3767. [Google Scholar] [CrossRef]
- Brunetti, J.; Roscia, G.; Lampronti, I.; Gambari, R.; Quercini, L.; Falciani, C.; Bracci, L.; Pini, A. Immunomodulatory and Anti-inflammatory Activity in Vitro and in Vivo of a Novel Antimicrobial Candidate. J. Biol. Chem. 2016, 291, 25742–25748. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.T.; Kuo, T.Y.; Chiang, J.C.; Pei, M.J.; Yang, W.T.; Yu, H.C.; Lin, S.B.; Chen, W.J. Design and synthesis of cationic antimicrobial peptides with improved activity and selectivity against Vibrio spp. Int. J. Antimicrob. Agents 2008, 32, 130–138. [Google Scholar] [CrossRef]
- Li, L.-H.; Ju, T.; Hsieh, C.-Y.; Dong, W.-C.; Chen, W.-T.; Hua, K.-F.; Chen, W.-J. A synthetic cationic antimicrobial peptide inhibits inflammatory response and the NLRP3 inflammasome by neutralizing LPS and ATP. PLoS ONE 2017, 12, e0182057. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Wang, Z.; Jin, L.; Li, H.; Hwang, J.; Hanrahan, J.W.; Wang, Q. Development of a Novel Antimicrobial Pep-tide AWRK6. Bangladesh J. Pharmacol. 2016, 11, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jin, L.; Wang, H.; Tai, S.; Liu, H.; Zhang, D. AWRK6, A Synthetic Cationic Peptide Derived from Antimicrobial Peptide Dybowskin-2CDYa, Inhibits Lipopolysaccharide-Induced Inflammatory Response. Int. J. Mol. Sci. 2018, 19, 600. [Google Scholar] [CrossRef] [Green Version]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Williams, R.L.; Sroussi, H.Y.; Leung, K.; Marucha, P.T. Antimicrobial decapeptide KSL-W enhances neutrophil chemotaxis and function. Peptides 2012, 33, 1–8. [Google Scholar] [CrossRef]
- Gabrielli, E.; Pericolini, E.; Cenci, E.; Ortelli, F.; Magliani, W.; Ciociola, T.; Bistoni, F.; Conti, S.; Vecchiarelli, A.; Polonelli, L. Antibody Complementarity-Determining Regions (CDRs): A Bridge between Adaptive and Innate Immunity. PLoS ONE 2009, 4, e8187. [Google Scholar] [CrossRef] [Green Version]
- Kudrimoti, M.; Curtis, A.; Azawi, S.; Worden, F.; Katz, S.; Adkins, D.; Bonomi, M.; Elder, J.; Sonis, S.T.; Straube, R.; et al. Dusquetide: A novel innate defense regulator demonstrating a significant and consistent reduction in the duration of oral mucositis in preclinical data and a randomized, placebo-controlled phase 2a clinical study. J. Biotechnol. 2016, 239, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Heinbockel, L.; Weindl, G.; Correa, W.; Brandenburg, J.; Reiling, N.; Wiesmüller, K.H.; Schürholz, T.; Gutsmann, T.; Martinez de Tejada, G.; Mauss, K.; et al. Anti-Infective and Anti-Inflammatory Mode of Action of Peptide 19-2.5. Int. J. Mol. Sci. 2021, 22, 1465. [Google Scholar] [CrossRef]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Greber, K.E.; Zielińska, J.; Nierzwicki, Ł.; Ciura, K.; Kawczak, P.; Nowakowska, J.; Bączek, T.; Sawicki, W. Are the short cationic lipopep-tides bacterial membrane disruptors? Structure-Activity Relationship and molecular dynamic evaluation. Biochim. Biophys. Acta Biomembr. 2019, 1861, 93–99. [Google Scholar] [CrossRef]
- Benfield, A.H.; Henriques, S.T. Mode-of-Action of Antimicrobial Peptides: Mem-brane Disruption vs. Intracellular Mechanisms. Front. Med. Technol. 2020, 2, 610997. [Google Scholar] [CrossRef]
- Fields, F.R.; Manzo, G.; Hind, C.; Janardhanan, J.; Foik, I.P.; Silva, P.D.C.; Balsara, R.D.; Clifford, M.; Vu, H.M.; Ross, J.N.; et al. Synthetic Antimicrobial Peptide Tuning Permits Membrane Disruption and Interpeptide Synergy. ACS Pharmacol. Transl. Sci. 2020, 3, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Farkas, A.; Maróti, G.; Kereszt, A.; Kondorosi, É. Comparative Analysis of the Bac-terial Membrane Disruption Effect of Two Natural Plant Antimicrobial Peptides. Front. Microbiol. 2017, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Greber, K.E.; Roch, M.; Rosato, M.A.; Martinez, M.P.; Rosato, A.E. Efficacy of newly generated short antimicrobial cationic lipopeptides against methicil-lin-resistant Staphylococcus aureus (MRSA). Int. J. Antimicrob. Agents 2020, 55, 105827. [Google Scholar] [CrossRef]
- Zharkova, M.S.; Orlov, D.S.; Golubeva, O.Y.; Chakchir, O.B.; Eliseev, I.E.; Grinchuk, T.M.; Shamova, O.V. Application of Antimicrobial Peptides of the Innate Immune System in Combination with Conventional Antibi-otics-A Novel Way to Combat Antibiotic Re-sistance. Front. Cell. Infect. Microbiol. 2019, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Haney, E.F.; Hancock, R.E. Peptide design for antimicrobial and immuno-modulatory ap-plications. Biopolymers 2013, 100, 572–583. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E. Immune modulation by multifaceted cat-ionic host defense (antimicrobial) peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef]
- Drayton, M.; Deisinger, J.P.; Ludwig, K.C.; Raheem, N.; Müller, A.; Schneider, T.; Straus, S.K. Host Defense Peptides: Dual Anti-microbial and Immunomodulatory Action. Int. J. Mol. Sci. 2021, 22, 11172. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.; Middelkoop, E.; Boekema, B. Review: Lessons Learned from Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef]
| Microorganism | MIC (μM) | |
|---|---|---|
| Clavanin A | Clavanin-MO | |
| Bacillus subtilis ATCC6633 | 6.0 | 1.5 |
| Enterococcus faecalis ATCC12953 | 6.0 | 1.5 |
| Staphylococcus aureus ATCC29213 | 48.0 | 24.0 |
| Staphylococcus aureus (MRSA) ATCC33591 | 12.0 | 6.0 |
| Escherichia coli ATCC8739 | 24.0 | 12.0 |
| Escherichia coli KPC-positive (1812446) | 48.0 | 6.0 |
| Klebsiella pneumoniae ATCC13885 | 6.0 | 3.0 |
| Klebsiella pneumoniae 1825971 (KPC971) | 6.0 | 3.0 |
| Pseudomonas aeruginosa ATCC 15442 | 12.0 | 3.0 |
| Microorganism | MIC (μM) | |||
|---|---|---|---|---|
| rApoE PM (133–150) | sApoE (133–150) | Ac-ApoE (133–150)-NH2 | Cog-133 | |
| Escherichia coli DH5α | 12.5 | 12.5 | 12.5 | 12.5 |
| E. coli ATCC 25922 | 6.25 | 6.25 | 6.25 | 6.25 |
| P. aeruginosa PAO1 | 25 | 25 | 25 | 25 |
| P. aeruginosa PA14 | 50 | 50 | 50 | 50 |
| K. pneumoniae ATCC 700603 | 6.25 | 6.25 | 3.12 | 3.12 |
| S. aureus ATCC 6538P | 3.12 | 3.12 | 100 | 100 |
| B. subtilis PY79 | 25 | 25 | >100 | >100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesiuk, M.; Paduszyńska, M.; Greber, K.E. Synthetic Antimicrobial Immunomodulatory Peptides: Ongoing Studies and Clinical Trials. Antibiotics 2022, 11, 1062. https://doi.org/10.3390/antibiotics11081062
Lesiuk M, Paduszyńska M, Greber KE. Synthetic Antimicrobial Immunomodulatory Peptides: Ongoing Studies and Clinical Trials. Antibiotics. 2022; 11(8):1062. https://doi.org/10.3390/antibiotics11081062
Chicago/Turabian StyleLesiuk, Małgorzata, Małgorzata Paduszyńska, and Katarzyna E. Greber. 2022. "Synthetic Antimicrobial Immunomodulatory Peptides: Ongoing Studies and Clinical Trials" Antibiotics 11, no. 8: 1062. https://doi.org/10.3390/antibiotics11081062
APA StyleLesiuk, M., Paduszyńska, M., & Greber, K. E. (2022). Synthetic Antimicrobial Immunomodulatory Peptides: Ongoing Studies and Clinical Trials. Antibiotics, 11(8), 1062. https://doi.org/10.3390/antibiotics11081062