
The Monitoring of Mycoplasma gallisepticum Minimum Inhibitory Concentrations during the Last Decade (2010–2020) Seems to Reveal a Comeback of Susceptibility to Macrolides, Tiamulin, and Lincomycin

 ,
,  ,
,  and
and
Abstract
1. Introduction
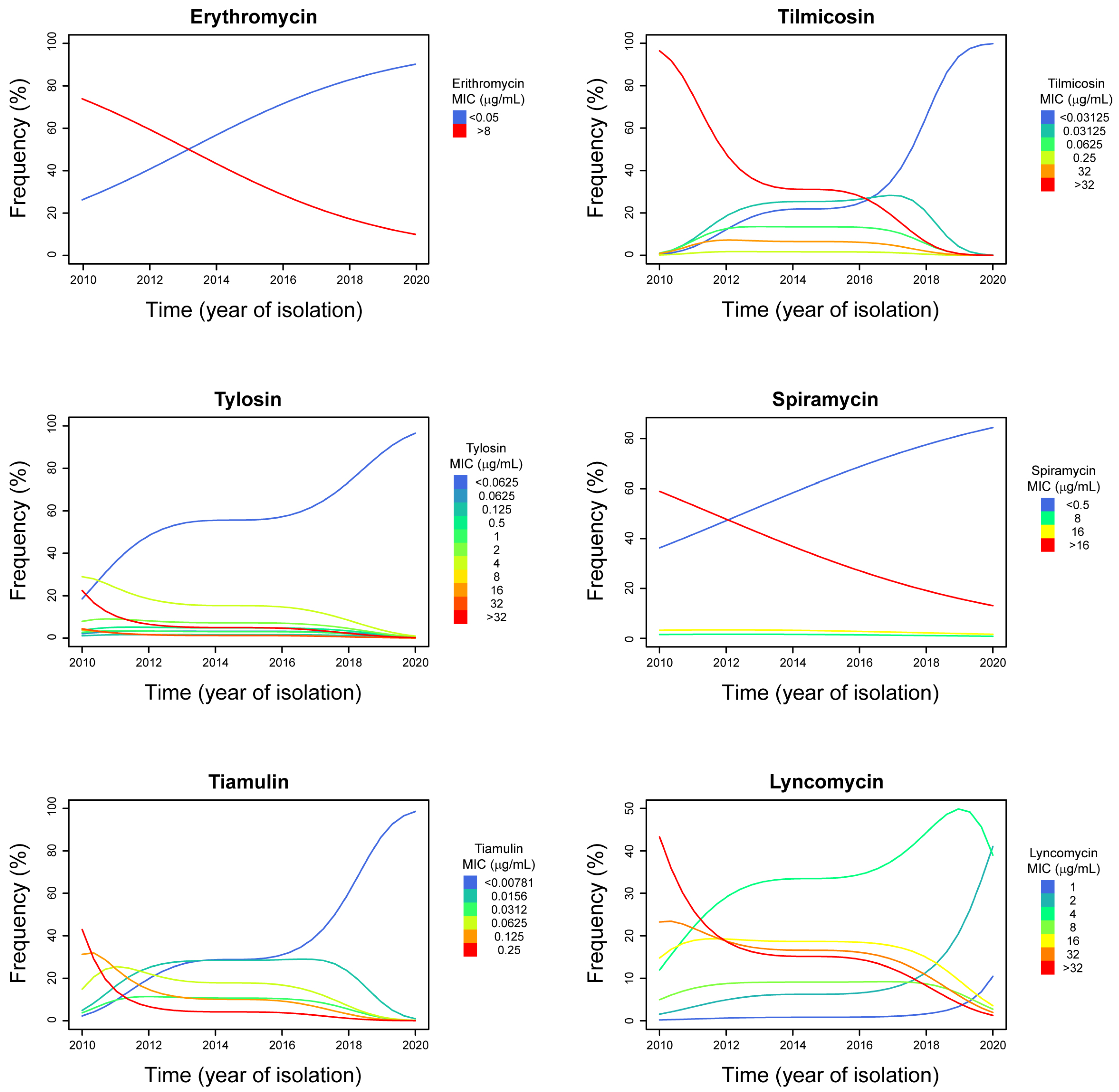
2. Results
3. Discussion
4. Materials and Methods
4.1. Mycoplasma gallisepticum Isolates
4.2. Mycoplasma gallisepticum Cultivation
4.3. Mycoplasma gallisepticum Identification and Genotyping
4.4. MIC Test
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Esri’s StoryMaps Team (Farm) Animal Planet. Available online: https://storymaps.arcgis.com/stories/58ae71f58fd7418294f34c4f841895d8 (accessed on 22 June 2022).
- Gilbert, M.; Nicolas, G.; Cinardi, G.; Van Boeckel, T.P.; Vanwambeke, S.O.; Wint, G.R.W.; Robinson, T.P. Global distribution data for cattle, buffaloes, horses, sheep, goats, pigs, chickens and ducks in 2010. Sci. Data 2018, 5, 180227. [Google Scholar] [CrossRef] [PubMed]
- FAO. Agricultural Production Statistics. 2000–2020; FAOSTAT Analytical Brief Series No. 41. Rome; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/3/cb9180en/cb9180en.pdf (accessed on 29 December 2019).
- Armour, N.K. Mycoplasma gallisepticum infection. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 911–923. ISBN 9781119371168. [Google Scholar]
- Feberwee, A.; de Wit, S.; Dijkman, R. Clinical expression, epidemiology, and monitoring of Mycoplasma gallisepticum and Mycoplasma synoviae: An update. Avian Pathol. 2021, 51, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Kleven, S.H. Control of avian mycoplasma infections in commercial poultry. Avian Dis. 2008, 52, 367–374. [Google Scholar] [CrossRef]
- Landman, W.J.M. Is Mycoplasma synoviae outrunning Mycoplasma gallisepticum? A viewpoint from the Netherlands. Avian Pathol. 2014, 43, 2–8. [Google Scholar] [CrossRef]
- Cummings, T.S.; Kleven, S.H.; Brown, J. Effect of medicated feed on tracheal infection and population of Mycoplasma gallisepticum in chickens. Avian Dis. 1986, 30, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Gigueré, S. Antimicrobial Therapy of Selected Bacterial Infections. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Gigueré, S., Prescott, J., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 421–430. ISBN 9780470963029. [Google Scholar]
- Browning, G.F.; Citti, C. Mollicutes: Molecular Biology and Pathogenesis; Browning, G.F., Citti, C., Eds.; Caister Academic Press: Norfolk, UK, 2014; ISBN 1908230304. [Google Scholar]
- Gautier-Bouchardon, A.V. Antimicrobial Resistance in Mycoplasma spp. Microbiol. Spectr. 2018, 6, ARBA-0030-2018. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Mackie, R.; Aminov, R. Agricultural use of antibiotics and antibiotic resistance. In Antibiotic Resistance Genes in Natural Environments and Long-Term Effects; Mirete, S., Perez, M.L., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; pp. 217–250. [Google Scholar]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial resistance in bacterial poultry pathogens: A review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar] [CrossRef]
- Caucci, C.; Di Martino, G.; Dalla Costa, A.; Santagiuliana, M.; Lorenzetto, M.; Capello, K.; Mughini-Gras, L.; Gavazzi, L.; Bonfanti, L. Trends and correlates of antimicrobial use in broiler and turkey farms: A poultry company registry-based study in Italy. J. Antimicrob. Chemother. 2019, 74, 2784–2787. [Google Scholar] [CrossRef]
- Butenko, I.; Vanyushkina, A.; Pobeguts, O.; Matyushkina, D.; Kovalchuk, S.; Gorbachev, A.; Anikanov, N.; Fisunov, G.; Govorun, V. Response induced in Mycoplasma gallisepticum under heat shock might be relevant to infection process. Sci. Rep. 2017, 7, 11330. [Google Scholar] [CrossRef]
- Benčina, D. Haemagglutinins of pathogenic avian mycoplasmas. Avian Pathol. 2002, 31, 535–547. [Google Scholar] [CrossRef]
- Hofacre, C.L.; Fricke, J.A.; Inglis, T. Antimicrobial Drug Use in Poultry. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Giguère, S., Prescott, J.F., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 569–587. ISBN 9780470963029. [Google Scholar]
- Bradbury, J.M.; Giles, C.J.; Yavari, C.A. In vitro evaluation of various antimicrobials against Mycoplasma gallisepticum and Mycoplasma synoviae by the micro-broth method, and comparison with a commercially-prepared test system. Avian Pathol. 1994, 23, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ferguson-Noel, N.; Armour, N.K.; Noormohammadi, A.H.; El-Gazzar, M.; Bradbury, J.M. Mycoplasmosis. In Diseases of Poultry; Wiley: Hoboken, NJ, USA, 2020; pp. 907–965. [Google Scholar]
- Chernova, O.A.; Chernov, V.M.; Mouzykantov, A.A.; Baranova, N.B.; Edelstein, I.A.; Aminov, R.I. Antimicrobial drug resistance mechanisms among Mollicutes. Int. J. Antimicrob. Agents 2021, 57, 106253. [Google Scholar] [CrossRef] [PubMed]
- Taiyari, H.; Faiz, N.M.; Abu, J.; Zakaria, Z. Antimicrobial minimum inhibitory concentration of Mycoplasma gallisepticum: A systematic review. J. Appl. Poult. Res. 2021, 30, 100160. [Google Scholar] [CrossRef]
- Hannan, P.C.T. Guidelines and recommendations for antimicrobial minimum inhibitory concentration (MIC) testing against veterinary mycoplasma species. Vet. Res. 2000, 31, 373–395. [Google Scholar] [CrossRef] [PubMed]
- Vogl, G.; Plaickner, A.; Szathmary, S.; Stipkovits, L.; Rosengarten, R.; Szostak, M.P. Mycoplasma gallisepticum invades chicken erythrocytes during infection. Infect. Immun. 2008, 76, 71–77. [Google Scholar] [CrossRef]
- Winner, F.; Rosengarten, R.; Citti, C. In vitro cell invasion of Mycoplasma gallisepticum. Infect. Immun. 2000, 68, 4238–4244. [Google Scholar] [CrossRef] [PubMed]
- Much, P.; Winner, F.; Stipkovits, L.; Rosengarten, R.; Citti, C. Mycoplasma gallisepticum: Influence of cell invasiveness on the outcome of experimental infection in chickens. FEMS Immunol. Med. Microbiol. 2002, 34, 181–186. [Google Scholar] [CrossRef] [PubMed]
- del Castillo, J.R.E. Tetracyclines. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Gigueré, S., Prescott, J., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 256–268. ISBN 9780470963029. [Google Scholar]
- Pereyre, S.; Goret, J.; Bébéar, C. Mycoplasma pneumoniae: Current Knowledge on Macrolide Resistance and Treatment. Front. Microbiol. 2016, 7, 974. [Google Scholar] [CrossRef]
- Lysnyansky, I.; Ayling, R.D. Mycoplasma bovis: Mechanisms of Resistance and Trends in Antimicrobial Susceptibility. Front. Microbiol. 2016, 7, 595. [Google Scholar] [CrossRef]
- van der Schalk, T.E.; Braam, J.F.; Kusters, J.G. Molecular basis of antimicrobial resistance in Mycoplasma genitalium. Int. J. Antimicrob. Agents 2020, 55, 105911. [Google Scholar] [CrossRef]
- Bébéar, C.; Pereyre, S.; Peuchant, O. Mycoplasma pneumoniae: Susceptibility and resistance to antibiotics. Future Microbiol. 2011, 6, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Gu, X.; Ye, X.; Wu, X.; Zhang, B.; Zhang, L.; Shen, X.; Jiang, H.; Ding, H. The PK/PD Interactions of Doxycycline against Mycoplasma gallisepticum. Front. Microbiol. 2016, 7, 653. [Google Scholar] [CrossRef] [PubMed]
- Gbylik-Sikorska, M.; Posyniak, A.; Sniegocki, T.; Sell, B.; Gajda, A.; Sawicka, A.; Olszewska-Tomczyk, M.; Bladek, T.; Tomczyk, G.; Zmudzki, J. Influence of enrofloxacin traces in drinking water to doxycycline tissue pharmacokinetics in healthy and infected by Mycoplasma gallisepticum broiler chickens. Food Chem. Toxicol. 2016, 90, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Gigueré, S.; Dowling, P.M. Fluoroquinolones. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Gigueré, S., Prescott, J., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 295–314. ISBN 9780470963029. [Google Scholar]
- EFSA; ECDC. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. EFSA J. 2021, 19, 6490. [Google Scholar] [CrossRef]
- Chniba, I.; Boujemaa, S.; Mardassi, B.B.A.; Ben Abdelmoumen Mardassi, B. Clonal dissemination of antibiotic resistance among Tunisian Mycoplasma gallisepticum isolates as revealed by gene-targeted sequencing analysis. Avian Dis. 2020, 65, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, A.K.; Kempf, I.; Kobisch, M.; Gautier-Bouchardon, A.V. Fluoroquinolone resistance in Mycoplasma gallisepticum: DNA gyrase as primary target of enrofloxacin and impact of mutations in topoisomerases on resistance level. J. Antimicrob. Chemother. 2002, 50, 589–592. [Google Scholar] [CrossRef]
- Khalil, D.; Becker, C.A.M.; Tardy, F. Alterations in the Quinolone Resistance-Determining Regions and Fluoroquinolone Resistance in Clinical Isolates and Laboratory-Derived Mutants of Mycoplasma bovis: Not All Genotypes May Be Equal. Appl. Environ. Microbiol. 2015, 82, 1060–1068. [Google Scholar] [CrossRef]
- Zanella, A.; Martino, P.A.; Pratelli, A.; Stonfer, M. Development of antibiotic resistance in Mycoplasma gallisepticum in vitro. Avian Pathol. 1998, 27, 591–596. [Google Scholar] [CrossRef]
- Gautier-Bouchardon, A.V.; Reinhardt, A.K.; Kobisch, M.; Kempf, I. In vitro development of resistance to enrofloxacin, erythromycin, tylosin, tiamulin and oxytetracycline in Mycoplasma gallisepticum, Mycoplasma iowae and Mycoplasma synoviae. Vet. Microbiol. 2002, 88, 47–58. [Google Scholar] [CrossRef]
- Hannan, P.C.T.; Windsor, G.D.; De Jong, A.; Schmeer, N.; Stegemann, M. Comparative susceptibilities of various animal-pathogenic mycoplasmas to fluoroquinolones. Antimicrob. Agents Chemother. 1997, 41, 2037–2040. [Google Scholar] [CrossRef]
- Gerchman, I.; Lysnyansky, I.; Perk, S.; Levisohn, S. In vitro susceptibilities to fluoroquinolones in current and archived Mycoplasma gallisepticum and Mycoplasma synoviae isolates from meat-type turkeys. Vet. Microbiol. 2008, 131, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Gerchman, I.; Levisohn, S.; Mikula, I.; Manso-Silván, L.; Lysnyansky, I. Characterization of in vivo-acquired resistance to macrolides of Mycoplasma gallisepticum strains isolated from poultry. Vet. Res. 2011, 42, 90. [Google Scholar] [CrossRef] [PubMed]
- de Jong, A.; Youala, M.; Klein, U.; El Garch, F.; Simjee, S.; Moyaert, H.; Rose, M.; Gautier-Bouchardon, A.V.; Catania, S.; Ganapathy, K.; et al. Minimal inhibitory concentration of seven antimicrobials to Mycoplasma gallisepticum and Mycoplasma synoviae isolates from six European countries. Avian Pathol. 2021, 50, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Morrow, C.J.; Kreizinger, Z.; Achari, R.R.; Bekő, K.; Yvon, C.; Gyuranecz, M. Antimicrobial susceptibility of pathogenic mycoplasmas in chickens in Asia. Vet. Microbiol. 2020, 250, 108840. [Google Scholar] [CrossRef]
- Page, S.W.; Gautier, P. Use of antimicrobial agents in livestock. Rev. Sci. Tech. Off. Int. Epiz 2012, 31, 145. Available online: https://doc.woah.org/dyn/portal/index.xhtml?page=alo&aloId=31376 (accessed on 29 December 2019). [CrossRef] [PubMed]
- Gigueré, S. Macrolides, Azalides, and Ketolides. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Gigueré, S., Prescott, J., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 211–232. ISBN 9780470963029. [Google Scholar]
- Catania, S.; Bottinelli, M.; Fincato, A.; Gastaldelli, M.; Barberio, A.; Gobbo, F.; Vicenzoni, G. Evaluation of Minimum Inhibitory Concentrations for 154 Mycoplasma synoviae isolates from Italy collected during 2012–2017. PLoS ONE 2019, 14, e0224903. [Google Scholar] [CrossRef]
- Charleston, B.; Gate, J.J.; Aitken, I.A.; Reeve-Johnson, L. Assessment of the efficacy of tilmicosin as a treatment for Mycoplasma gallisepticum infections in chickens. Avian Pathol. 1998, 27, 190–195. [Google Scholar] [CrossRef][Green Version]
- Warner, K.; Clark, M.I.; Perez, S.; Jennison, R. Control of Ornithobacterium rhinotracheale in poultry. Vet. Rec. 2009, 165, 668. [Google Scholar] [CrossRef]
- Abu-Basha, E.A.; Idkaidek, N.M.; Al-Shunnaq, A.F. Pharmacokinetics of tilmicosin (Provitil powder and Pulmotil liquid AC) oral formulations in chickens. Vet. Res. Commun. 2007, 31, 477–485. [Google Scholar] [CrossRef]
- Jordan, F.T.W.; Horrocks, B.K. The minimum inhibitory concentration of tilmicosin and tylosin for Mycoplasma gallisepticum and Mycoplasma synoviae and a comparison of their efficacy in the control of Mycoplasma gallisepticum infection in broiler chicks. Avian Dis. 1996, 40, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Jordan, F.T.W.; Forrester, C.A.; Hodge, A.; Reeve-Johnson, L.G. The comparison of an aqueous preparation of tilmicosin with tylosin in the treatment of Mycoplasma gallisepticum infection of turkey poults. Avian Dis. 1999, 43, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Kempf, I.; Reeve-Johnson, L.; Gesbert, F.; Guittet, M. Efficacy of tilmicosin in the control of experimental Mycoplasma gallisepticum infection in chickens. Avian Dis. 1997, 41, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Lysnyansky, I.; Gerchman, I.; Levisohn, S.; Mikula, I.; Feberwee, A.; Ferguson, N.M.; Noormohammadi, A.H.; Spergser, J.; Windsor, H.M. Discrepancy between minimal inhibitory concentration to enrofloxacin and mutations present in the quinolone-resistance determining regions of Mycoplasma gallisepticum field strains. Vet. Microbiol. 2012, 160, 222–226. [Google Scholar] [CrossRef]
- Gharaibeh, S.; Al-Rashdan, M. Change in antimicrobial susceptibility of Mycoplasma gallisepticum field isolates. Vet. Microbiol. 2011, 150, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Pakpinyo, S.; Sasipreeyajan, J. Molecular characterization and determination of antimicrobial resistance of Mycoplasma gallisepticum isolated from chickens. Vet. Microbiol. 2007, 125, 59–65. [Google Scholar] [CrossRef]
- Wang, C.; Ewing, M.; A’Arabi, S.Y. In vitro susceptibility of avian Mycoplasmas to enrofloxacin, sarafloxacin, tylosin, and oxytetracycline. Avian Dis. 2001, 45, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Jordan, F.T.W.; Gilbert, S.; Knight, D.L.; Yavari, C.A. Effects of Baytril, Tylosin and Tiamulin on avian mycoplasmas. Avian Pathol. 1989, 18, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Jordan, F.T.W.; Knight, D. The minimum inhibitory concentration of kitasamycin, tylosin and tiamulin for Mycoplasma gallisepticum and their protective effect on infected chicks. Avian Pathol. 1984, 13, 151–162. [Google Scholar] [CrossRef]
- Debets-Ossenkopp, Y.J.; Brinkman, A.B.; Kuipers, E.J.; Vandenbroucke-Grauls, C.M.J.E.; Kusters, J.G. Explaining the bias in the 23S rRNA gene mutations associated with clarithromycin resistance in clinical isolates of Helicobacter pylori. Antimicrob. Agents Chemother. 1998, 42, 2749–2751. [Google Scholar] [CrossRef]
- Wang, G.; Taylor, D.E. Site-specific mutations in the 23S rRNA gene of Helicobacter pylori confer two types of resistance to macrolide-lincosamide-streptogramin B antibiotics. Antimicrob. Agents Chemother. 1998, 42, 1952–1958. [Google Scholar] [CrossRef]
- Matucci, A.; Stefani, E.; Gastaldelli, M.; Rossi, I.; De Grandi, G.; Gyuranecz, M.; Catania, S. Molecular differentiation of Mycoplasma gallisepticum outbreaks: A last decade study on italian farms using gts and mlst. Vaccines 2020, 8, 665. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, L.; Monne, I.; Tamba, M.; Santucci, U.; Massi, P.; Patregnani, T.; Loli Piccolomini, L.; Natalini, S.; Ferri, G.; Cattoli, G.; et al. Highly pathogenic H7N7 avian influenza in Italy. Vet. Rec. 2014, 174, 382. [Google Scholar] [CrossRef]
- Catania, S.; Bilato, D.; Gobbo, F.; Granato, A.; Terregino, C.; Iob, L.; Nicholas, R.A.J. Treatment of Eggshell Abnormalities and Reduced Egg Production Caused by Mycoplasma synoviae Infection. Avian Dis. 2010, 54, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Catania, S.; Gobbo, F.; Bilato, D.; Gagliazzo, L.; Moronato, M.L.; Terregino, C.; Bradbury, J.M.; Ramírez, A.S. Two strains of Mycoplasma synoviae from chicken flocks on the same layer farm differ in their ability to produce eggshell apex abnormality. Vet. Microbiol. 2016, 193, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Gigueré, S. Lincosamides, pleuromutilins and streptogramins. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Gigueré, S., Prescott, J.F., Dowling, P.M., Eds.; John Wiley & Sons, Inc: Ames, IA, USA, 2013; pp. 199–210. [Google Scholar]
- Jordan, F.T.W.; Forrester, A.C.A.; Ripley, P.H.; Burch, D.G.S. In vitro and in vivo comparisons of valnemulin, tiamulin, tylosin, enrofloxacin, and lincomycin/spectinomycin against Mycoplasma gallisepticum. Avian Dis. 1998, 42, 738–745. [Google Scholar] [CrossRef] [PubMed]
- Rosales, R.S.; Ramírez, A.S.; Tavío, M.M.; Poveda, C.; Poveda, J.B. Antimicrobial susceptibility profiles of porcine mycoplasmas isolated from samples collected in southern Europe. BMC Vet. Res. 2020, 16, 324. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Shryock, T.R.; Lin, T.L.; Faderan, M.; Veenhuizen, M.F. Antimicrobial susceptibility of Mycoplasma hyorhinis. Vet. Microbiol. 2000, 76, 25–30. [Google Scholar] [CrossRef]
- Klein, U.; de Jong, A.; Moyaert, H.; El Garch, F.; Leon, R.; Richard-Mazet, A.; Rose, M.; Maes, D.; Pridmore, A.; Thomson, J.R.; et al. Antimicrobial susceptibility monitoring of Mycoplasma hyopneumoniae and Mycoplasma bovis isolated in Europe. Vet. Microbiol. 2017, 204, 188–193. [Google Scholar] [CrossRef]
- Hamdy, A.H.; Saif, Y.M.; Kasson, C.W. Efficacy of lincomycin-spectinomycin water medication on Mycoplasma meleagridis airsacculitis in commercially reared turkey poults. Avian Dis. 1982, 26, 227–233. [Google Scholar] [CrossRef]
- Hamdy, A.H.; Kleven, S.H.; McCune, E.L. Efficacy of Linco-Spectin water medication on Mycoplasma synoviae Airsacculitis in broilers. Avian Dis. 1976, 20, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Dowling, P.M. Chloramphenicol, Thiamphenicol and Florfenicol, 5th ed.; Gigueré, S., Prescott, J., Dowling, P.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 269–277. ISBN 9780470963029. [Google Scholar]
- Papich, M.G. Papich Handbook of Veterinary Drugs: Small and Large Animal; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780323709576. [Google Scholar]
- Bekő, K.; Felde, O.; Sulyok, K.M.; Kreizinger, Z.; Hrivnák, V.; Kiss, K.; Biksi, I.; Jerzsele, Á.; Gyuranecz, M. Antibiotic susceptibility profiles of Mycoplasma hyorhinis strains isolated from swine in Hungary. Vet. Microbiol. 2019, 228, 196–201. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, L.; Ellis, R.J.; Lawes, J.R.; Ayling, R.D.; Nicholas, R.A.J. 16S rDNA PCR and denaturing gradient gel electrophoresis; a single generic test for detecting and differentiating Mycoplasma species. J. Med. Microbiol. 2005, 54, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Lysnyansky, I.; García, M.; Levisohn, S. Use of mgc2-polymerase chain reaction-restriction fragment length polymorphism for rapid differentiation between field isolates and vaccine strains of Mycoplasma gallisepticum in Israel. Avian Dis. 2005, 49, 238–245. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Ikuta, N.; Levisohn, S.; Kleven, S.H. Evaluation and comparison of various PCR methods for detection of Mycoplasma gallisepticum infection in chickens. Avian Dis. 2005, 49, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Leigh, S.A. Differentiation of Mycoplasma gallisepticum vaccine strains ts-11 and 6/85 from commonly used Mycoplasma gallisepticum challenge strains by PCR. Avian Dis. 2008, 52, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. M43-A Methods for Antimicrobial Susceptibility Testing for Human Mycoplasmas; Approved Guideline; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011; Available online: http://shop.clsi.org/site/Sample_pdf/M43A_sample.pdf (accessed on 29 December 2019).
- Markey, B.; Leonard, F.; Archambault, M.; Cullinane, A.; Maguire, D. The Mycoplasmas (class: Mollicutes). In Clinical Veterinary Microbiology, 2nd ed; Markey, B., Leonard, F., Archambault, M., Cullinane, A., Maguire, D., Eds.; Mosby Elsevier: Maryland Heights, MO, USA, 2013; pp. 423–431. [Google Scholar]
- Blodgett, R. FDA’s Bacteriological Analytical Manual, Appendix 2: Most Probable Number from Serial Dilutions. 2010. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-appendix-2-most-probable-number-serial-dilutions (accessed on 29 December 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 29 December 2019).
- Hothorn, T.; Hornik, K.; van de Wiel, M.A.; Zeileis, A. Implementing a class of permutation tests: The coin package. J. Stat. Softw. 2008, 28, 1–23. [Google Scholar] [CrossRef]
- Christensen, R.H.B. ordinal—Regression Models for Ordinal Data. R Package Version 2015, 28, 2015. [Google Scholar]


{kind=link}
| Antibiotic | MIC Values (µg/ML) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.003906 | 0.0078125 | 0.015625 | 0.03125 | 0.0625 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | |
| Oxytetracycline | 9 | 14 | 2650 | 9 | 390 | 5 | 1 * | ||||||||
| Doxycycline | 3 | 2550 | 18 | 590 | |||||||||||
| Enrofloxacin | 3 | 4 | 5 | 2 | 6 | 9 | 3850−90 | ||||||||
| Erythromycin | 3950 | 2890 | |||||||||||||
| Tilmicosin | 31 | 750 | 1 | 4 | 2490 | ||||||||||
| Tylosin | 6 | 14 | 1550 | 3 | 1 | 3 | 2 | 5 | 1190 | 1 | 1 | 1 | 4 | ||
| Spiramycin | 3950 | 1 | 2 | 2590 | |||||||||||
| Tiamulin | 25 | 1350 | 6 | 11 | 890 | 4 | |||||||||
| Lincomycin | 1 | 6 | 20 | 6 | 1250 | 11 | 1190 | ||||||||
| Florfenicol | 5 | 27 | 3250−90 | 3 | |||||||||||
| Antibiotic | Z-value | p-Value | Adjusted p-Value |
|---|---|---|---|
| Oxytetracycline | −0.97 | 0.33 | 0.38 |
| Enrofloxacin | −1.53 | 8.65⋅10−2 | 0.11 |
| Erythromycin | −3.14 | 1.71⋅10−3 | 5.12⋅10−3 |
| Tilmicosin | −3.95 | 7.87⋅10−5 | 3.54⋅10−4 |
| Tylosin | −2.82 | 4.84⋅10−3 | 1.09⋅10−2 |
| Spiramycin | −2.65 | 7.97⋅10−3 | 1.43⋅10−2 |
| Tiamulin | −4.18 | 2.94⋅10−5 | 2.65⋅10−4 |
| Lyncomycin | −2.57 | 1.03⋅10−2 | 1.55⋅10−2 |
| Florfenicol | 0.45 | 0.65 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bottinelli, M.; Gastaldelli, M.; Picchi, M.; Dall’Ora, A.; Cristovao Borges, L.; Ramírez, A.S.; Matucci, A.; Catania, S. The Monitoring of Mycoplasma gallisepticum Minimum Inhibitory Concentrations during the Last Decade (2010–2020) Seems to Reveal a Comeback of Susceptibility to Macrolides, Tiamulin, and Lincomycin. Antibiotics 2022, 11, 1021. https://doi.org/10.3390/antibiotics11081021
Bottinelli M, Gastaldelli M, Picchi M, Dall’Ora A, Cristovao Borges L, Ramírez AS, Matucci A, Catania S. The Monitoring of Mycoplasma gallisepticum Minimum Inhibitory Concentrations during the Last Decade (2010–2020) Seems to Reveal a Comeback of Susceptibility to Macrolides, Tiamulin, and Lincomycin. Antibiotics. 2022; 11(8):1021. https://doi.org/10.3390/antibiotics11081021
Chicago/Turabian StyleBottinelli, Marco, Michele Gastaldelli, Micaela Picchi, Arianna Dall’Ora, Lorena Cristovao Borges, Ana Sofía Ramírez, Andrea Matucci, and Salvatore Catania. 2022. "The Monitoring of Mycoplasma gallisepticum Minimum Inhibitory Concentrations during the Last Decade (2010–2020) Seems to Reveal a Comeback of Susceptibility to Macrolides, Tiamulin, and Lincomycin" Antibiotics 11, no. 8: 1021. https://doi.org/10.3390/antibiotics11081021
APA StyleBottinelli, M., Gastaldelli, M., Picchi, M., Dall’Ora, A., Cristovao Borges, L., Ramírez, A. S., Matucci, A., & Catania, S. (2022). The Monitoring of Mycoplasma gallisepticum Minimum Inhibitory Concentrations during the Last Decade (2010–2020) Seems to Reveal a Comeback of Susceptibility to Macrolides, Tiamulin, and Lincomycin. Antibiotics, 11(8), 1021. https://doi.org/10.3390/antibiotics11081021