
Leucyl-tRNA Synthetase Inhibitor, D-Norvaline, in Combination with Oxacillin, Is Effective against Methicillin-Resistant Staphylococcus aureus

 , ,
, ,
Abstract
:1. Introduction
2. Results
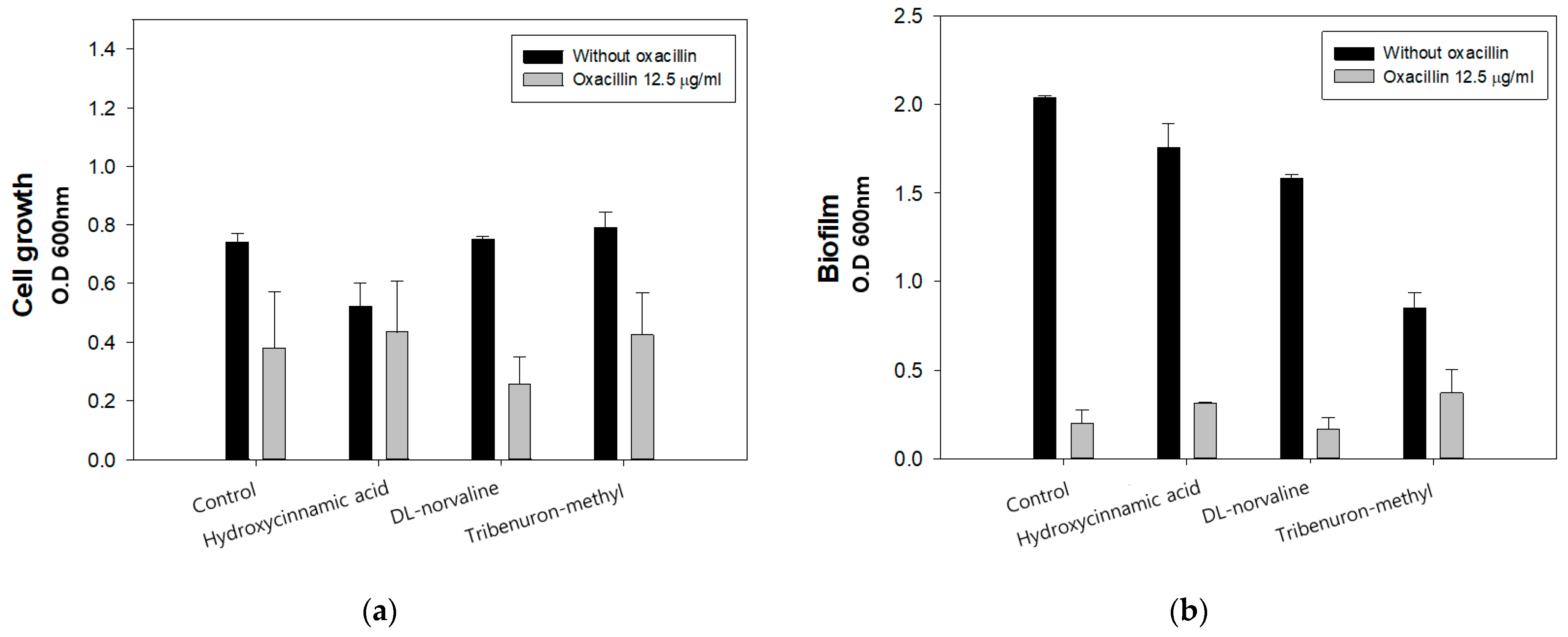
2.1. Compound Screening Inhibiting Cell Growth of LAC with Oxacillin
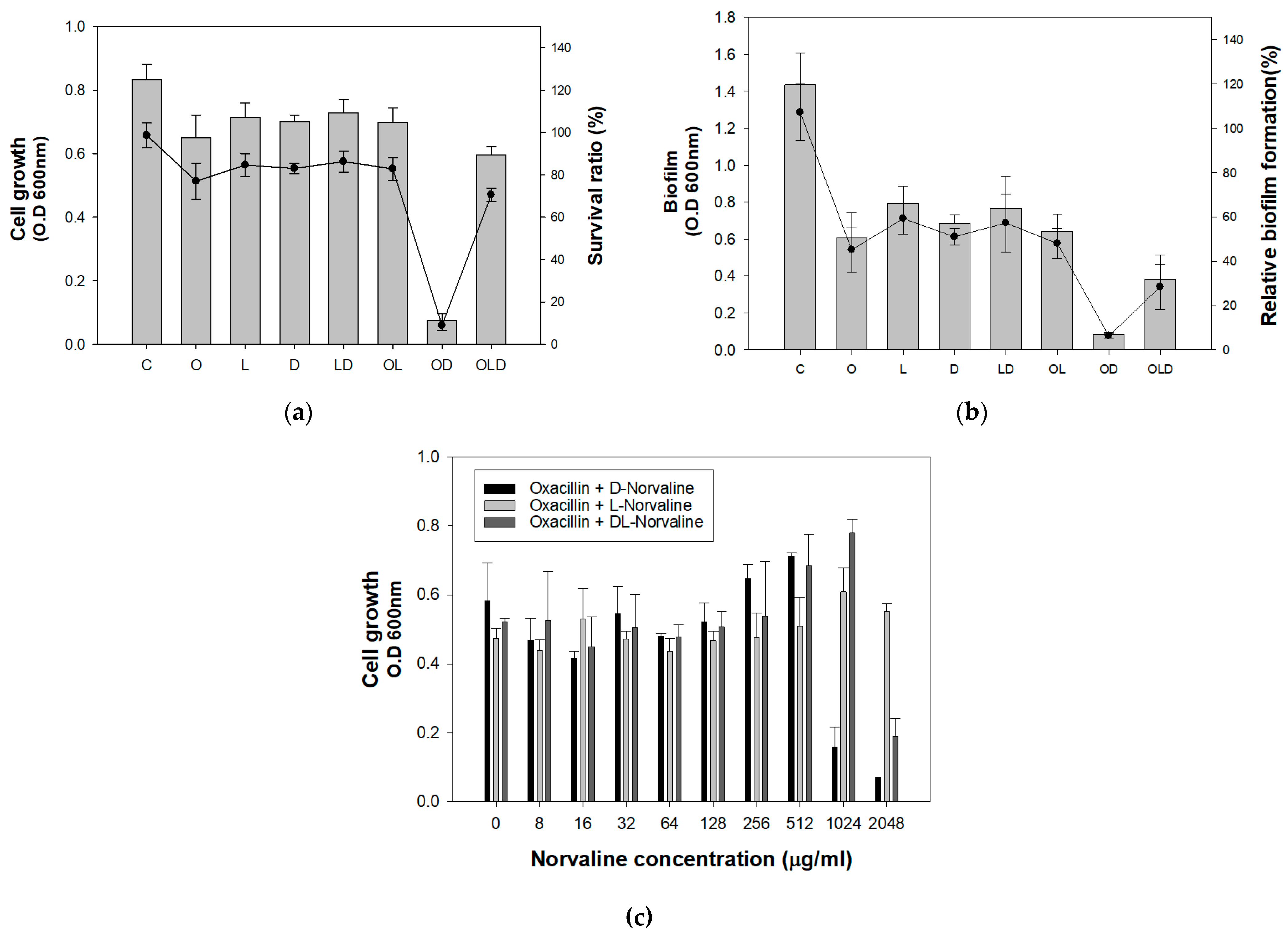
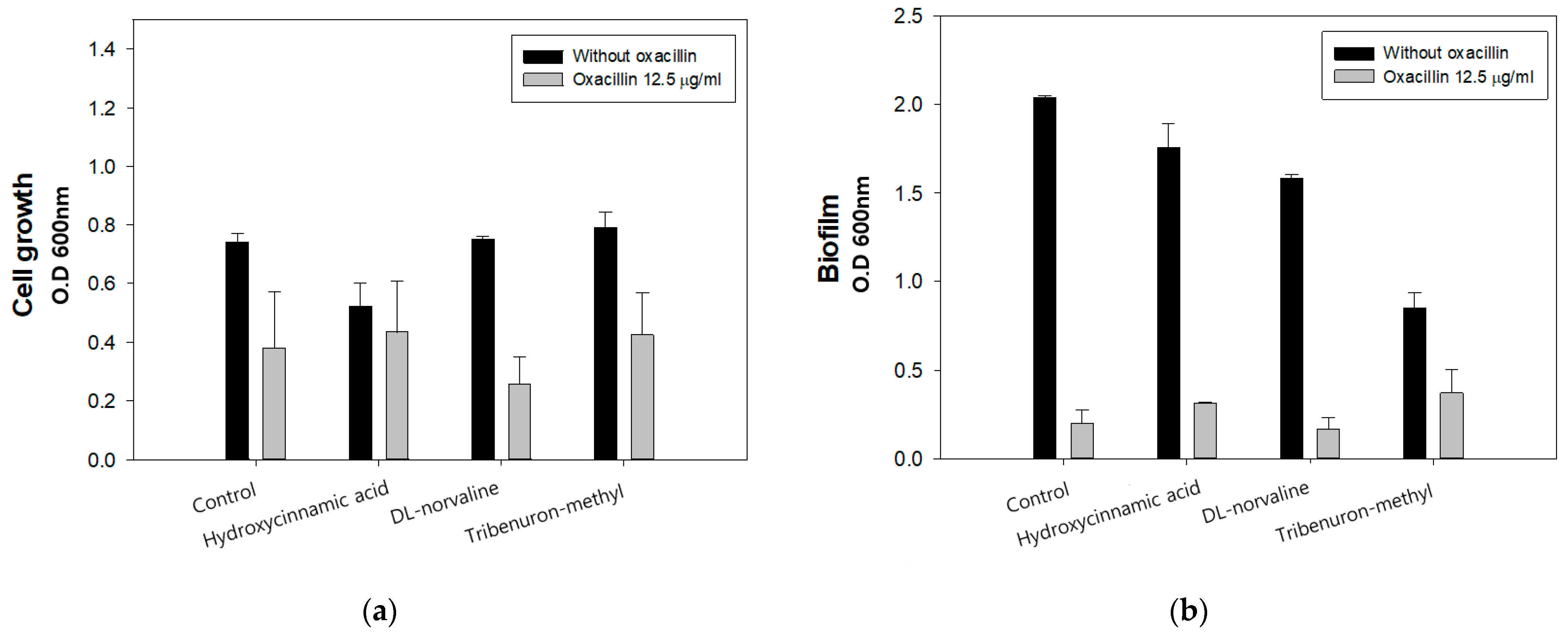
2.2. Inhibitory Effect of Norvaline with Oxacillin
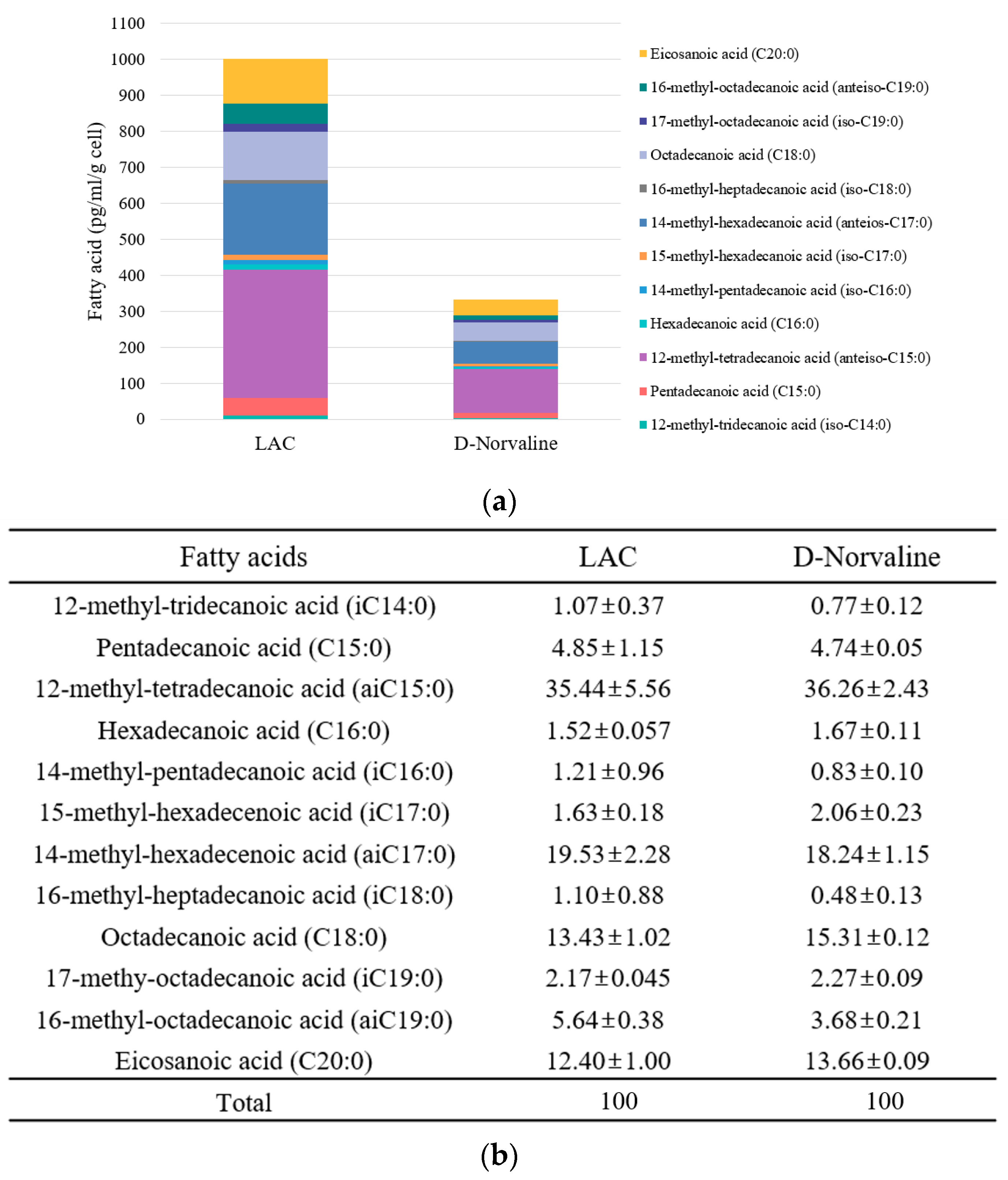
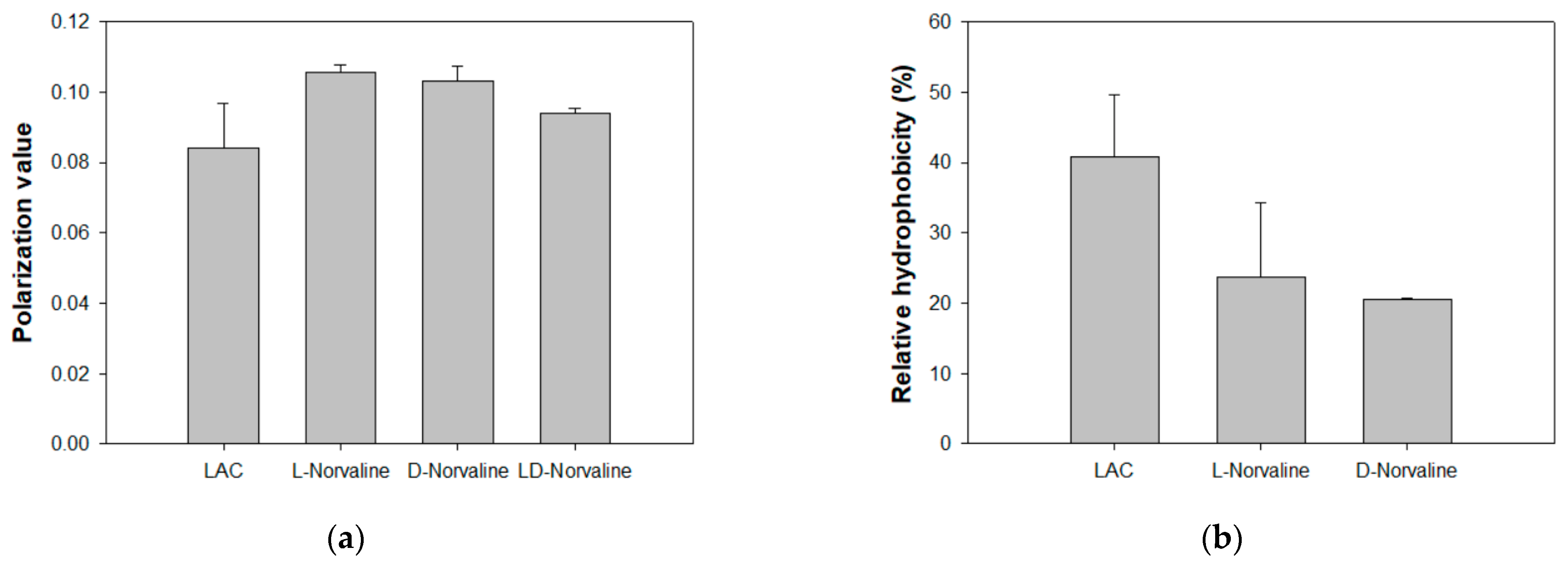
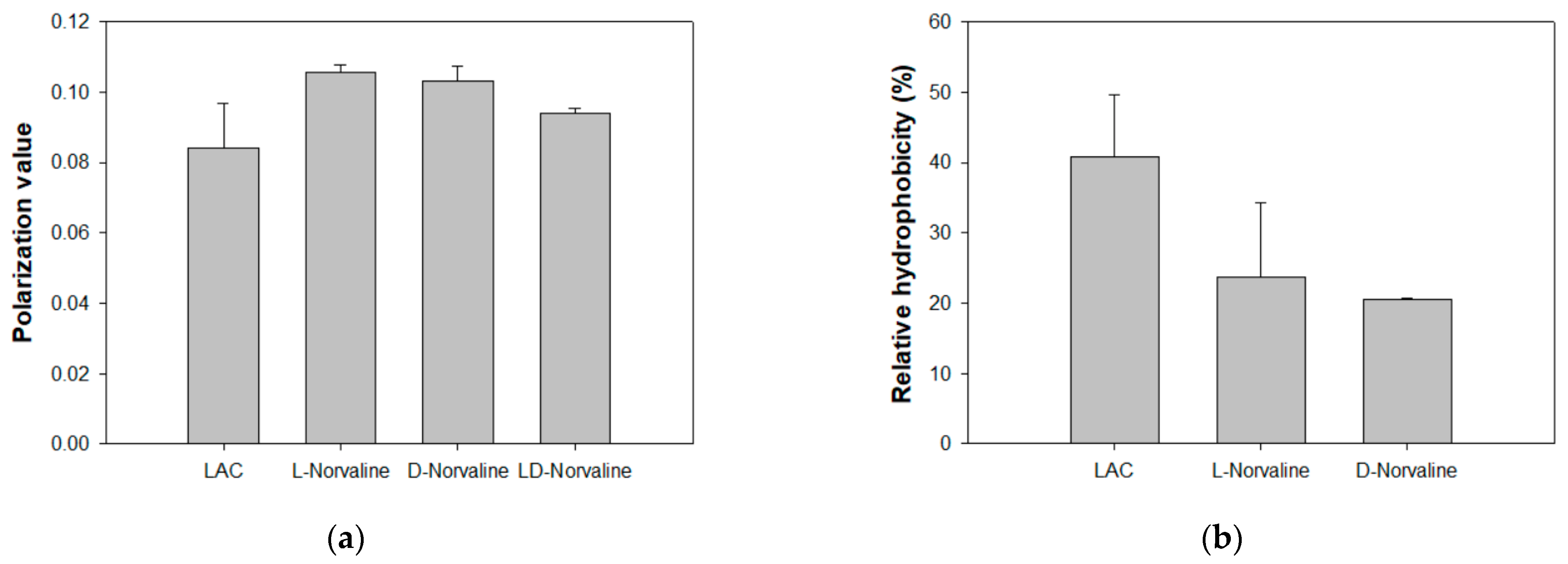
2.3. Effect of D-Norvaline on Phospholipid Fatty Acid Composition and Membrane Properties of LAC
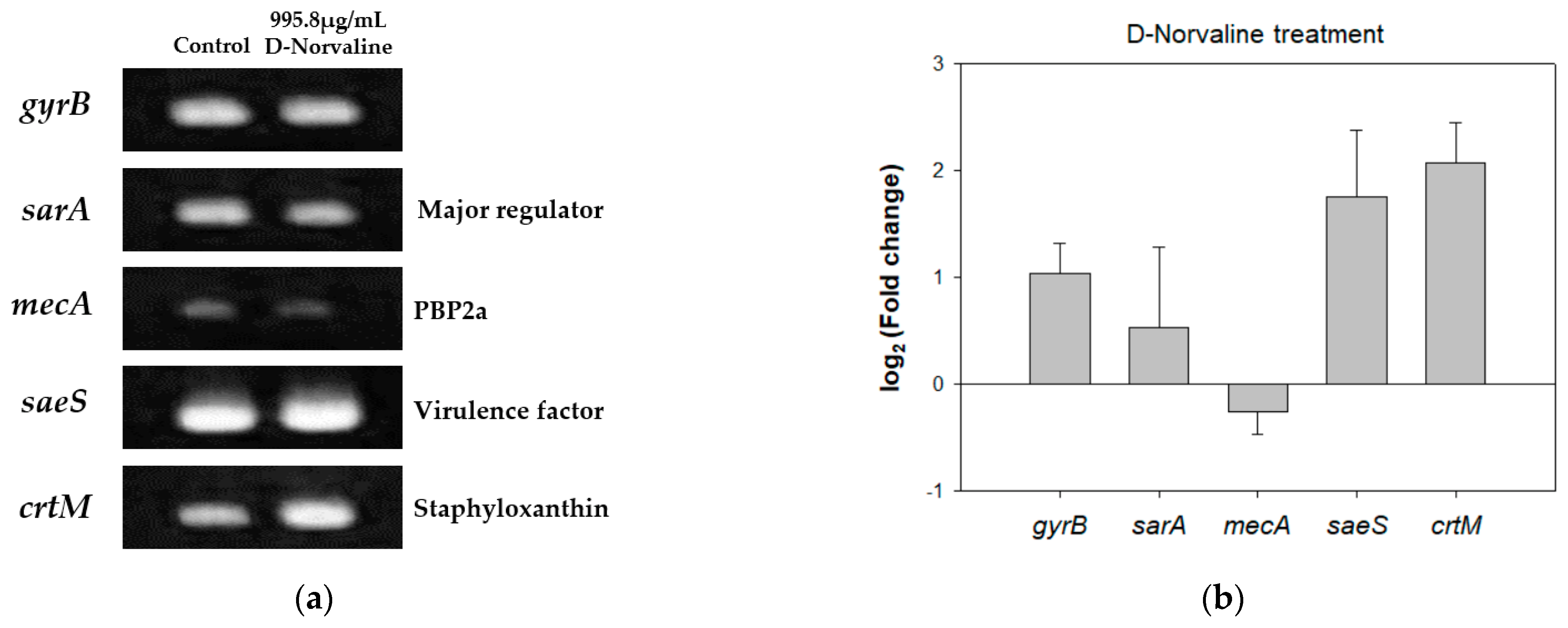
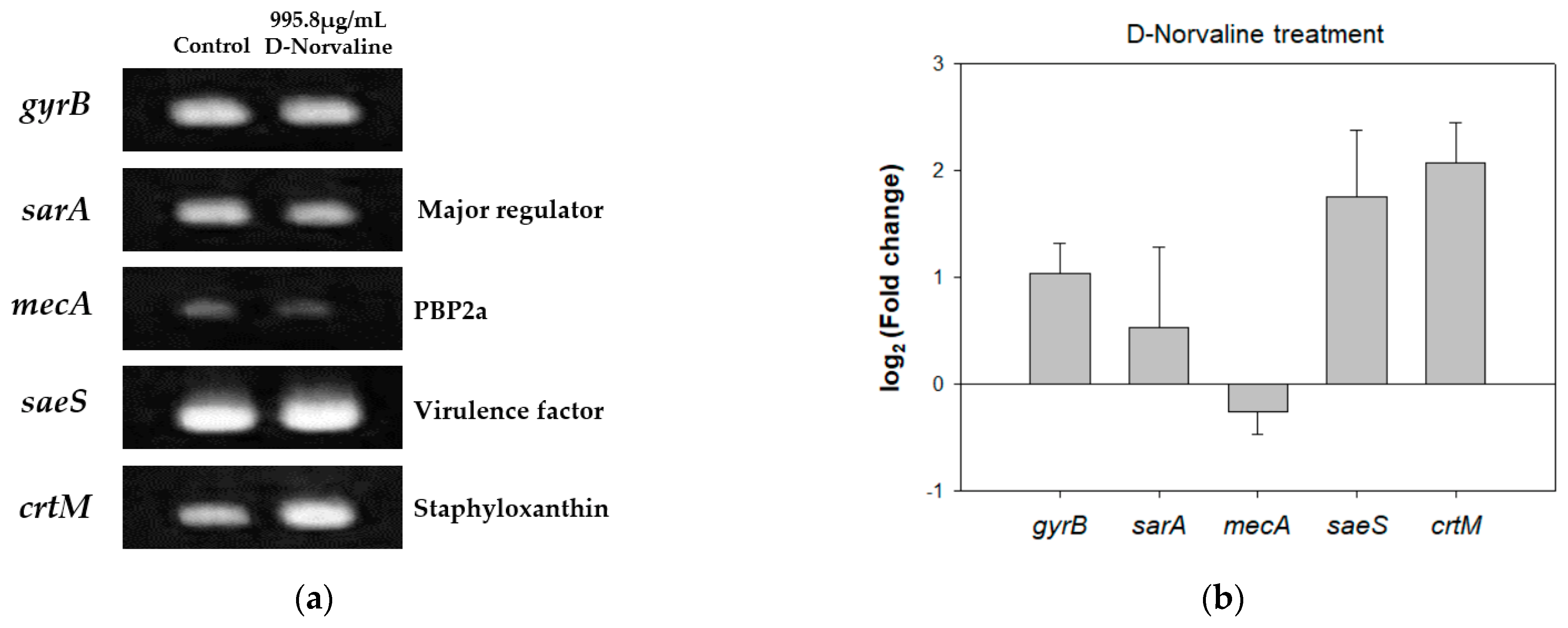
2.4. Different Gene Expression Patterns of LAC with D-Norvaline and CHANGES in Phenotypes
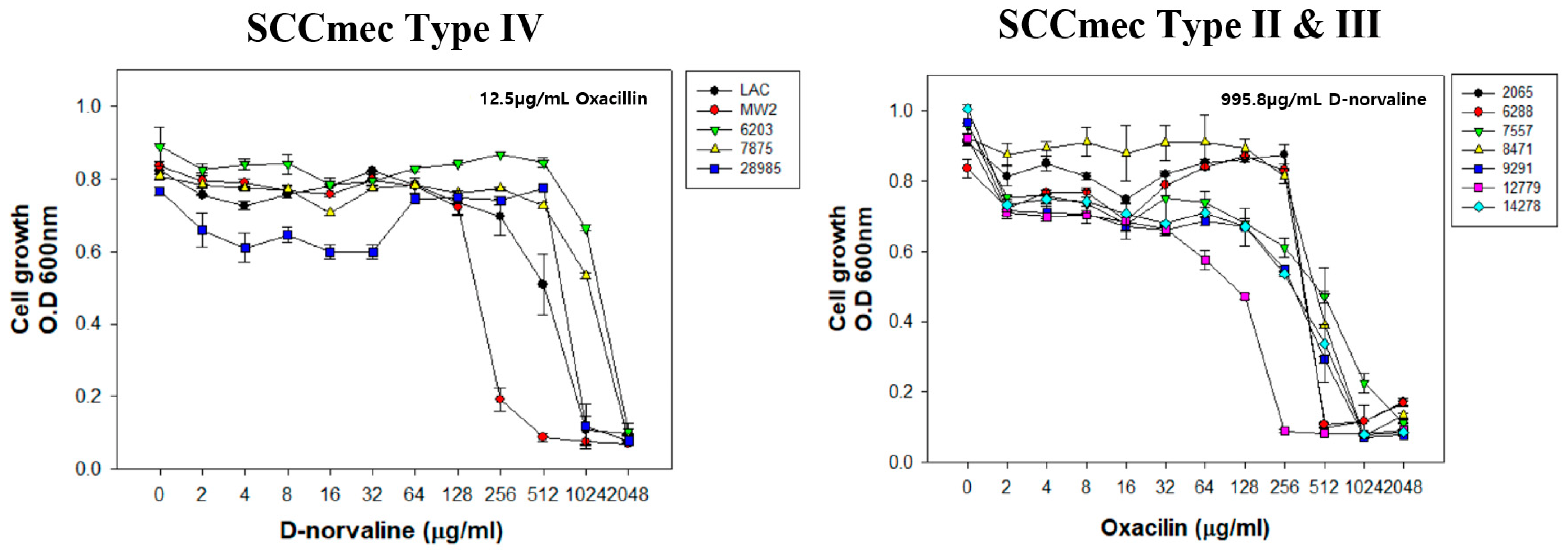
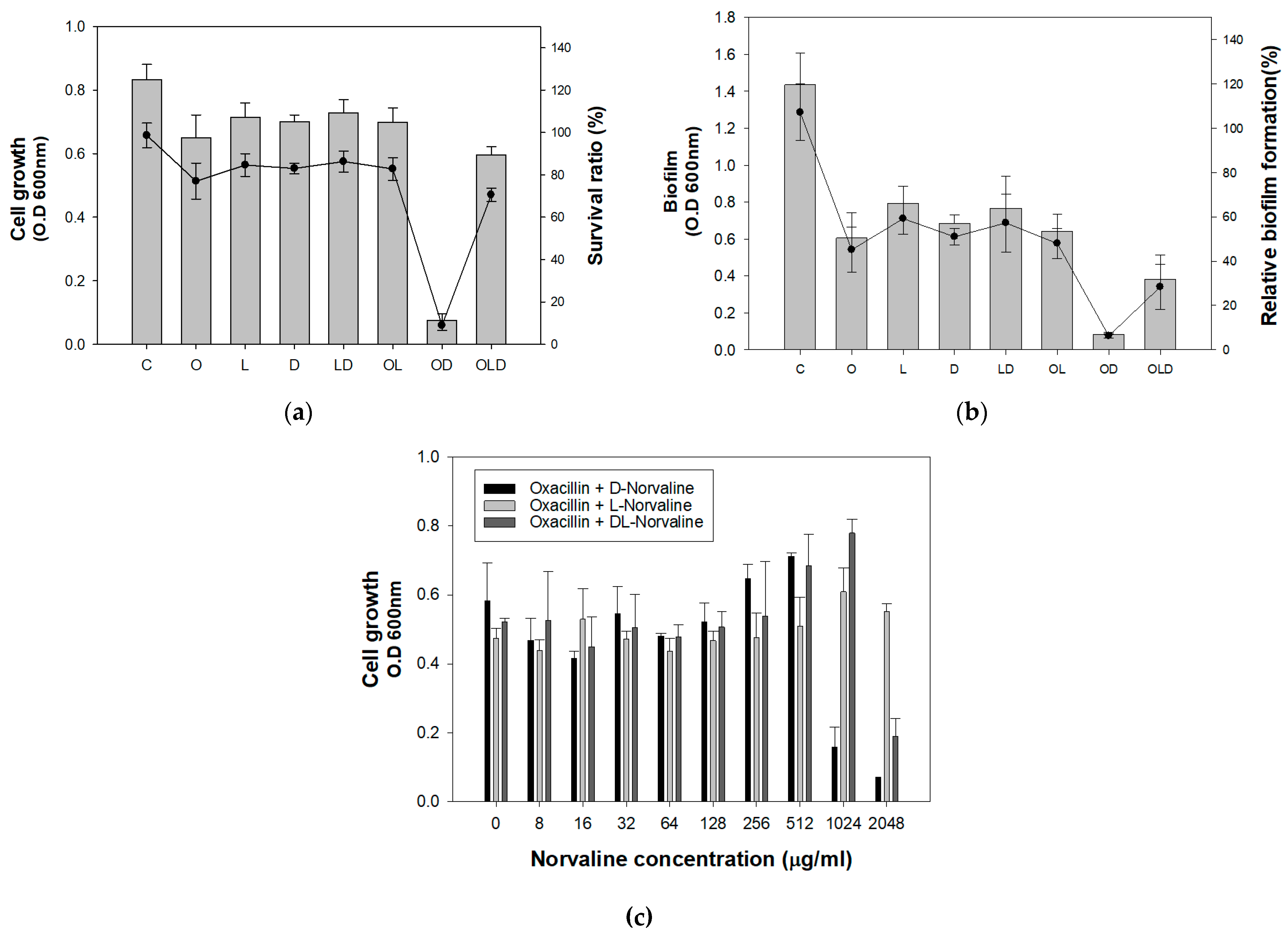
2.5. D-Norvaline on Deletion Mutant Strains and Clinically Isolated MRSA Strains
3. Discussion
4. Materials and Methods
4.1. S. aureus Strain, Media, and Culture Condition
4.2. Minimum Inhibitory Concentration (MIC) and Biofilm Formation Test
4.3. Screening of Inhibitory Compound in LAC Strain
4.4. Phospholipid Fatty Acid (PLFA) Analysis
4.5. Analysis of Membrane Hydrophobicity and Fluidity
4.6. Quantification of Staphyloxanthin (STX)
4.7. Semi-Quantitative RT-PCR
4.8. Quantitative Reverse Transcription PCR (RT-qPCR)
4.9. Plate-Based Motility and Protease Activity Test
4.10. Checkerboard MIC Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Abbreviation in This Paper | Explanation |
| aaRS | Aminoacyl-tRNA synthetase |
| BCAAs | Branched-chain amino acids |
| BCFAs | Branched-chain fatty acids |
| CA-MRSA | Community-associated MRSA |
| GC-MS | Gas chromatography-mass spectrometry |
| HA-MRSA | Healthcare-associated MRSA |
| HCA | Hydroxycinnamic acid |
| LeuRS | Leucyl-tRNA synthetase |
| MIC | Minimum inhibitory concentration |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| PLFA | Phospholipid fatty acid |
| RT-PCR | Reverse transcription PCR |
| RT-qPCR | Quantitative reverse transcription PCR |
| S. aureus | Staphylococcus aureus |
| SCCmec | Staphylococcal cassette chromosome mec |
| SCFAs | Straight-chain fatty acids |
| STX | Staphyloxanthin |
| TCA | Trans-cinnamaldehyde |
| TSA | Tryptic soy agar |
| TSB | Tryptic soybean broth |
References
- Pollitt, E.J.G.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus aureus Infection Dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giulieri, S.G.; Guérillot, R.; Kwong, J.C.; Monk, I.R.; Hayes, A.S.; Daniel, D.; Baines, S.; Sherry, N.L.; Holmes, N.E.; Ward, P.; et al. Comprehensive Genomic Investigation of Adaptive Mutations Driving the Low-Level Oxacillin Resistance Phenotype in Staphylococcus aureus. mBio 2020, 11, e02882-20. [Google Scholar] [CrossRef]
- Kim, W.; Hendricks, G.L.; Tori, K.; Fuchs, B.B.; Mylonakis, E. Strategies against Methicillin-Resistant Staphylococcus aureus Persisters. Future Med. Chem. 2018, 10, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.B.; Montgomery, C.P.; Boyle-Vavra, S.; Shatzkes, K.; Maybank, R.; Frank, B.C.; Peterson, S.N.; Daum, R.S. Genomic and Transcriptomic Differences in Community Acquired Methicillin Resistant Staphylococcus aureus USA300 and USA400 Strains. BMC Genom. 2014, 15, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhungel, S.; Rijal, K.R.; Yadav, B.; Dhungel, B.; Adhikari, N.; Shrestha, U.T.; Adhikari, B.; Banjara, M.R.; Ghimire, P. Methicillin-Resistant Staphylococcus aureus (MRSA): Prevalence, Antimicrobial Susceptibility Pattern, and Detection of Mec A Gene among Cardiac Patients from a Tertiary Care Heart Center in Kathmandu, Nepal. Infect. Dis. Res. Treat. 2021, 14, 117863372110373. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.; Shashindran, N.; Kumar, A.; Vinodh, V.; Biswas, L.; Biswas, R. Comparison of Phenotypic MRSA Detection Methods with PCR for MecA Gene in the Background of Emergence of Oxacillin-Susceptible MRSA. Microb. Drug Resist. 2021, 27, 1190–1194. [Google Scholar] [CrossRef]
- Chambers, H.F.; DeLeo, F.R. Waves of Resistance: Staphylococcus aureus in the Antibiotic Era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Bassetti, M.; Nicco, E.; Mikulska, M. Why Is Community-Associated MRSA Spreading across the World and How Will It Change Clinical Practice? Int. J. Antimicrob. Agents 2009, 34, S15–S19. [Google Scholar] [CrossRef]
- Cascioferro, S.; Carbone, D.; Parrino, B.; Pecoraro, C.; Giovannetti, E.; Cirrincione, G.; Diana, P. Therapeutic Strategies to Counteract Antibiotic Resistance in MRSA Biofilm-Associated Infections. ChemMedChem 2021, 16, 65–80. [Google Scholar] [CrossRef]
- Hadera, M.; Mehari, S.; Basha, N.S.; Amha, N.D.; Berhane, Y. Study on Antimicrobial Potential of Selected Non-Antibiotics and Its Interaction with Conventional Antibiotics. Pharm. Biosci. J. 2018, 6, 1–7. [Google Scholar] [CrossRef]
- Xu, H.-X.; Lee, S.F. Activity of Plant Flavonoids against Antibiotic-Resistant Bacteria. Phytother. Res. 2001, 15, 39–43. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I. Prospects for Aminoacyl-TRNA Synthetase Inhibitors as New Antimicrobial Agents. Antimicrob. Agents Chemother. 2005, 49, 4821–4833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, C.; Rauter, A.P. Membrane-Targeting Antibiotics: Recent Developments Outside the Peptide Space. Future Med. Chem. 2019, 11, 211–228. [Google Scholar] [CrossRef]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial Peptides: Interaction with Model and Biological Membranes and Synergism with Chemical Antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-H.; Hofferek, V.; Separovic, F.; Reid, G.E.; Aguilar, M.-I. The Role of Bacterial Lipid Diversity and Membrane Properties in Modulating Antimicrobial Peptide Activity and Drug Resistance. Curr. Opin. Chem. Biol. 2019, 52, 85–92. [Google Scholar] [CrossRef]
- Lee, H.S.; Song, H.S.; Lee, H.J.; Kim, S.H.; Suh, M.J.; Cho, J.Y.; Ham, S.; Kim, Y.G.; Joo, H.S.; Kim, W.; et al. Comparative Study of the Difference in Behavior of the Accessory Gene Regulator (Agr) in USA300 and USA400 Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA). J. Microbiol. Biotechnol. 2021, 31, 1060–1068. [Google Scholar] [CrossRef]
- Song, H.S.; Bhatia, S.K.; Choi, T.R.; Gurav, R.; Kim, H.J.; Lee, S.M.; Park, S.L.; Lee, H.S.; Joo, H.S.; Kim, W.; et al. Increased Antibiotic Resistance of Methicillin-Resistant Staphylococcus aureus USA300 Δpsm Mutants and a Complementation Study of Δpsm Mutants Using Synthetic Phenol-Soluble Modulins. J. Microbiol. Biotechnol. 2021, 31, 115–122. [Google Scholar] [CrossRef]
- Song, H.S.; Choi, T.R.; Bhatia, S.K.; Lee, S.M.; Park, S.L.; Lee, H.S.; Kim, Y.G.; Kim, J.S.; Kim, W.; Yang, Y.H. Combination Therapy Using Low-Concentration Oxacillin with Palmitic Acid and Span85 to Control Clinical Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 682. [Google Scholar] [CrossRef]
- Coman, V.; Vodnar, D.C. Hydroxycinnamic Acids and Human Health: Recent Advances. J. Sci. Food Agric. 2020, 100, 483–499. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.-K.; Verpoorte, R. Biosynthesis, Natural Sources, Dietary Intake, Pharmacokinetic Properties, and Biological Activities of Hydroxycinnamic Acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef] [PubMed]
- Wrzesińska, B.; Praczyk, T. Genetic Variability of Acetolactate Synthase (ALS) Sequence in Centaurea Cyanus Plants Resistant and Susceptible to Tribenuron-Methyl. Agronomy 2021, 11, 2311. [Google Scholar] [CrossRef]
- Nyamai, D.W.; Tastan Bishop, Ö. Aminoacyl TRNA Synthetases as Malarial Drug Targets: A Comparative Bioinformatics Study. Malar. J. 2019, 18, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faghih, O.; Zhang, Z.; Ranade, R.M.; Gillespie, J.R.; Creason, S.A.; Huang, W.; Shibata, S.; Barros-Álvarez, X.; Verlinde, C.L.M.J.; Hol, W.G.J.; et al. Development of Methionyl-TRNA Synthetase Inhibitors as Antibiotics for Gram-Positive Bacterial Infections. Antimicrob. Agents Chemother. 2017, 61, e00999-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Ma, S. Recent Development of Leucyl-TRNA Synthetase Inhibitors as Antimicrobial Agents. MedChemComm 2019, 10, 1329–1341. [Google Scholar] [CrossRef]
- Dahl, J.L.; Arora, K.; Boshoff, H.I.; Whiteford, D.C.; Pacheco, S.A.; Walsh, O.J.; Lau-Bonilla, D.; Davis, W.B.; Garza, A.G. The RelA Homolog of Mycobacterium Smegmatis Affects Cell Appearance, Viability, and Gene Expression. J. Bacteriol. 2005, 187, 2439–2447. [Google Scholar] [CrossRef] [Green Version]
- Eymann, C.; Homuth, G.; Scharf, C.; Hecker, M. Bacillus Subtilis Functional Genomics: Global Characterization of the Stringent Response by Proteome and Transcriptome Analysis. J. Bacteriol. 2002, 184, 2500–2520. [Google Scholar] [CrossRef] [Green Version]
- Wheadon, M.J.; Townsend, C.A. Evolutionary and Functional Analysis of an NRPS Condensation Domain Integrates β-Lactam, d-Amino Acid, and Dehydroamino Acid Synthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2026017118. [Google Scholar] [CrossRef]
- Youssef, N.; Susko, E.; Roger, A.J.; Bielawski, J.P. Evolution of Amino Acid Propensities under Stability-Mediated Epistasis. Mol. Biol. Evol. 2022, 39, msac030. [Google Scholar] [CrossRef]
- Tawfik, D.S.; Gruic-Sovulj, I. How Evolution Shapes Enzyme Selectivity—Lessons from Aminoacyl-tRNA Synthetases and Other Amino Acid Utilizing Enzymes. FEBS J. 2020, 287, 1284–1305. [Google Scholar] [CrossRef] [Green Version]
- Bessa, L.J.; Ferreira, M.; Gameiro, P. Evaluation of Membrane Fluidity of Multidrug-Resistant Isolates of Escherichia coli and Staphylococcus aureus in Presence and Absence of Antibiotics. J. Photochem. Photobiol. B Biol. 2018, 181, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Cvetesic, N.; Palencia, A.; Halasz, I.; Cusack, S.; Gruic-Sovulj, I. The Physiological Target for Leu RS Translational Quality Control Is Norvaline. EMBO J. 2014, 33, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, C.P.; Boyle-Vavra, S.; Daum, R.S. Importance of the Global Regulators Agr and SaeRS in the Pathogenesis of CA-MRSA USA300 Infection. PLoS ONE 2010, 5, e15177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valliammai, A.; Sethupathy, S.; Ananthi, S.; Priya, A.; Selvaraj, A.; Nivetha, V.; Aravindraja, C.; Mahalingam, S.; Pandian, S.K. Proteomic Profiling Unveils Citral Modulating Expression of IsaA, CodY and SaeS to Inhibit Biofilm and Virulence in Methicillin-Resistant Staphylococcus aureus. Int. J. Biol. Macromol. 2020, 158, 208–221. [Google Scholar] [CrossRef]
- Jo, S.H.; Song, W.S.; Park, H.G.; Lee, J.S.; Jeon, H.J.; Lee, Y.H.; Kim, W.; Joo, H.S.; Yang, Y.H.; Kim, J.S.; et al. Multi-Omics Based Characterization of Antibiotic Response in Clinical Isogenic Isolates of Methicillin-Susceptible/-Resistant: Staphylococcus aureus. RSC Adv. 2020, 10, 27864–27873. [Google Scholar] [CrossRef]
- Rudkin, J.K.; Laabei, M.; Edwards, A.M.; Joo, H.-S.; Otto, M.; Lennon, K.L.; O’Gara, J.P.; Waterfield, N.R.; Massey, R.C. Oxacillin Alters the Toxin Expression Profile of Community-Associated Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 1100–1107. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kang, O.-H.; Kwon, D.-Y.; Moreno, D.A.; Ruiz-Alcaraz, A.J. Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 2752. [Google Scholar] [CrossRef]
- Song, H.S.; Choi, T.R.; Han, Y.H.; Park, Y.L.; Park, J.Y.; Yang, S.Y.; Bhatia, S.K.; Gurav, R.; Kim, Y.G.; Kim, J.S.; et al. Increased Resistance of a Methicillin-Resistant Staphylococcus aureus Δagr Mutant with Modified Control in Fatty Acid Metabolism. AMB Express 2020, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Choi, T.-R.; Song, H.-S.; Han, Y.-H.; Park, Y.-L.; Park, J.Y.; Yang, S.-Y.; Bhatia, S.K.; Gurav, R.; Kim, H.J.; Lee, Y.K.; et al. Enhanced Tolerance to Inhibitors of Escherichia coli by Heterologous Expression of Cyclopropane-Fatty Acid-Acyl-Phospholipid Synthase (Cfa) from Halomonas Socia. Bioprocess Biosyst. Eng. 2020, 43, 909–918. [Google Scholar] [CrossRef]
- Royce, L.A.; Liu, P.; Stebbins, M.J.; Hanson, B.C.; Jarboe, L.R. The Damaging Effects of Short Chain Fatty Acids on Escherichia coli Membranes. Appl. Microbiol. Biotechnol. 2013, 97, 8317–8327. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Type | SCCmec Type | Oxacillin MIC (µg/mL) | Spa Type | MLST (ST) | |
|---|---|---|---|---|---|---|
| Without D-Norvaline | With 8.5 mM D-Norvaline | |||||
| LAC | MRSA | IV | 32 | 12.5 | t008 | 8 |
| MW2 | MRSA | IV | 32 | 12.5 | t131 | 1 |
| 2065 | MRSA | III | 1024 | 512 | t037 | 239 |
| 6230 | MRSA | IV | 128 | 32 | t324 | 72 |
| 6288 | MRSA | III | 1024 | 512 | t037 | 239 |
| 7557 | MRSA | II | 1024 | 1024 | t9353 | 5 |
| 7875 | MRSA | IV | 128 | 32 | t664 | 72 |
| 8471 | MRSA | II | 1024 | 1024 | t9353 | 5 |
| 9291 | MRSA | II | 1024 | 1024 | t601 | 5 |
| 12779 | MRSA | II | 1024 | 256 | t2460 | 5 |
| 14278 | MRSA | II | 1024 | 1024 | t9353 | 5 |
| 14459 | MRSA | IV | 1024 | 12.5 | t324 | 72 |
| 28985 | MRSA | IV | 64 | 12.5 | - | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-J.; Kim, B.; Kim, S.; Cho, D.-H.; Jung, H.; Kim, W.; Kim, Y.-G.; Kim, J.-S.; Joo, H.-S.; Lee, S.-H.; et al. Leucyl-tRNA Synthetase Inhibitor, D-Norvaline, in Combination with Oxacillin, Is Effective against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2022, 11, 683. https://doi.org/10.3390/antibiotics11050683
Lee H-J, Kim B, Kim S, Cho D-H, Jung H, Kim W, Kim Y-G, Kim J-S, Joo H-S, Lee S-H, et al. Leucyl-tRNA Synthetase Inhibitor, D-Norvaline, in Combination with Oxacillin, Is Effective against Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2022; 11(5):683. https://doi.org/10.3390/antibiotics11050683
Chicago/Turabian StyleLee, Hong-Ju, Byungchan Kim, Suhyun Kim, Do-Hyun Cho, Heeju Jung, Wooseong Kim, Yun-Gon Kim, Jae-Seok Kim, Hwang-Soo Joo, Sang-Ho Lee, and et al. 2022. "Leucyl-tRNA Synthetase Inhibitor, D-Norvaline, in Combination with Oxacillin, Is Effective against Methicillin-Resistant Staphylococcus aureus" Antibiotics 11, no. 5: 683. https://doi.org/10.3390/antibiotics11050683
APA StyleLee, H.-J., Kim, B., Kim, S., Cho, D.-H., Jung, H., Kim, W., Kim, Y.-G., Kim, J.-S., Joo, H.-S., Lee, S.-H., & Yang, Y.-H. (2022). Leucyl-tRNA Synthetase Inhibitor, D-Norvaline, in Combination with Oxacillin, Is Effective against Methicillin-Resistant Staphylococcus aureus. Antibiotics, 11(5), 683. https://doi.org/10.3390/antibiotics11050683