
Prevalence, Mechanism, Genetic Diversity, and Cross-Resistance Patterns of Methicillin-Resistant Staphylococcus Isolated from Companion Animal Clinical Samples Submitted to a Veterinary Diagnostic Laboratory in the Midwestern United States

 ,
,
Abstract
1. Introduction
2. Results and Discussion
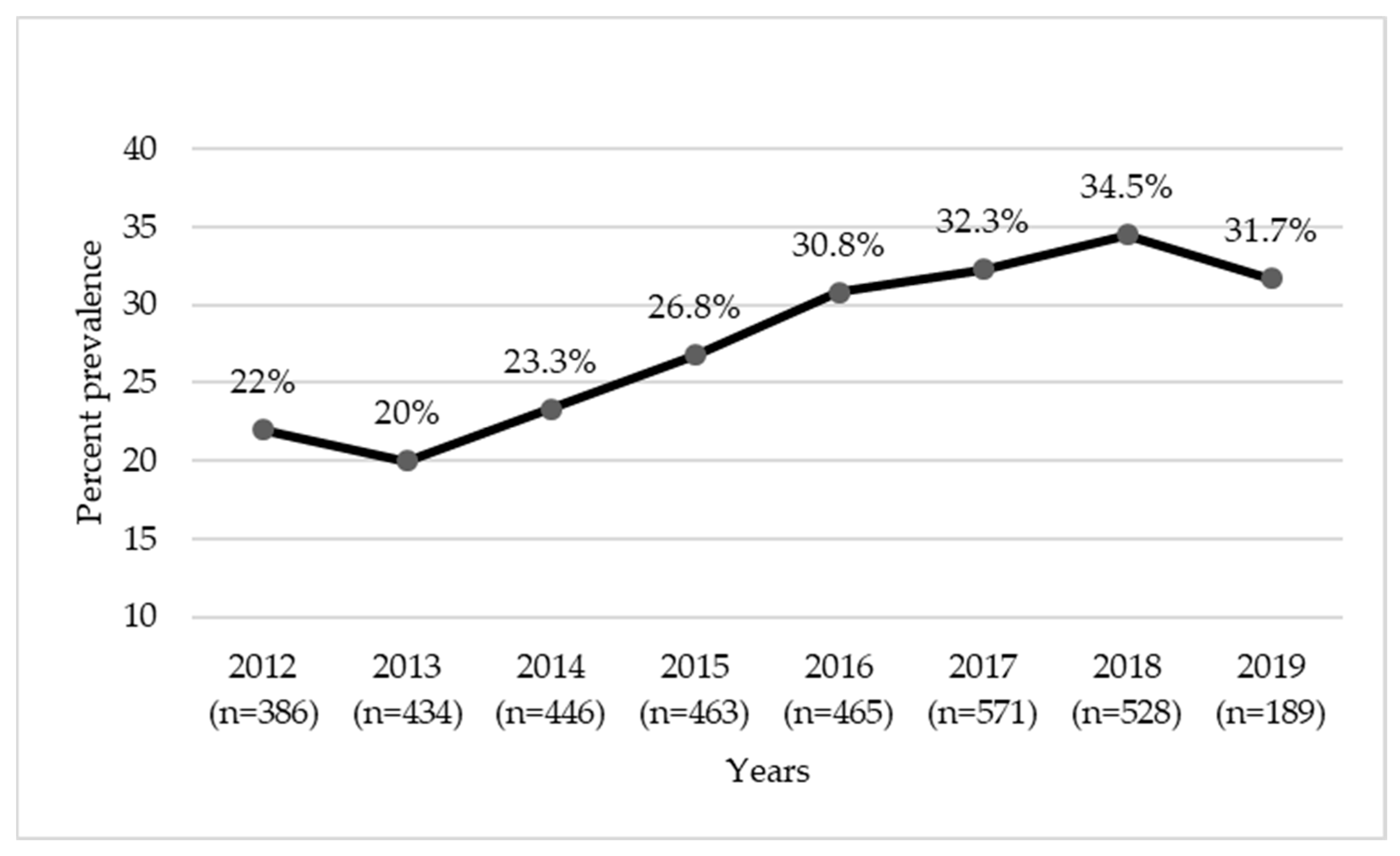
2.1. Prevalence of MRS
2.2. Cross-Resistance Patterns of MRS
2.3. Resistance to Anti-Gram-Positive Antimicrobials in MRS
2.4. Mechanism of Methicillin Resistance in MRS
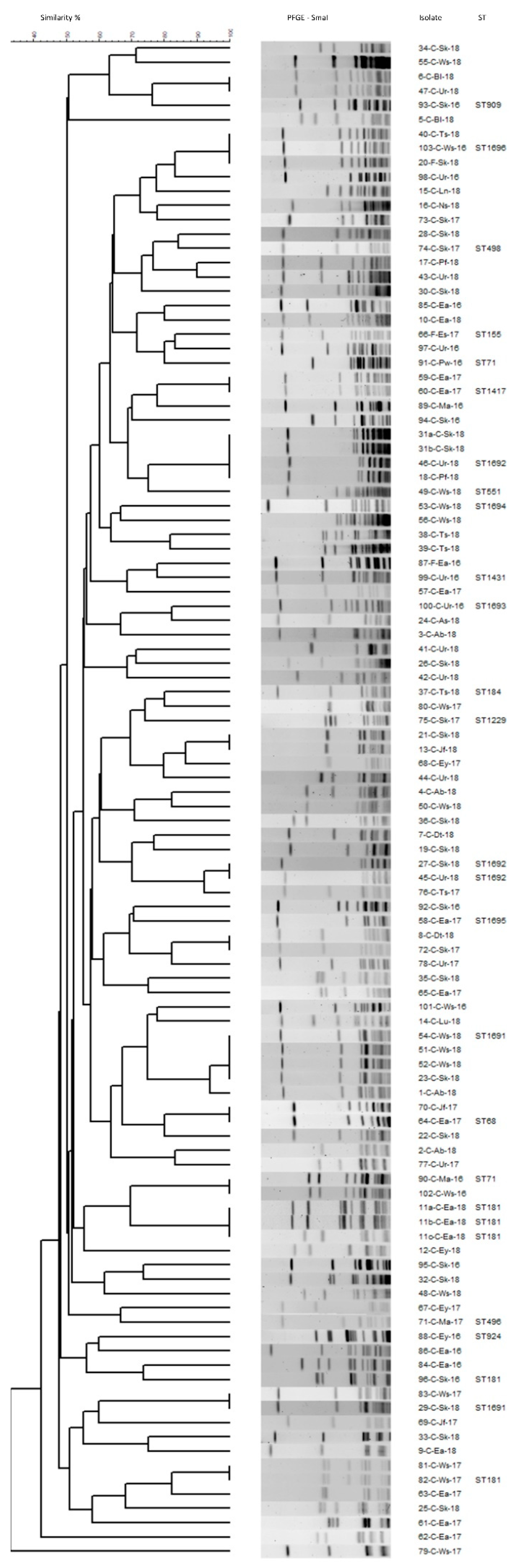
2.5. Genetic Diversity in MRSP
3. Materials and Methods
3.1. Sample Source and Bacterial Isolation and Identification
3.2. Antimicrobial Susceptibility Testing
3.3. Confirmation of Methicillin Resistance
3.4. Molecular Typing of Isolates
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guardabassi, L.; Damborg, P.; Stamm, I.; Kopp, P.A.; Broens, E.M.; Toutain, P.L.; ESCMID Study Group for Veterinary Microbiology. Diagnostic microbiology in veterinary dermatology: Present and future. Vet. Dermatol. 2017, 8, 163–172. [Google Scholar]
- Ruzauskas, M.; Couto, N.; Kerziene, S.; Siugzdiniene, R.; Klimiene, I.; Virgailis, M.; Pomba, C. Prevalence, species distribution and antimicrobial resistance patterns of methicillin-resistant staphylococci in Lithuanian pet animals. Acta Vet. Scand. 2015, 57, 27. [Google Scholar] [CrossRef] [PubMed]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253.e52. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.A.; Helbig, K.J. The Complex diseases of Staphylococcus pseudintermedius in canines: Where to Next? Vet. Sci. 2021, 8, 11. [Google Scholar] [CrossRef]
- Grönthal, T.; Eklund, M.; Thomson, K.; Piiparinen, H.; Sironen, T.; Rantala, M. Antimicrobial resistance in Staphylococcus pseudintermedius and the molecular epidemiology of methicillin-resistant S. pseudintermedius in small animals in Finland. J. Antimicrob. Chemother. 2017, 72, 1021–1030. [Google Scholar] [CrossRef]
- Saputra, S.; Jordan, D.; Worthing, K.A.; Norris, J.M.; Wong, H.S.; Abraham, R.; Trott, D.J.; Abraham, S. Antimicrobial resistance in coagulase-positive staphylococci isolated from companion animals in Australia: A one-year study. PLoS ONE 2017, 12, e0176379. [Google Scholar] [CrossRef]
- Couto, N.; Monchique, C.; Belas, A.; Marques, C.; Gama, L.T.; Pomba, C. Trends and molecular mechanisms of antimicrobial resistance in clinical staphylococci isolated from companion animals over a 16 year period. J. Antimicrob. Chemother. 2016, 71, 1479–1487. [Google Scholar] [CrossRef]
- Joffe, D.; Goulding, F.; Langelier, K.; Magyar, G.; McCurdy, L.; Milstein, M.; Nielsen, K.; Villemaire, S. Prevalence of methicillin-resistant staphylococci in canine pyoderma cases in primary care veterinary practices in Canada: A preliminary study. Can. Vet. J. 2015, 56, 1084–1086. [Google Scholar]
- Kern, A.; Perreten, V. Clinical and molecular features of methicillin-resistant, coagulase-negative staphylococci of pets and horses. J. Antimicrob. Chemother. 2013, 68, 1256–1266. [Google Scholar] [CrossRef]
- Feng, Y.; Tian, W.; Lin, D.; Luo, Q.; Zhou, Y.; Yang, T.; Deng, Y.H.; Liu, Y.H.; Liu, J.H. Prevalence and characterization of methicillin-resistant Staphylococcus pseudintermedius in pets from South China. Vet. Microbiol. 2012, 160, 517–524. [Google Scholar] [CrossRef]
- Perreten, V.; Kadlec, K.; Schwarz, S.; Grönlund Andersson, U.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A.; et al. Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: An international multicentre study. J. Antimicrob. Chemother. 2010, 65, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Nisa, S.; Bercker, C.; Midwinter, A.C.; Bruce, I.; Graham, C.F.; Venter, P.; Bell, N.P.; French, J.; Benschop, K.M.B.; Wilkinson, D.A. Combining MALDI-TOF and genomics in the study of methicillin resistant and multidrug resistant Staphylococcus pseudintermedius in New Zealand. Sci. Rep. 2019, 9, 1271. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.M.; Wu, M.T.; Westblade, L.F.; Robertson, A.E.; Burnham, C.A.; Wallace, M.A.; Burd, E.M.; Lawhon, S.; Hindler, S.A. In vitro antimicrobial susceptibility of Staphylococcus pseudintermedius isolates of human and animal origin. J. Clin. Microbiol. 2016, 54, 1391–1394. [Google Scholar] [CrossRef] [PubMed]
- Somayaji, R.; Priyantha, M.A.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Somayaji, R.; Rubin, J.E.; Priyantha, M.A.; Church, D. Exploring Staphylococcus pseudintermedius: An emerging zoonotic pathogen? Future Microbiol. 2016, 11, 1371–1374. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Ceballos, S.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. Clonally diverse methicillin and multidrug resistant coagulase negative staphylococci are ubiquitous and pose transfer ability between pets and their owners. Front. Microbiol. 2019, 10, 485. [Google Scholar] [CrossRef]
- Little, S.V.; Bryan, L.K.; Hillhouse, A.E.; Cohen, N.D.; Lawhon, S.D. Characterization of agr groups of Staphylococcus pseudintermedius isolates from dogs in Texas. Msphere 2019, 4, 1–11. [Google Scholar] [CrossRef]
- Worthing, K.A.; Abraham, S.; Coombs, G.W.; Pang, S.; Saputra, S.; Jordan, D.; Trott, D.J.; Norris, J.M. Clonal diversity and geographic distribution of methicillin-resistant Staphylococcus pseudintermedius from Australian animals: Discovery of novel sequence types. Vet. Microbiol. 2018, 213, 58–65. [Google Scholar] [CrossRef]
- Lozano, C.; Rezusta, A.; Ferrer, I.; Pérez-Laguna, V.; Zarazaga, M.; Ruiz-Ripa, L.; Revillo, M.J.; Torres, C. Staphylococcus pseudintermedius human infection cases in Spain: Dog-to-human transmission. Vector Borne Zoonotic Dis. 2017, 17, 268–270. [Google Scholar] [CrossRef]
- Dupieux, C.; Trouillet-Assant, S.; Tasse, J.; Freydière, A.M.; Raulin, O.; Roure-Sobas, C.; Salord, H.; Tigaud, S.; Laurent, F. Evaluation of a commercial immunochromatographic assay for rapid routine identification of PBP2a-positive Staphylococcus aureus and coagulase-negative staphylococci. Diagn. Microbiol. Infect. Dis. 2016, 86, 262–264. [Google Scholar] [CrossRef]
- Mantion, B.; Cavalié, L.; Prère, M.F. Evaluation of an immunochromatographic assay for detection of PBP2a on non-Staphylococcus aureus clinical isolates. J. Microbiol. Methods 2015, 112, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Ballhausen, B.; Köck, R.; Kriegeskorte, A. Methicillin resistance in Staphylococcus isolates: The “mec alphabet” with specific consideration of mecC, a mec homolog associated with zoonotic S. aureus lineages. Int. J. Med. Microbiol. 2014, 304, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Peacock, S.J.; Paterson, G.K. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-encoded transferable mecB-mediated methicillin resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242. [Google Scholar] [CrossRef]
- Schwendener, S.; Cotting, K.; Perreten, V. Novel methicillin resistance gene mecD in clinical Macrococcus caseolyticus strains from bovine and canine sources. Sci. Rep. 2017, 7, 43797. [Google Scholar] [CrossRef]
- Dupieux, C.; Bouchiat, C.; Larsen, A.R.; Pichon, B.; Holmes, M.; Teale, C.; Edwards, G.; Hill, R.; Decousser, J.W.; Trouillet-Assant, S.; et al. Detection of mecC-positive Staphylococcus aureus: What to expect from immunological tests targeting PBP2a? J. Clin. Microbiol. 2017, 55, 1961–1963. [Google Scholar] [CrossRef]
- Chen, M.M.; Boardman, W.S.; Brown, M.H. Methicillin resistance gene diversity in staphylococci isolated from captive and free-ranging wallabies. Infect. Ecol. Epidemiol. 2016, 6, 31507. [Google Scholar] [CrossRef][Green Version]
- Paterson, G.K.; Harrison, E.M.; Holmes, M.A. The emergence of mecC methicillin-resistant Staphylococcus aureus. Trends Immunol. 2014, 22, 42–47. [Google Scholar] [CrossRef]
- Stegger, Á.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef]
- Wu, M.T.; Burnham, C.A.; Westblade, L.F.; Bard, J.D.; Lawhon, S.D.; Wallace, M.A.; Stanley, T.; Burd, E.; Hindler, J.; Humphries, R.M. Evaluation of oxacillin and cefoxitin disk and MIC breakpoints for prediction of methicillin resistance in human and veterinary isolates of Staphylococcus intermedius group. J. Clin. Microbiol. 2016, 54, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Delport, J.A.; Mohorovic, I.; Burn, S.; McCormick, J.K.; Schaus, D.; Lannigan, R.; John, M. Rapid detection of meticillin-resistant Staphylococcus aureus bacteraemia using combined three-hour short-incubation matrix-assisted laser desorption/ionization time-of-flight MS identification and alere culture colony PBP2a detection test. J. Med. Microbiol. 2016, 65, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.R.; Burnham, C.A.; Ford, B.A.; Lawhon, S.D.; McAllister, S.K.; Lonsway, D.; Albrecht, V.; Jerris, R.C.; Rasheed, J.K.; Limbago, B.; et al. Evaluation of an immunochromatographic assay for rapid detection of penicillin-binding protein 2a in human and animal Staphylococcus intermedius group, Staphylococcus lugdunensis, and Staphylococcus schleiferi clinical isolates. J. Clin. Microbiol. 2016, 54, 745–748. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Videla, R.; Solyman, S.M.; Brahmbhatt, A.; Sadeghi, L.; Bemis, D.A.; Kania, S.A. Clonal complexes and antimicrobial susceptibility profiles of Staphylococcus pseudintermedius isolates from dogs in the United States. Microb. Drug Resist. 2018, 24, 83–88. [Google Scholar] [CrossRef]
- Duim, B.; Verstappen, K.M.; Broens, E.M.; Laarhoven, L.M.; Van Duijkeren, E.; Hordijk, J.; de Heus, P.; Spaninks, M.; Timmerman, A.J.; Wagenaar, J.A. Changes in the population of methicillin-resistant Staphylococcus pseudintermedius and dissemination of antimicrobial-resistant phenotypes in the Netherlands. J. Clin. Microbiol. 2016, 54, 283–288. [Google Scholar] [CrossRef]
- Lord, J.; Millis, N.; Jones, R.D.; Johnson, B.; Kania, S.A. Patterns of antimicrobial, multidrug and methicillin resistance among Staphylococcus spp. isolated from canine specimens submitted to a diagnostic laboratory in Tennessee, USA: A descriptive study. BMC Vet. Res. 2022, 18, 91. [Google Scholar] [CrossRef]
- Soimala, T.; Lübke-Becker, A.; Hanke, D.; Eichhorn, I.; Feßler, A.T.; Schwarz, S.; Eule, J.C. Molecular and phenotypic characterization of methicillin-resistant Staphylococcus pseudintermedius from ocular surfaces of dogs and cats suffering from ophthalmological disease. Vet. Microbiol. 2020, 244, 108687. [Google Scholar] [CrossRef]
- Wegener, A.; Broens, E.M.; Zomer, A.; Spaninks, M.; Wagenaar, J.A.; Duim, B. Comparative genomics of phenotypic antimicrobial resistances in methicillin-resistant Staphylococcus pseudintermedius of canine origin. Vet. Microbiol. 2018, 225, 125–131. [Google Scholar] [CrossRef]
- Loiacono, M.; Martino, P.A.; Albonico, F.; Dell’Orco, F.; Ferretti, M.; Zanzani, S. High-resolution melting analysis of gyrA codon 84 and grlA codon 80 mutations conferring resistance to fluoroquinolones in Staphylococcus pseudintermedius isolates from canine clinical samples. J. Vet. Diagn. Investig. 2017, 29, 711–715. [Google Scholar] [CrossRef]
- Ventrella, G.; Moodley, A.; Grandolfo, E.; Parisi, A.; Corrente, M.; Buonavoglia, D.; Guardabassi, L. Frequency, antimicrobial susceptibility and clonal distribution of methicillin-resistant Staphylococcus pseudintermedius in canine clinical samples submitted to a veterinary diagnostic laboratory in Italy: A 3-year retrospective investigation. Vet. Microbiol. 2017, 211, 103–106. [Google Scholar] [CrossRef]
- Moodley, A.; Stegger, M.; Zakour, N.L.; Fitzgerald, J.R.; Guardabassi, L. Tandem repeat sequence analysis of staphylococcal protein A (spa) gene in methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 2009, 135, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Gold, R.M.; Cohen, N.D.; Lawhon, S.D. Amikacin resistance in Staphylococcus pseudintermedius isolated from dogs. J. Clin. Microbiol. 2014, 25, 3641–3646. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.T.; Amador, S.; McGonagle, C.J.; Needle, D.; Gibson, R.; Andam, C.P. Population genomics of Staphylococcus pseudintermedius in companion animals in the United States. Commun. Biol. 2020, 3, 282. [Google Scholar] [CrossRef] [PubMed]
- Han, J.I.; Rhim, H.; Yang, C.H.; Park, H.M. Molecular characteristics of new clonal complexes of Staphylococcus pseudintermedius from clinically normal dogs. Vet. Q. 2018, 38, 14–20. [Google Scholar] [CrossRef]
- Pires dos Santos, T.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic review on global epidemiology of methicillin-resistant Staphylococcus pseudintermedius: Inference of population structure from multilocus sequence typing data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef]
- Damborg, P.; Moodley, A.; Aalbæk, B.; Ventrella, G.; Dos Santos, T.P.; Guardabassi, L. High genotypic diversity among methicillin-resistant Staphylococcus pseudintermedius isolated from canine infections in Denmark. BMC Vet. Res. 2016, 12, 131. [Google Scholar] [CrossRef][Green Version]
- Kjellman, E.E.; Slettemeås, J.S.; Small, H.; Sunde, M. Methicillin-resistant Staphylococcus pseudintermedius (MRSP) from healthy dogs in Norway–occurrence, genotypes and comparison to clinical MRSP. Microbiologyopen 2015, 4, 857–866. [Google Scholar] [CrossRef]
- Solyman, S.M.; Black, C.C.; Duim, B.; Perreten, V.; Van Duijkeren, E.; Wagenaar, J.A.; Eberlein, L.C.; Sadeghi, L.N.; Videla, R.; Bemis, D.A.; et al. Multilocus sequence typing for characterization of Staphylococcus pseudintermedius. J. Clin. Microbiol. 2013, 51, 306–310. [Google Scholar] [CrossRef]
- Nienhoff, U.; Kadlec, K.; Chaberny, I.F.; Verspohl, J.; Gerlach, G.F.; Kreienbrock, L.; Schwarz, S.; Simon, D.; Nolte, I. Methicillin-resistant Staphylococcus pseudintermedius among dogs admitted to a small animal hospital. Vet. Microbiol. 2011, 150, 191–197. [Google Scholar] [CrossRef]
- Black, C.C.; Solyman, S.M.; Eberlein, L.C.; Bemis, D.A.; Woron, A.M.; Kania, S.A. Identification of a predominant multilocus sequence type, pulsed-field gel electrophoresis cluster, and novel staphylococcal chromosomal cassette in clinical isolates of mecA-containing, methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 2009, 139, 333–338. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Simón, C.; Ceballos, S.; Ortega, C.; Zarazaga, M.; Torres, C.; Gómez-Sanz, E. S. pseudintermedius and S. aureus lineages with transmission ability circulate as causative agents of infections in pets for years. BMC Vet. Res. 2021, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Bergot, M.; Martins-Simoes, P.; Kilian, H.; Châtre, P.; Worthing, K.A.; Norris, J.M.; Madec, J.Y.; Laurent, F.; Haenni, M. Evolution of the population structure of Staphylococcus pseudintermedius in France. Front. Microbiol. 2018, 9, 3055. [Google Scholar] [CrossRef] [PubMed]
- Bruce, S.A.; Smith, J.T.; Mydosh, J.L.; Ball, J.; Needle, D.B.; Gibson, R.; Andam, C.P. Accessory genome dynamics of local and global Staphylococcus pseudintermedius populations. Front. Microbiol. 2022, 13, 798175. [Google Scholar] [CrossRef]
- Marques, C.; Gama, L.T.; Belas, A.; Bergström, K.; Beurlet, S.; Briend-Marchal, A.; Broens, E.M.; Costa, M.; Criel, D.; Damborg, P.; et al. European multicenter study on antimicrobial resistance in bacteria isolated from companion animal urinary tract infections. BMC Vet. Res. 2016, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Eckholm, N.G.; Outerbridge, C.A.; White, S.D.; Sykes, J.E. Prevalence of and risk factors for isolation of meticillin-resistant Staphylococcus spp. from dogs with pyoderma in northern California, USA. Vet. Dermatol. 2013, 24, 154-e34. [Google Scholar]
- Jones, R.D.; Kania, S.A.; Rohrbach, B.W.; Frank, L.A.; Bemis, D.A. Prevalence of oxacillin-and multidrug-resistant staphylococci in clinical samples from dogs: 1772 samples (2001–2005). J. Am. Vet. Med. Assoc. 2007, 230, 221–227. [Google Scholar] [CrossRef]
- Saab, M.E.; Muckle, C.A.; Stryhn, H.; McClure, J.T. Comparison of culture methodology for the detection of methicillin-resistant Staphylococcus pseudintermedius in clinical specimens collected from dogs. J. Vet. Diagn. Investig. 2018, 30, 93–98. [Google Scholar] [CrossRef]
- Ruzauskas, M.; Couto, M.; Pavilonis, A.; Klimiene, I.; Siugzdiniene, R.; Virgailis, M.; Vaskeviciute, L.; Anskiene, L. Characterization of Staphylococcus pseudintermedius isolated from diseased dogs in Lithuania. Pol. J. Vet. Sci. 2016, 19, 7–14. [Google Scholar] [CrossRef][Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Supplement VET08: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI Supplement M100: Wayne, PA, USA, 2017. [Google Scholar]
- Leclercq, R.; Cantón, R.; Brown, D.F.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef]
- Mendes, R.E.; Sader, H.S.; Farrell, D.J.; Jones, R.N. Telavancin activity tested against a contemporary collection of Gram-positive pathogens from USA hospitals (2007–2009). Diagn. Microbiol. Infect. Dis. 2012, 72, 113–117. [Google Scholar] [CrossRef]
- Shariati, A.; Dadashi, M.; Chegini, Z.; van Belkum, A.; Mirzaii, M.; Khoramrooz, S.S.; Darban-Sarokhalil, D. The global prevalence of Daptomycin, Tigecycline, Quinupristin/Dalfopristin, and Linezolid-resistant Staphylococcus aureus and coagulase negative staphylococci strains: A systematic review and meta-analysis. Antimicrob. Resist. Infect. Control. 2020, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Feßler, A.T.; Scholtzek, A.D.; Schug, A.R.; Kohn, B.; Weingart, C.; Schink, A.K.; Bethe, A.; Lübke-Becker, A.; Schwarz, S. Antimicrobial and biocide resistance among feline and canine Staphylococcus aureus and Staphylococcus pseudintermedius isolates from diagnostic submissions. Antibiotics 2022, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; Van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M.; et al. Public health risk of antimicrobial resistance transfer from companion animals. J. Antimicrob. Chemother. 2017, 72, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Bressler, A.M.; Williams, T.; Culler, E.E.; Zhu, W.; Lonsway, D.; Patel, J.B.; Nolte, F.S. Correlation of penicillin binding protein 2a detection with oxacillin resistance in Staphylococcus aureus and discovery of a novel penicillin binding protein 2a mutation. J. Clin. Microbiol. 2005, 43, 4541–4544. [Google Scholar] [CrossRef][Green Version]
- Argudín, M.A.; Roisin, S.; Nienhaus, L.; Dodémont, M.; De Mendonça, R.; Nonhoff, C.; Deplano, A.; Denis, O. Genetic diversity among Staphylococcus aureus isolates showing oxacillin and/or cefoxitin resistance not linked to the presence of mec genes. Antimicrob. Agents Chemother. 2018, 62, e00091-18. [Google Scholar] [CrossRef]
- Perkins, A.V.; Sellon, D.C.; Gay, J.M.; Lofgren, E.T.; Moore, D.A.; Jones, L.P.; Davis, M.A. Prevalence of methicillin-resistant Staphylococcus pseudintermedius on hand-contact and animal-contact surfaces in companion animal community hospitals. Can. Vet. J. 2020, 61, 613–620. [Google Scholar]
- Van Duijkeren, E.; Kamphuis, M.; Van der Mije, I.C.; Laarhoven, L.M.; Duim, B.; Wagenaar, J.A.; Houwers, D.J. Transmission of methicillin-resistant Staphylococcus pseudintermedius between infected dogs and cats and contact pets, humans and the environment in households and veterinary clinics. Vet. Microbiol. 2011, 150, 338–343. [Google Scholar] [CrossRef]
- Kronbichler, A.; Blane, B.; Holmes, M.A.; Wagner, J.; Parkhill, J.; Peacock, S.J.; Jayne, D.R.W.; Harrison, E.M. Nasal carriage of Staphylococcus pseudintermedius in patients with granulomatosis with polyangiitis. Rheumatology 2019, 58, 548–550. [Google Scholar] [CrossRef]
- Phumthanakorn, N.; Fungwithaya, P.; Chanchaithong, P.; Prapasarakul, N. Enterotoxin gene profile of methicillin-resistant Staphylococcus pseudintermedius isolates from dogs, humans and the environment. J. Med. Microbiol. 2018, 67, 866–873. [Google Scholar] [CrossRef]
- Stegmann, R.; Burnens, A.; Maranta, C.A.; Perreten, V. Human infection associated with methicillin-resistant Staphylococcus pseudintermedius ST71. J. Antimicrob. Chemother. 2010, 65, 2047–2048. [Google Scholar] [CrossRef]
- Scott, B.A.; Mortensen, J.E.; McKeever, T.M.; Logas, D.B.; McKeever, P.J. Efficacy of tylosin tartrate on canine Staphylococcus intermedius isolates in vitro. Vet. Ther. Res. Appl. Vet. Med. 2010, 11, E1–E7. [Google Scholar]
- Bahraminia, F.; Emadi, S.R.; Emaneini, M.; Farzaneh, N.; Rad, M.; Khoramian, B. A high prevalence of tylosin resistance among Staphylococcus aureus strains isolated from bovine mastitis. Vet. Res. Forum. 2017, 8, 121–125. [Google Scholar] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing EUCAST. Breakpoint tables for interpretation of MICs and zone diameters. EUCAST 2019, 9, 10–15. [Google Scholar]
- Nicolau, D.P.; Quintana, A.; Korth-Bradley, J.M.; Wible, M.; Dowzicky, M.J. A rationale for maintaining current tigecycline breakpoints as established by the USA food and drug administration. Arch. Clin. Microbiol. 2015, 6, 1–12. [Google Scholar]
- Kohner, P.; Uhl, J.; Kolbert, C.; Persing, D.; Cockerill, F. Comparison of susceptibility testing methods with mecA gene analysis for determining oxacillin (methicillin) resistance in clinical isolates of Staphylococcus aureus and coagulase-negative Staphylococcus spp. J. Clin. Microbiol. 1999, 37, 2952–2961. [Google Scholar] [CrossRef]
- Goering, R.V. Pulsed field gel electrophoresis: A review of application and interpretation in the molecular epidemiology of infectious disease. Infect. Genet. Evol. 2010, 10, 866–875. [Google Scholar] [CrossRef]
- André, M.C.; Campos, M.R.; Borges, L.J.; Kipnis, A.; Pimenta, F.C.; Serafini, A.B. Comparison of Staphylococcus aureus isolates from food handlers, raw bovine milk and Minas Frescal cheese by antibiogram and pulsed-field gel electrophoresis following SmaI digestion. Food. Control 2008, 19, 200–207. [Google Scholar] [CrossRef]
- Mulvey, M.R.; Chui, L.; Ismail, J.; Louie, L.; Murphy, C.; Chang, N.; Alfa, M.; Canadian Committee for the Standardization of Molecular Methods. Development of a Canadian standardized protocol for subtyping methicillin-resistant Staphylococcus aureus using pulsed-field gel electrophoresis. J. Clin. Microbiol. 2001, 39, 3481–3485. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Class | Antibiotic | Oxa MIC Breakpoint (µg/mL) * | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| <0.5 (S) | ≥0.5–<4 (R) | ≥4 (R-SA) | |||||||||||
| # | %NS | MIC50 | MIC90 | # | %NS | MIC50 | MIC90 | # | %NS | MIC50 | MIC90 | ||
| β-lactam | Ampicillin A | 1523 | 30.5 | ≤0.25 | 1 | 243 | 90.9 | >1 | >1 | 188 | 98.4 | >1 | >1 |
| Cefazolin A | 1524 | 0.3 | ≤2 | ≤2 | 243 | 1.6 | ≤2 | ≤2 | 188 | 57.4 | 4 | >4 | |
| Cefovecin A | 1524 | 2.0 | ≤0.5 | ≤0.5 | 243 | 86.0 | 2 | >4 | 188 | 98.9 | >4 | >4 | |
| Cefoxitin B | 1168 | 0.3 | ≤2 | ≤2 | 140 | 0.7 | ≤2 | ≤2 | 137 | 11.7 | ≤2 | 8 | |
| Cefpodoxime A | 1524 | 0.5 | ≤2 | ≤2 | 243 | 30.0 | ≤2 | 8 | 188 | 94.7 | >8 | >8 | |
| Ceftiofur C | 1168 | NA | ≤0.25 | ≤0.25 | 140 | NA | 1 | 4 | 137 | NA | >4 | >4 | |
| Cephalotin A | 1524 | 0.4 | ≤2 | ≤2 | 243 | 2.1 | ≤2 | ≤2 | 188 | 45.7 | ≤2 | >4 | |
| Imipenem C | 1524 | NA | ≤1 | ≤1 | 243 | NA | ≤1 | ≤1 | 188 | NA | ≤1 | ≤1 | |
| Penicillin A | 1524 | 51.8 | 0.5 | 8 | 243 | 93.4 | >8 | >8 | 188 | 98.9 | >8 | >8 | |
| Aminoglycoside | Amikacin A | 1168 | 0.7 | ≤4 | ≤4 | 141 | 1.4 | ≤4 | ≤4 | 139 | 5.0 | ≤4 | 8 |
| Gentamicin B | 1524 | 13.1 | ≤4 | 8 | 243 | 42.4 | ≤4 | >8 | 188 | 63.8 | 8 | >8 | |
| Fluoroquinolone | Enrofloxacin A | 1523 | 12.1 | ≤0.25 | 1 | 243 | 65.4 | >2 | >2 | 188 | 67.0 | >2 | >2 |
| MarbofloxacinA | 1524 | 6.5 | ≤1 | ≤1 | 243 | 56.4 | >2 | >2 | 188 | 57.4 | >2 | >2 | |
| PradofloxacinA | 356 | 8.1 | ≤0.25 | ≤0.25 | 103 | 61.2 | 2 | >2 | 51 | 72.5 | 2 | >2 | |
| Tetracycline | Doxycycline A | 768 | 68.8 | >0.5 | >0.5 | 195 | 88.2 | >0.5 | >0.5 | 132 | 91.7 | >0.5 | >0.5 |
| Minocycline C | 356 | NA | ≤0.5 | >2 | 103 | NA | >2 | >2 | 51 | NA | >2 | >2 | |
| Tetracycline A | 356 | 33.7 | ≤0.25 | >1 | 103 | 78.6 | >1 | >1 | 51 | 80.4 | >1 | >1 | |
| Lincosamide | Clindamycin A | 1524 | 13.3 | ≤0.5 | >4 | 243 | 72.8 | >4 | >4 | 188 | 71.8 | >4 | >4 |
| Macrolide | Erythromycin B | 1524 | 14.5 | ≤0.5 | >4 | 243 | 73.3 | >4 | >4 | 188 | 75.0 | >4 | >4 |
| Phenicol | Chloramphenicol B | 1524 | 8.5 | ≤8 | ≤8 | 243 | 27.6 | ≤8 | >16 | 188 | 37.8 | ≤8 | >16 |
| Nitrofurantion B | 356 | 0.0 | ≤16 | ≤16 | 103 | 0.0 | ≤16 | ≤16 | 51 | 2.0 | ≤16 | ≤16 | |
| Trimethoprim/Sulfamethoxazole B | 1524 | 12.1 | ≤2 | >2 | 243 | 67.9 | >2 | >2 | 188 | 70.7 | >2 | >2 | |
| Antibiotic Class | Antibiotic | Oxa MIC Breakpoint (µg/mL) * | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| <0.5 (S) | ≥0.5–<4 (R) | ≥4 (R-SA) | |||||||||||
| # | %NS | MIC50 | MIC90 | # | %NS | MIC50 | MIC90 | # | %NS | MIC50 | MIC90 | ||
| β-lactam | Ampicillin A | 450 | 5.3 | ≤0.25 | ≤0.25 | 204 | 46.6 | 0.25 | >1 | 103 | 84.5 | >1 | >1 |
| Cefazolin A | 450 | 0.2 | ≤2 | ≤2 | 204 | 4.9 | ≤2 | ≤2 | 101 | 60.4 | 4 | >4 | |
| Cefovecin A | 450 | 6.4 | ≤2 | ≤2 | 204 | 71.6 | 1 | 4 | 99 | 97.0 | >4 | >4 | |
| Cefoxitin B | 353 | 0.9 | ≤2 | ≤2 | 154 | 22.1 | 4 | 8 | 78 | 71.8 | 16 | >16 | |
| Cefpodoxime A | 450 | 2.4 | ≤2 | ≤2 | 204 | 38.2 | ≤2 | 4 | 99 | 94.9 | >8 | >8 | |
| Ceftiofur C | 353 | NA | ≤0.25 | 1 | 154 | NA | 1 | 2 | 82 | NA | >4 | >4 | |
| Cephalotin A | 450 | 0.2 | ≤2 | ≤2 | 204 | 2.9 | ≤2 | ≤2 | 99 | 29.3 | ≤2 | >4 | |
| Imipenem C | 450 | NA | ≤1 | ≤1 | 204 | NA | ≤1 | ≤1 | 103 | NA | ≤1 | 2 | |
| Penicillin A | 450 | 12.0 | ≤0.06 | 0.5 | 204 | 47.5 | 0.25 | 4 | 103 | 92.2 | 4 | >8 | |
| Aminoglycoside | Amikacin A | 353 | 1.4 | ≤4 | ≤4 | 155 | 0.6 | ≤4 | ≤4 | 85 | 18.8 | ≤4 | >32 |
| Gentamicin B | 450 | 2.2 | ≤4 | ≤4 | 204 | 8.8 | ≤4 | ≤4 | 103 | 40.0 | ≤4 | >8 | |
| Fluoroquinolone | Enrofloxacin A | 450 | 9.3 | ≤0.25 | 0.5 | 204 | 31.9 | ≤0.25 | >2 | 103 | 51.5 | 1 | >2 |
| Marbofloxacin B | 450 | 7.3 | ≤1 | ≤1 | 204 | 30.4 | ≤1 | >2 | 99 | 45.5 | ≤1 | >2 | |
| Pradofloxacin A | 97 | 3.1 | ≤0.25 | ≤0.25 | 50 | 38.0 | ≤0.25 | 2 | 21 | 28.6 | ≤0.25 | 1 | |
| Tetracycline | Doxycycline A | 130 | 49.2 | ≤0.12 | >0.5 | 75 | 70.7 | >0.5 | >0.5 | 38 | 92.1 | >0.5 | >0.5 |
| Minocycline C | 97 | NA | ≤0.5 | ≤0.5 | 50 | NA | ≤0.5 | ≤0.5 | 21 | NA | ≤0.5 | ≤0.5 | |
| Tetracycline A | 97 | 32.0 | ≤0.25 | >1 | 50 | 66.0 | 0.5 | >1 | 23 | 91.3 | >1 | >1 | |
| Lincosamide | Clindamycin A | 450 | 13.6 | ≤0.5 | 2 | 204 | 38.7 | ≤0.5 | >4 | 99 | 45.4 | ≤0.5 | >4 |
| Macrolide | Erythromycin B | 450 | 24.7 | ≤0.5 | >4 | 204 | 49.5 | ≤0.5 | >4 | 103 | 49.5 | ≤0.5 | >4 |
| Phenicol | Chloramphenicol B | 450 | 3.1 | ≤8 | ≤8 | 204 | 2.4 | ≤8 | ≤8 | 103 | 14.6 | ≤8 | >16 |
| Nitrofurantion B | 97 | 3.1 | ≤16 | ≤16 | 50 | 2.0 | ≤16 | ≤16 | 21 | 14.3 | ≤16 | 64 | |
| Trimethoprim/Sulfamethoxazole B | 450 | 5.3 | ≤2 | ≤2 | 204 | 19.1 | ≤2 | >2 | 103 | 28.1 | ≤2 | >2 | |
| Antibiotic | No. of Isolates with MIC (µg/mL) a | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | ≥128 | MIC50 | MIC90 | %NS | |
| TGC | 0 | 60 | 85 | 71 | 19 |  1 1 | 0.12 | 0.5 | 0.4 | |||||||
| DAP | 196 | 22 | 13 |  3 3 | 0 | 0 | 0 | 2 | 0.25 | 0.5 | 2.1 | |||||
| LZD | 14 | 155 | 54 | 13 | 0 | 1 | 2 | 0.0 | ||||||||
| VAN | 5 | 66 | 140 | 25 | 0 | 0 | 0 | 0 | 1 | 2 | 0.0 | |||||
| QUI/DAL | 205 | 22 | 9 | 0 | 0 | 0 | 0 | 0.5 | 1 | 3.8 | ||||||
| PEN | 38 | 10 | 13 | 10 | 17 | 10 | 33 | 105 | ≥16 | ≥16 | 83.9 | |||||
| CHL | 0 | 35 | 146 | 25 | 2 | 28 | 8 | ≥32 | 23.3 | |||||||
| NIT | 1 | 8 | 103 | 117 | 5 | 2 | 8 | 16 | 0.8 | |||||||
| EYR | 52 | 35 | 1 | 0 | 1 | 4 | 143 | ≥8 | ≥8 | 63.1 | ||||||
| CIP | 11 | 59 | 31 | 20 | 4 | 3 | 108 | 1 | ≥4 | 57.2 | ||||||
| TYL | 0 | 36 | 64 | 22 | 3 | 2 | 0 | 109 | 1 | ≥32 | 46.2 | |||||
| TET | 101 | 5 | 7 | 0 | 1 | 9 | 113 | 32 | >32 | 100 | ||||||
| Group///Isolate | Oxa MIC | PBP2a | mecA | mecB | mecC | mecD |
|---|---|---|---|---|---|---|
| PBP2a-; mec- CoNS | 0.5 | − | − | − | − | − |
| S. equorum | 0.5 | − | − | − | − | − |
| S. nepalensis | 0.5 | − | − | − | − | − |
| S. pasteuri | 0.5 | − | − | − | − | − |
| S. vitulinus | 0.5 | − | − | − | − | − |
| S. warneri | 0.5 | − | − | − | − | − |
| S. warneri | 0.5 | − | − | − | − | − |
| S. xylosus | 0.5 | − | − | − | − | − |
| S. xylosus | 0.5 | − | − | − | − | − |
| S. xylosus | 0.5 | − | − | − | − | − |
| S. pettenkoferi | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 1 | − | − | − | − | − |
| S. sciuri | 2 | − | − | − | − | − |
| S. sciuri | 2 | − | − | − | − | − |
| S. aureus | 4 | − | − | − | − | − |
| PBP2a-; mec+ S. haemolyticus | 0.5 | − | + | NT | NT | NT |
| S. haemolyticus | >4 | − | + | NT | NT | NT |
| S. epidermidis | 1 | − | + | NT | NT | NT |
| S. epidermidis | 2 | − | + | NT | NT | NT |
| S. epidermidis | 2 | − | + | NT | NT | NT |
| S. epidermidis | 2 | − | + | NT | NT | NT |
| S. epidermidis | 4 | − | + | NT | NT | NT |
| S. epidermidis | >4 | − | + | NT | NT | NT |
| S. epidermidis | >4 | − | + | NT | NT | NT |
| S. epidermidis | >4 | − | + | NT | NT | NT |
| S. warneri | >4 | − | + | NT | NT | NT |
| PBP2a+; mec+ S. simulans | 0.5 | + | + | NT | NT | NT |
| S. epidermidis | 4 | + | + | NT | NT | NT |
| S. hominis | >4 | + | + | NT | NT | NT |
| S. sciuri | >4 | + | + | NT | NT | NT |
| PBP2a+; mec- S. sciuri | 1 | + * | − | − | − | − |
| Indistinguishable Pattern (P) | Same Cluster (C) (≥90% Similarity) | ST | Year of Isolation | Remarks |
|---|---|---|---|---|
| P1 | ||||
| 40-C-Ts-18 | ND | 2018 | Isolates in this pattern were epidemiologically unrelated | |
| 103-C-Ws-16 | 1696 | 2016 | ||
| 20-F-Sk-18 | ND | 2018 | ||
| C1 | ||||
| 17-C-Pf-18 | ND | 2018 | Epidemiologically unrelated | |
| 43-C-Ur-18 | ND | 2018 | ||
| P2 | ||||
| 59-C-Ea-17 | ND | 2017 | Related by location of owners (same town), rDVM (same), time of submissions (a week apart) | |
| 60-C-Ea-17 | 1417 | 2017 | ||
| P3 | ||||
| 31a-C-Sk-18 | ND | 2018 | The first two are from the same animal (different sites at the same time); last two are from the same animal/different sites at the same time; first two and last two are unrelated, but treated in the same hospital two months apart | |
| 31b-C-Sk-18 | ND | 2018 | ||
| 46-C-Ur-18 | 1692 | 2018 | ||
| 18-C-Pf-18 | ND | 2018 | ||
| P4 | ||||
| 54-C-Ws-18 | 1691 | 2018 | The first two are from the same animal/site (two months apart); last two unrelated to each other and the first two but the last three were treated in the same hospital within 2 weeks | |
| 51-C-Ws-18 | ND | 2018 | ||
| 52-C-Ws-18 | ND | 2018 | ||
| 23-C-Sk-18 | ND | 2018 | ||
| C2 | ||||
| 1-C-Ab-18 This isolate forms a cluster with P4 | ND | 2018 | Epidemiologically unrelated to isolates in P4 | |
| P5 | ||||
| 70-C-Jf-17 | ND | Epidemiologically unrelated | ||
| 64-C-Ea-17 | 68 | |||
| P6 | ||||
| 27-C-Sk-18 | 1692 | 2018 | Epidemiologically unrelated | |
| 45-C-Ur-18 | 1692 | 2018 | ||
| C3 | ||||
| 76-C-Ts-17 This isolate forms a cluster P6 | ND | 2017 | Epidemiologically unrelated to isolates in P6 | |
| P7 | ||||
| 21-C-Sk-18 | ND | 2018 | Treated in the same hospital within the same week | |
| 13-C-Jf-18 | ND | 2018 | ||
| P8 | ||||
| 8-C-Dt-18 | ND | 2018 | Epidemiologically unrelated | |
| 72-C-Sk-17 | ND | 2017 | ||
| P9 | ||||
| 83-C-Ws-17 | ND | 2017 | Epidemiologically unrelated | |
| 29-C-Sk-18 | 1691 | 2018 | ||
| P10 | ||||
| 81-C-Ws-17 | ND | 2017 | Treated in the same hospital within the same week | |
| 82-C-WS-17 | 181 | 2017 | ||
| P11 | ||||
| 6-C-Bl-18 | ND | 2018 | From the same animal, a few days apart, bladder stone and urine. | |
| 47-C-Ur-18 | ND | 2018 | ||
| P12 | ||||
| 11a-C-Ea-18 | 181 | 2018 | Same animal from different ears at the same time | |
| 11b-C-Ea-18 | 181 | 2018 | ||
| 11c-C-Ea-18 | 181 | 2018 | ||
| P13 | ||||
| 90-C-Ma-16 | 71 | 2016 | Treated in the same hospital within the same month |
| Antibiotics | Breakpoints | |||
|---|---|---|---|---|
| S | I | R | Reference | |
| Amikacin | ≤4 | ≥16 | [59] | |
| Ampicillin | ≤0.25 | ≥0.5 | [59] | |
| Cefazolin | ≤2 | 4 | ≥8 | [59] |
| Cefoxitin | ≤4 | ≥8 | [59] | |
| Cefovecin | ≤0.5 | 1 | ≥2 | [59] |
| Cefpodoxime | ≤2 | 4 | ≥8 | [59] |
| Cephalothin | ≤2 | 4 | ≥8 | [59] |
| Chloramphenicol | ≤8 | 16 | ≥32 | [59] |
| Ciprofloxacin | ≤0.5 | 1–2 | ≥4 | [59,75] |
| Clindamycin | ≤0.5 | 1–2 | ≥4 | [59] |
| Daptomycin | ≤1 | [60] | ||
| Doxycycline | ≤0.12 | 0.25 | ≥0.5 | [59] |
| Enrofloxacin | ≤0.5 | 2 | ≥4 | [59,75] |
| Erythromycin | ≤0.5 | 1–4 | ≥8 | [60] |
| Gentamicin | ≤4 | 8 | ≥16 | [59] |
| Kanamycin | ≥64 | [59] | ||
| Lincomycin | ≤0.5 | 1–2 | ≥4 | [59] |
| Linezolid | ≤4 | ≥8 | [60] | |
| Marbofloxacin | ≤1 | 2 | ≥4 | [59] |
| Nitrofurantoin | ≤32 | 64 | ≥128 | [60] |
| Penicillin | ≤0.25 | ≥0.5 | [59] | |
| Pradofloxacin | ≤0.25 | 0.5–1 | ≥2 | [59] |
| Quinupristin/Dalfopristin | ≤1 | 2 | ≥4 | [60] |
| Tetracycline | ≤0.25 | 0.5 | ≥1 | [59] |
| Tigecycline | ≤0.5 | ≥1 | [75,76] | |
| Trimethoprim/Sulphamethoxazole | ≤2/38 | ≥4/76 | [59] | |
| Tylosin | ≥32 | [73,74] | ||
| Vancomycin | ≤2 (≤4) | 4–8 (8–16) | ≥16 (≥32) | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adiguzel, M.C.; Schaefer, K.; Rodriguez, T.; Ortiz, J.; Sahin, O. Prevalence, Mechanism, Genetic Diversity, and Cross-Resistance Patterns of Methicillin-Resistant Staphylococcus Isolated from Companion Animal Clinical Samples Submitted to a Veterinary Diagnostic Laboratory in the Midwestern United States. Antibiotics 2022, 11, 609. https://doi.org/10.3390/antibiotics11050609
Adiguzel MC, Schaefer K, Rodriguez T, Ortiz J, Sahin O. Prevalence, Mechanism, Genetic Diversity, and Cross-Resistance Patterns of Methicillin-Resistant Staphylococcus Isolated from Companion Animal Clinical Samples Submitted to a Veterinary Diagnostic Laboratory in the Midwestern United States. Antibiotics. 2022; 11(5):609. https://doi.org/10.3390/antibiotics11050609
Chicago/Turabian StyleAdiguzel, Mehmet Cemal, Kayla Schaefer, Trevor Rodriguez, Jessica Ortiz, and Orhan Sahin. 2022. "Prevalence, Mechanism, Genetic Diversity, and Cross-Resistance Patterns of Methicillin-Resistant Staphylococcus Isolated from Companion Animal Clinical Samples Submitted to a Veterinary Diagnostic Laboratory in the Midwestern United States" Antibiotics 11, no. 5: 609. https://doi.org/10.3390/antibiotics11050609
APA StyleAdiguzel, M. C., Schaefer, K., Rodriguez, T., Ortiz, J., & Sahin, O. (2022). Prevalence, Mechanism, Genetic Diversity, and Cross-Resistance Patterns of Methicillin-Resistant Staphylococcus Isolated from Companion Animal Clinical Samples Submitted to a Veterinary Diagnostic Laboratory in the Midwestern United States. Antibiotics, 11(5), 609. https://doi.org/10.3390/antibiotics11050609