
Multifractal Analysis to Determine the Effect of Surface Topography on the Distribution, Density, Dispersion and Clustering of Differently Organised Coccal-Shaped Bacteria

 ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Surface Fabrication
2.2. Surface Characterisation
2.3. Preparation of Microorganisms
2.4. Retention Assays
2.5. Scanning Electron Microscopy
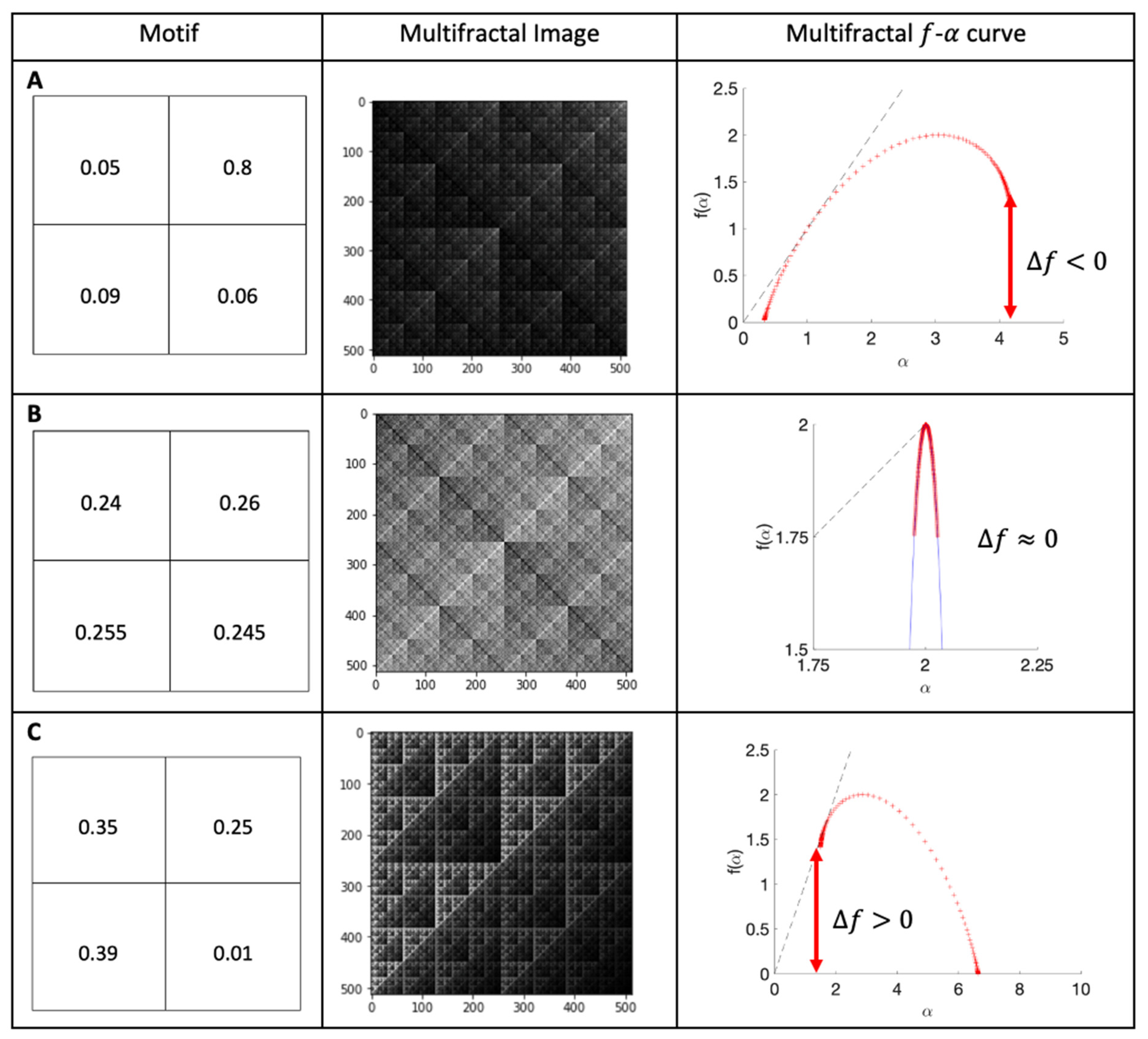
2.6. Multifractal Analysis
2.7. Microbial Adhesion to Hydrocarbons (MATH)
2.8. Statistical Analysis
3. Results
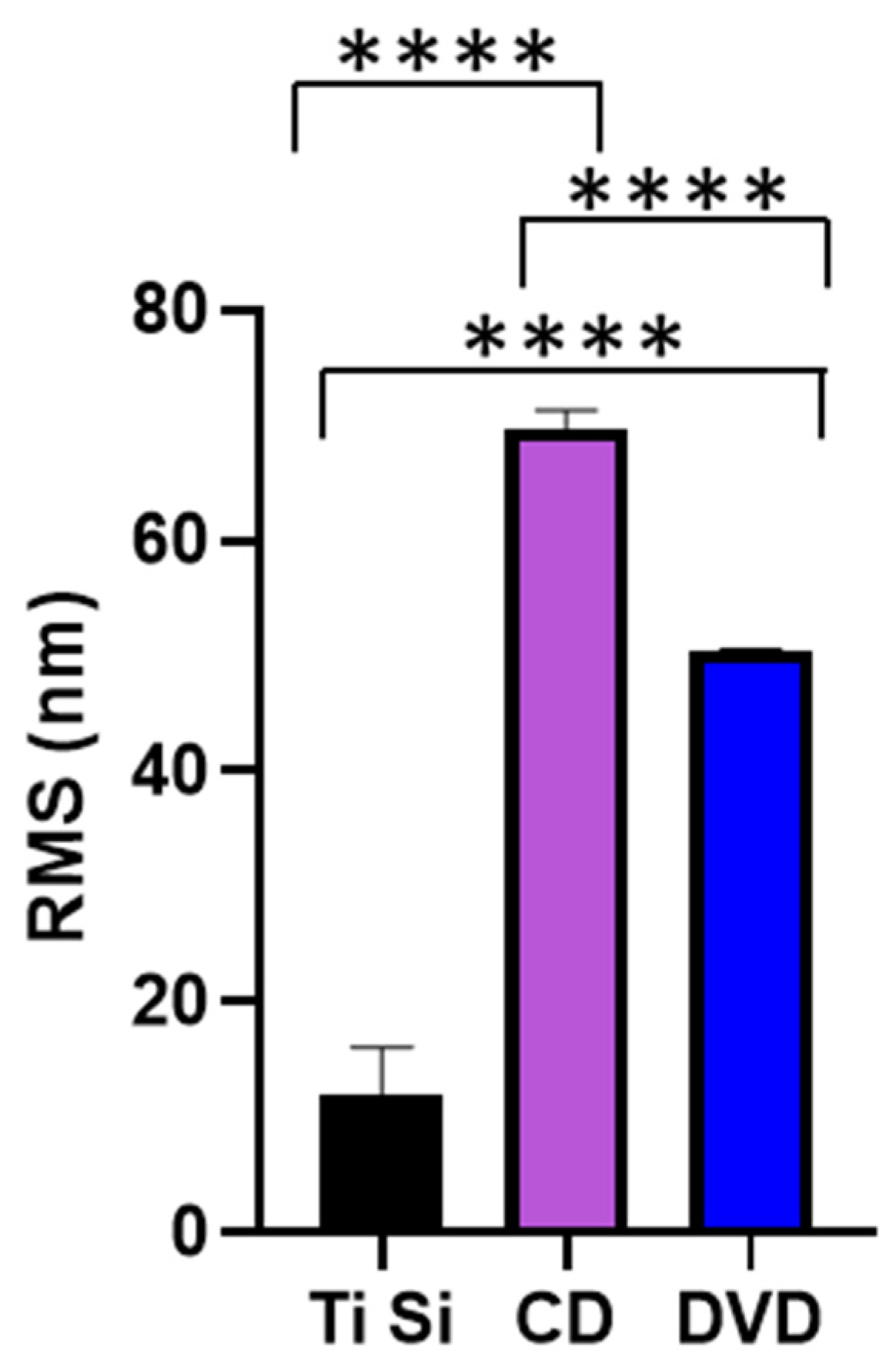
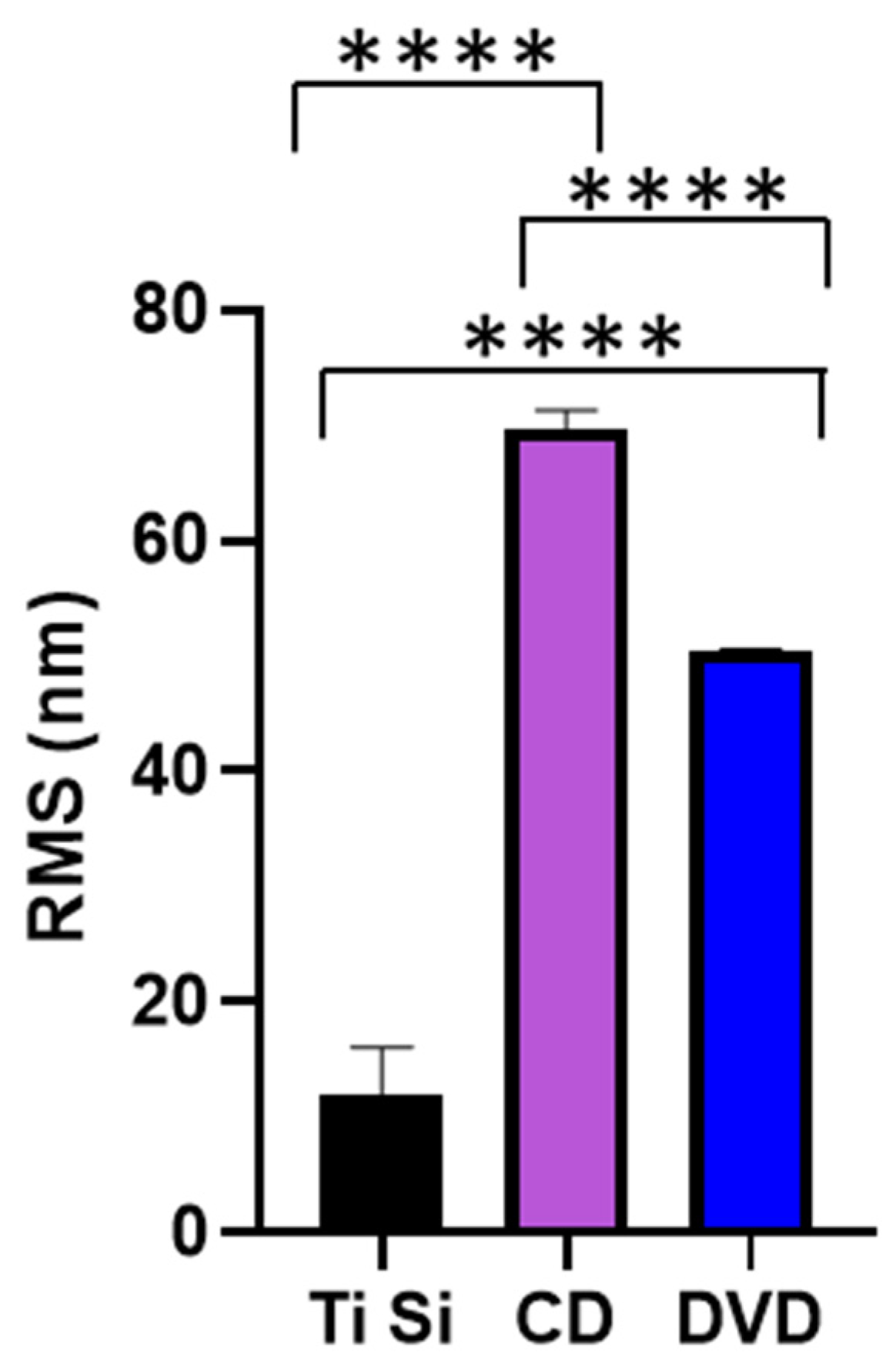
3.1. Surface Characterisation
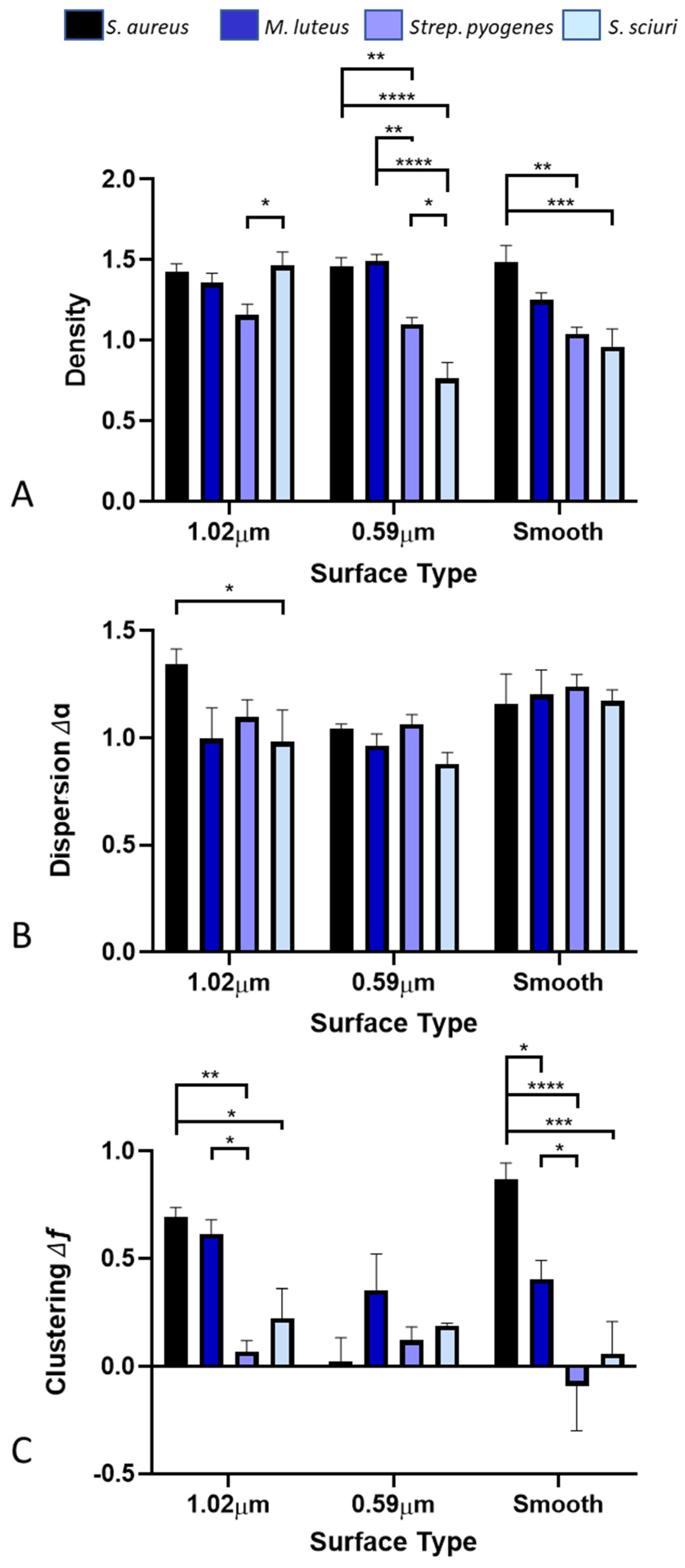
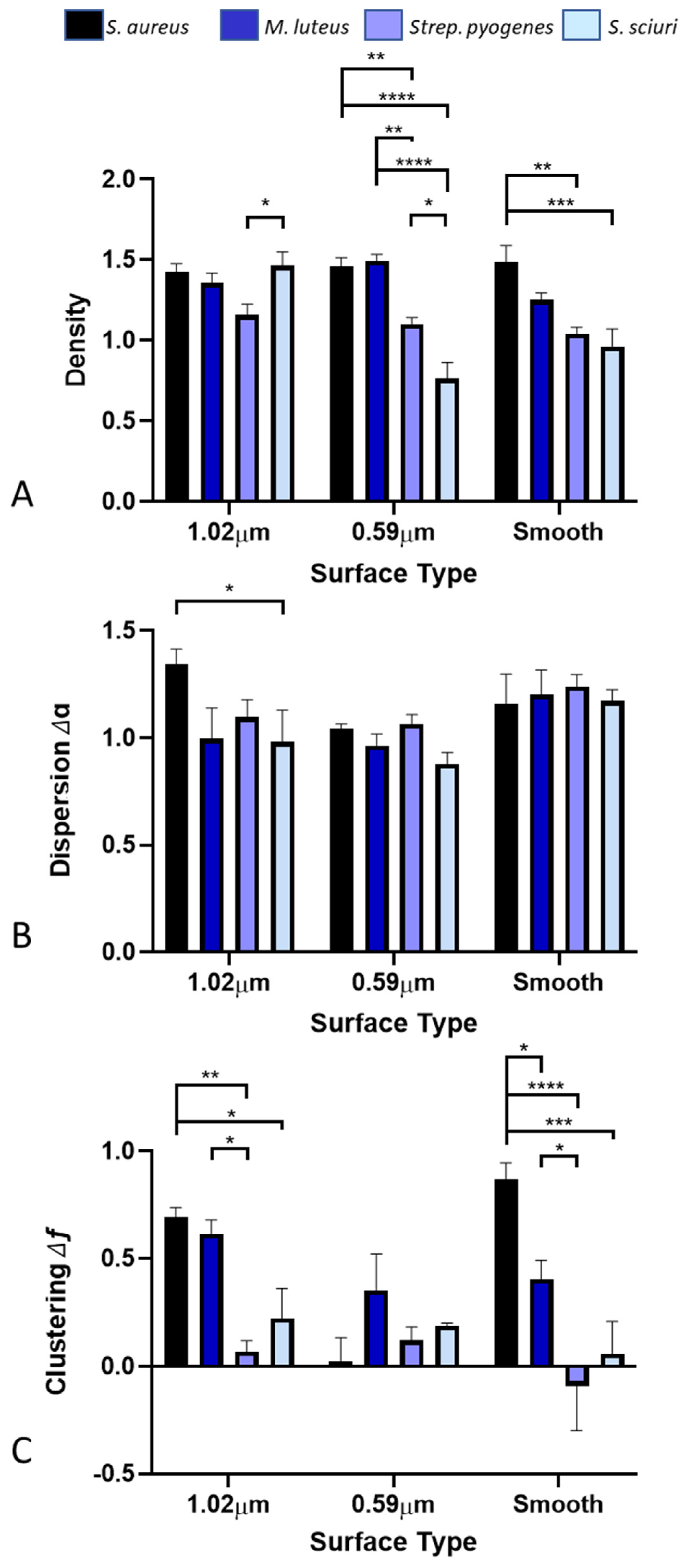
3.2. Distribution of Bacteria across the Surfaces
3.3. Density of Bacteria across the Surfaces
3.4. Dispersion of Bacteria across the Surfaces
3.5. Clustering of Bacteria across the Surfaces
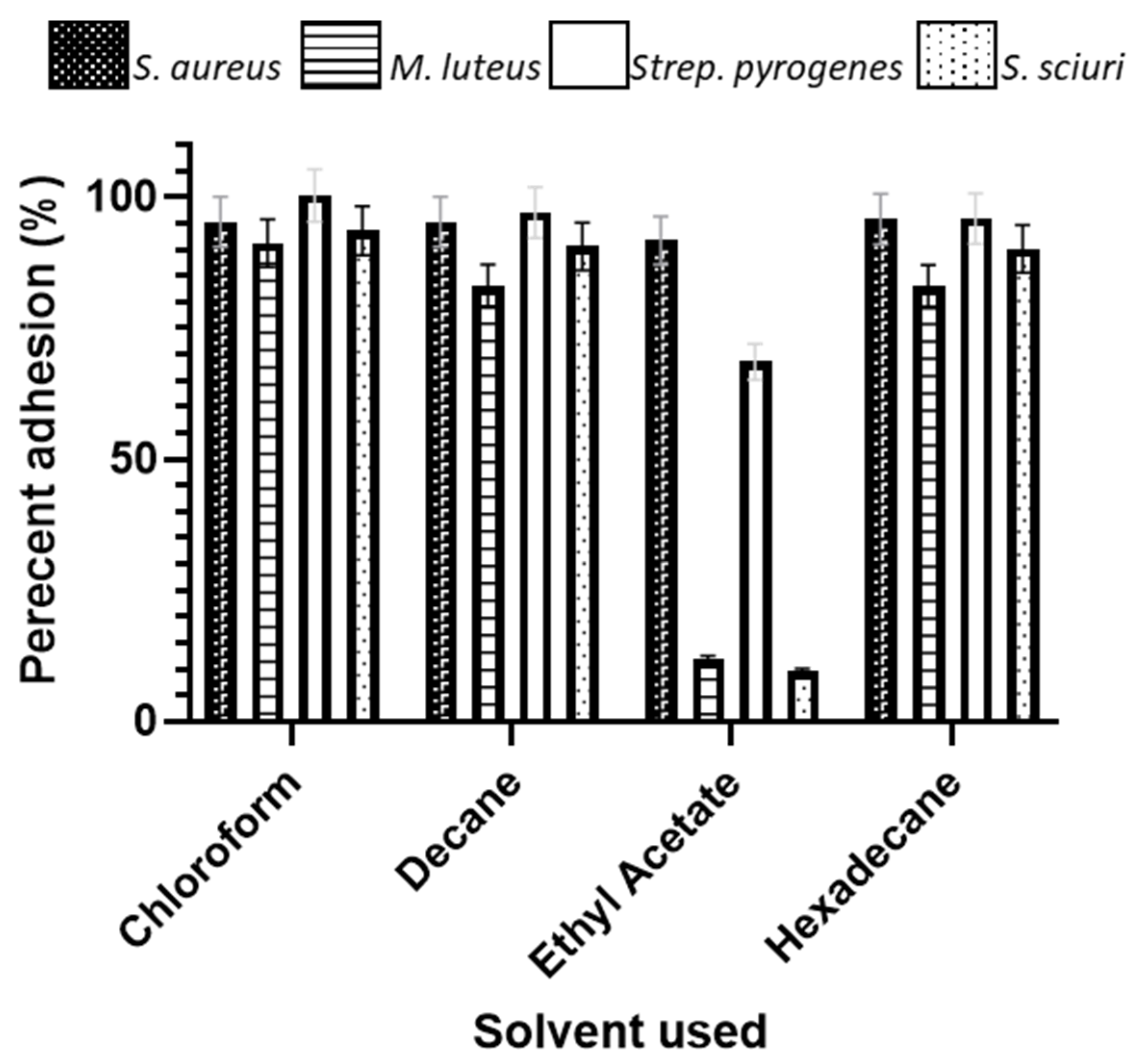
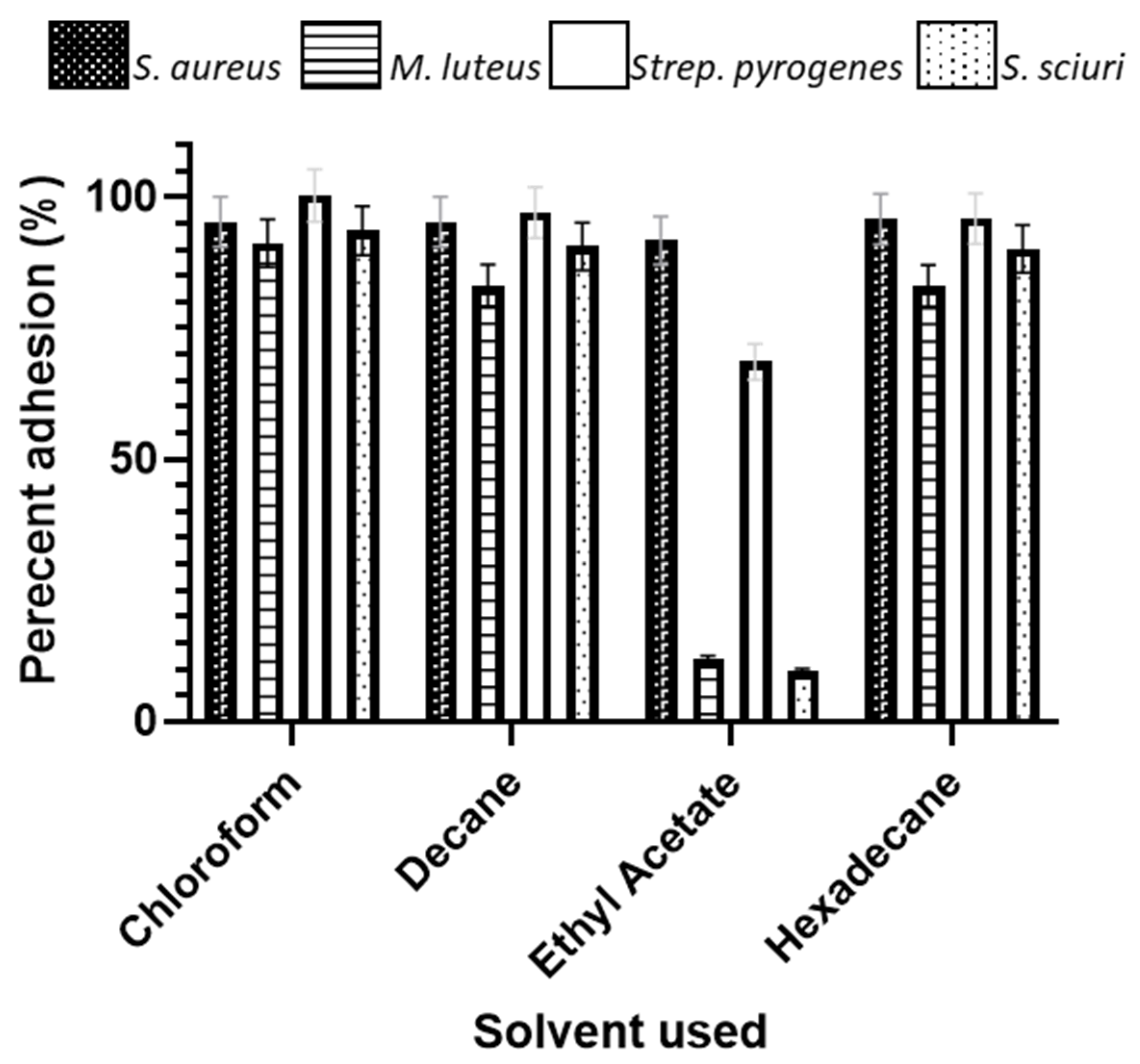
3.6. Microbial Adhesion to Hydrocarbons
4. Discussion
4.1. Microbial Adhesion to Hydrocarbons (MATH)
4.2. Distribution of Bacteria across the Surfaces
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Helbig, R.; Günther, D.; Friedrichs, J.; Rößler, F.; Lasagni, A.; Werner, C. The impact of structure dimensions on initial bacterial adhesion. Biomater. Sci. 2016, 4, 1074–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parreira, P.; Magalhães, A.; Gonçalves, I.C.; Gomes, J.; Vidal, R.; Reis, C.A.; Leckband, D.E.; Martins, M.C.L. Effect of surface chemistry on bacterial adhesion, viability, and morphology. J. Biomed. Mater. Res. Part A 2011, 99A, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Somashekhar, S.; Hiremath, S.; Ramachandran, B.; Muthuvijayan, V. Surface Modification Techniques. In Role of Surface Modification on Bacterial Adhesion of Bio-Implant Materials; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Crawford, R.J.; Webb, H.K.; Truong, V.K.; Hasan, J.; Ivanova, E.P. Surface topographical factors influencing bacterial attachment. Adv. Colloid. Inter. Sci. 2012, 179, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Katsikogianni, M.; Missirlis, Y. Concise review of mechanisms of bacterial adhesion to biomaterials and of techniques used in estimating bacteria-material interactions. Eur. Cells Mater. 2004, 8, 37–57. [Google Scholar] [CrossRef]
- Teughels, W.; Van Assche, N.; Sliepen, I.; Quirynen, M. Effect of material characteristics and/or surface topography on biofilm development. Clin. Oral Implants Res. 2006, 17, 68–81. [Google Scholar] [CrossRef]
- Bédard, E.; Prévost, M.; Déziel, E. Pseudomonas aeruginosa in premise plumbing of large buildings. Microbiol. Open 2016, 5, 937–956. [Google Scholar] [CrossRef]
- Tortora, G.J.; Funke, B.R.; Case, C.L.; Weber, D.; Bair, W. Microbiology: An Introduction; Benjamin Cummings: San Francisco, CA, USA, 2004. [Google Scholar]
- Whitehead, K.A.; Colligon, J.; Verran, J. Retention of microbial cells in substratum surface features of micrometer and sub-micrometer dimensions. Colloids Surf. B Biointerfaces 2005, 41, 129–138. [Google Scholar] [CrossRef]
- Caro-Lara, L.; Ramos-Moore, E.; Vargas, I.T.; Walczak, M.; Fuentes, C.; Gómez, A.V.; Barrera, N.P.; Castillo, J.; Pizarro, G. Initial adhesion suppression of biofilm-forming and copper-tolerant bacterium Variovorax sp. on laser microtextured copper surfaces. Colloids Surf. B Biointerfaces 2021, 202, 111656. [Google Scholar] [CrossRef]
- Goulter, R.M.; Gentle, I.R.; Dykes, G.A. Issues in determining factors influencing bacterial attachment: A review using the attachment of Escherichia coli to abiotic surfaces as an example. Lett. Appl. Microbiol. 2009, 49, 1–7. [Google Scholar] [CrossRef]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Sticky Situations: Key Components That Control Bacterial Surface Attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Cameotra, S.S. Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol. 2004, 22, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Berne, C.; Ducret, A.; Brun, Y.; Hardy, G. Adhesins Involved in Attachment to Abiotic Surfaces by Gram-Negative Bacteria. Microb. Biofilms 2015, 3, 163–199. [Google Scholar]
- Kimkes, T.E.P.; Heinemann, M. How bacteria recognise and respond to surface contact. FEMS Microbiol. Rev. 2019, 44, 106–122. [Google Scholar] [CrossRef]
- Barnes, L.-M.; Lo, M.; Adams, M.; Chamberlain, A. Biocontamination Risk of Staphylococcus aureus in Dairy Manufacturing Presence of Two Types of UHT Milk. Appl. Environ. Microbiol. 1999, 65, 4543–4548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møretrø, T.; Langsrud, S. Residential Bacteria on Surfaces in the Food Industry and Their Implications for Food Safety and Quality. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Verran, J.; Packer, A.; Kelly, P.; Whitehead, K. Use of the Atomic Force Microscope to Determine the Strength of Bacterial Attachment to Grooved Surface Features. J. Adh. Sci. Technol. 2010, 24, 2271–2285. [Google Scholar] [CrossRef]
- Wickens, D.; Lynch, S.; West, G.; Kelly, P.; Verran, J.; Whitehead, K.A. Quantifying the pattern of microbial cell dispersion, density and clustering on surfaces of differing chemistries and topographies using multifractal analysis. J. Microbiol. Method 2014, 104, 101–108. [Google Scholar] [CrossRef]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Hardie, J.M.; Whiley, R.A. Phenotypic differentiation of Streptococcus intermedius, Streptococcus constellatus, and Streptococcus anginosus strains within the “Streptococcus milleri group”. J. Appl. Microbiol. 1997, 83, 1S–11S. [Google Scholar] [CrossRef] [PubMed]
- Kloos, W.E.; Schleifer, K.H.; Smith, R.F.; Genus, I.V. Staphylococcus. Int. J. Syst. Evol. Microbiol. 1976, 26, 22–37. [Google Scholar]
- Zhou, W.; Li, Y. Atlas of Oral Microbiology: From Healthy Microflora to Disease; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Zheng, C.; Qu, B.; Wang, J.; Zhou, J.; Lu, H. Isolation and characterization of a novel nitrobenzene-degrading bacterium with high salinity tolerance: Micrococcus luteus. J. Hazard. Mater. 2009, 165, 1152–1158. [Google Scholar] [CrossRef]
- Monteiro, J.M.; Fernandes, P.B.; Vaz, F.; Pereira, A.R.; Tavares, A.C.; Ferreira, M.T.; Pereira, P.M.; Veiga, H.; Kuru, E.; VanNieuwenhze, M.S. Cell shape dynamics during the staphylococcal cell cycle. Nat. Commun. 2015, 6, 8055. [Google Scholar] [CrossRef]
- Chavez, P.; Yarleque, C.; Loayza, H.; Mares, V.; Hancco, P.; Priou, S.; del Pilar Marquez, M.; Posadas, A.; Zorogastua, P.; Flexas, J.; et al. Detection of Bacterial Wilt Infection Caused by Ralstonia solanacearum in Potato (Solanum tuberosum L.) through Multifractal Analysis Applied to Remotely Sensed Data. Precis. Agric. 2012, 13, 236–255. [Google Scholar] [CrossRef]
- van der Wal, A.; Tecon, R.; Kreft, J.-U.; Mooij, W.M.; Leveau, J.H.J. Explaining bacterial dispersion on leaf surfaces with an individual-based model (PHYLLOSIM). PLoS ONE 2013, 8, e75633. [Google Scholar] [CrossRef]
- Moreira, J.; Gomes, L.; Whitehead, K.; Lynch, S.; Tetlow, L.; Mergulhão, F. Effect of surface conditioning with cellular extracts on Escherichia coli adhesion and initial biofilm formation. Food Bioprod. Process. 2017, 104, 1–12. [Google Scholar] [CrossRef]
- Vourch, T.; Léopoldès, J.; Peerhossaini, H. Clustering of bacteria with heterogeneous motility. Phys. Rev. 2020, 101, 022612. [Google Scholar] [CrossRef] [Green Version]
- Verran, J.; Packer, A.; Kelly, P.; Whitehead, K.A. The retention of bacteria on hygienic surfaces presentingscratches of microbial dimensions. Lett. Appl. Microbiol. 2010, 50, 258–263. [Google Scholar] [CrossRef]
- Tetlow, L.A.; Lynch, S.; Whitehead, K.A. The effect of surface properties on bacterial retention: A study utilising stainless steel and TiN/25.65 at.% Ag substrata. Food Bioprod. Process. 2017, 102, 332–339. [Google Scholar] [CrossRef]
- Kunrath, M.F.; Monteiro, M.S.G.; Gupta, S.; Hubler, R.; de Oliveira, S.D. Influence of titanium and zirconia modified surfaces for rapid healing on adhesion and biofilm formation of Staphylococcus epidermidis. Arch. Oral Biol. 2020, 117, 104824. [Google Scholar] [CrossRef] [PubMed]
- Bollen, C.M.L.; Lambrechts, P.; Quirynen, M. Comparison of surface roughness of oral hard materials to the threshold surface roughness for bacterial plaque retention: A review of the literature. Dent. Mater. 1997, 13, 258–269. [Google Scholar] [CrossRef]
- Roehling, S.; Astasov-Frauenhoffer, M.; Hauser-Gerspach, I.; Braissant, O.; Woelfler, H.; Waltimo, T.; Kniha, H.; Gahlert, M. In vitro biofilm formation on titanium and zirconia implant surfaces. J. Periodontol. 2017, 88, 298–307. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Karygianni, L.; Wartenhorst, M.S.; Bächle, M.; Hellwig, E.; Follo, M.; Vach, K.; Han, J.-S. Bacterial adhesion and biofilm formation on yttria-stabilized, tetragonal zirconia and titanium oral implant materials with low surface roughness-An in situ study. J. Med. Microbiol. 2016, 65, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.E.; O’Sullivan, D.J.; Jenkinson, H.F.; Jagger, D.C. The effects of polishing methods on surface morphology, roughness and bacterial colonisation of titanium abutments. J. Mater. Sci. Mater. Med. 2007, 18, 1439–1447. [Google Scholar] [CrossRef]
- Duarte, P.M.; Reis, A.F.; Moreira de Freitas, P.; Ota-Tsuzuki, C. Bacterial adhesion on smooth and rough titanium surfaces after treatment with different instruments. J. Periodontol. 2009, 80, 1824–1832. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, A.; Slate, A.J.; Tobin, M.; Lynch, S.; Wilson Nieuwenhuis, J.; Verran, J.; Kelly, P.; Whitehead, K.A. Multifractal Analysis to Determine the Effect of Surface Topography on the Distribution, Density, Dispersion and Clustering of Differently Organised Coccal-Shaped Bacteria. Antibiotics 2022, 11, 551. https://doi.org/10.3390/antibiotics11050551
Evans A, Slate AJ, Tobin M, Lynch S, Wilson Nieuwenhuis J, Verran J, Kelly P, Whitehead KA. Multifractal Analysis to Determine the Effect of Surface Topography on the Distribution, Density, Dispersion and Clustering of Differently Organised Coccal-Shaped Bacteria. Antibiotics. 2022; 11(5):551. https://doi.org/10.3390/antibiotics11050551
Chicago/Turabian StyleEvans, Adele, Anthony J. Slate, Millie Tobin, Stephen Lynch, Joels Wilson Nieuwenhuis, Joanna Verran, Peter Kelly, and Kathryn A. Whitehead. 2022. "Multifractal Analysis to Determine the Effect of Surface Topography on the Distribution, Density, Dispersion and Clustering of Differently Organised Coccal-Shaped Bacteria" Antibiotics 11, no. 5: 551. https://doi.org/10.3390/antibiotics11050551
APA StyleEvans, A., Slate, A. J., Tobin, M., Lynch, S., Wilson Nieuwenhuis, J., Verran, J., Kelly, P., & Whitehead, K. A. (2022). Multifractal Analysis to Determine the Effect of Surface Topography on the Distribution, Density, Dispersion and Clustering of Differently Organised Coccal-Shaped Bacteria. Antibiotics, 11(5), 551. https://doi.org/10.3390/antibiotics11050551