
SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces

Abstract
:1. Introduction
2. Results
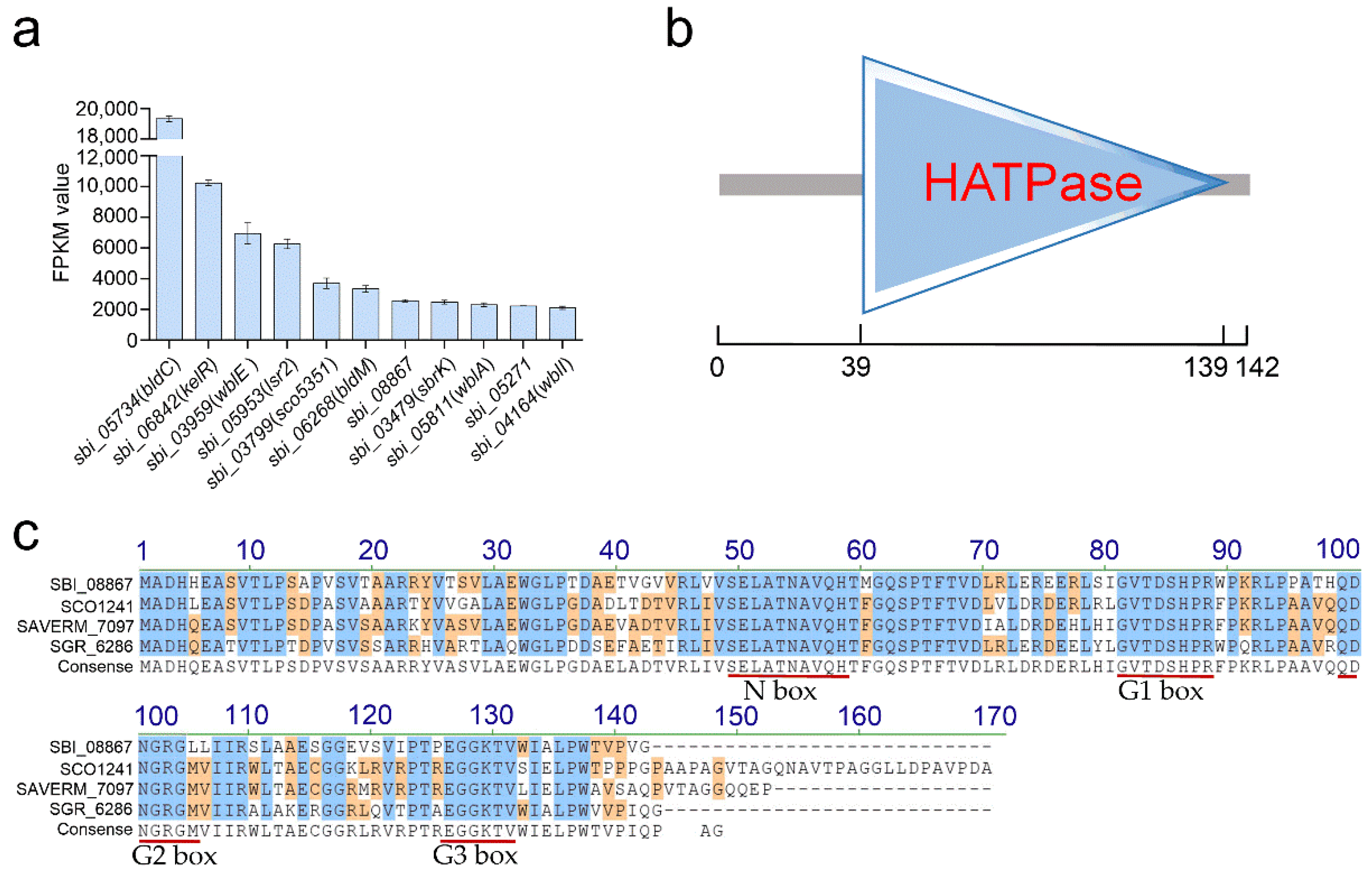
2.1. Identification of the Novel HATPase-ol Family Regulator SspH in S. bingchenggensis
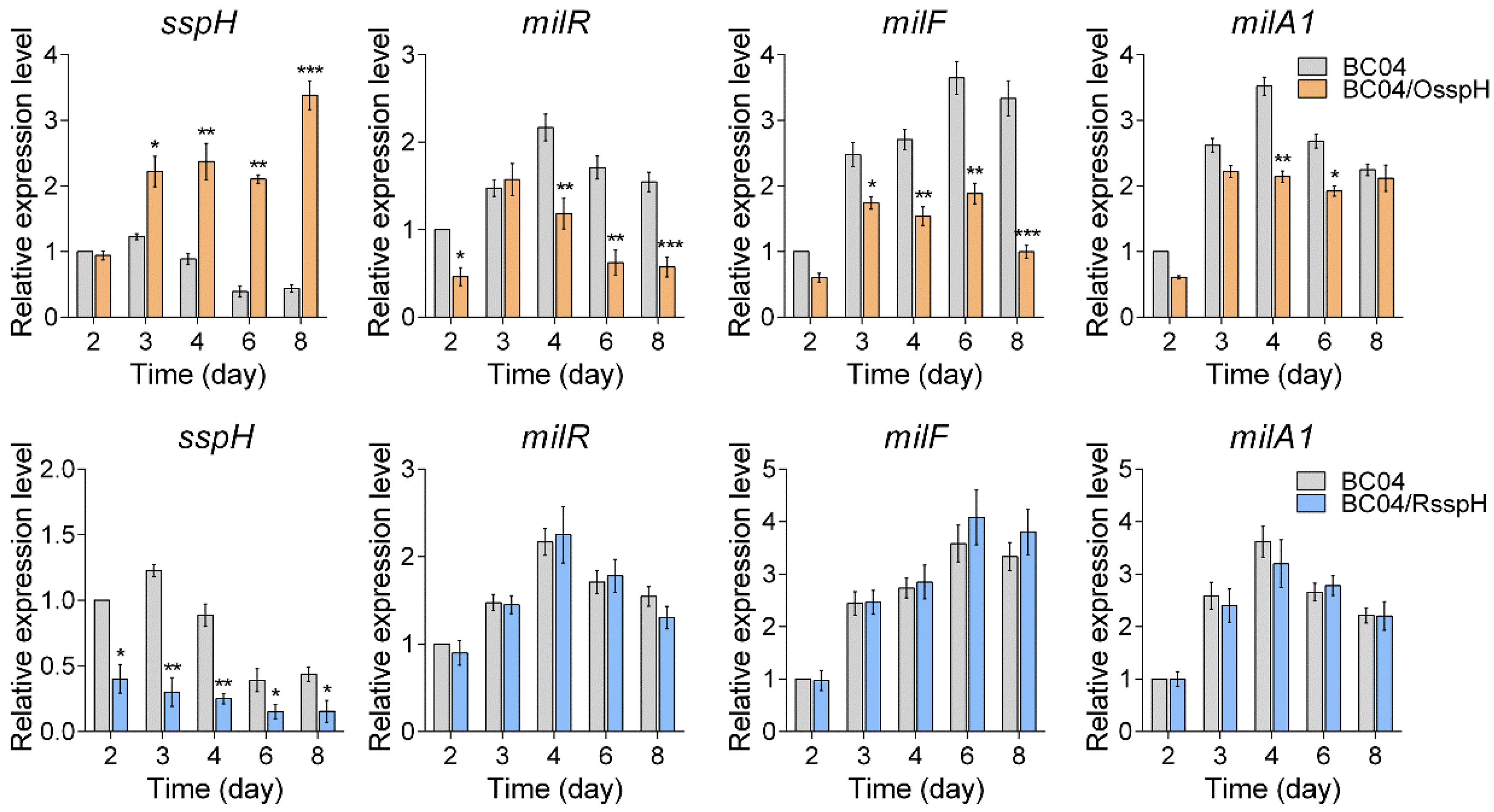
2.2. SspH Negatively Influences Milbemycin Production
2.3. SspH Represses the Transcription of milR, milF and milA1
2.4. The sspH Overexpression in BC04 Differentially Affects Expression of Multiple PKS/NRPS/PKS-NRPS Genes
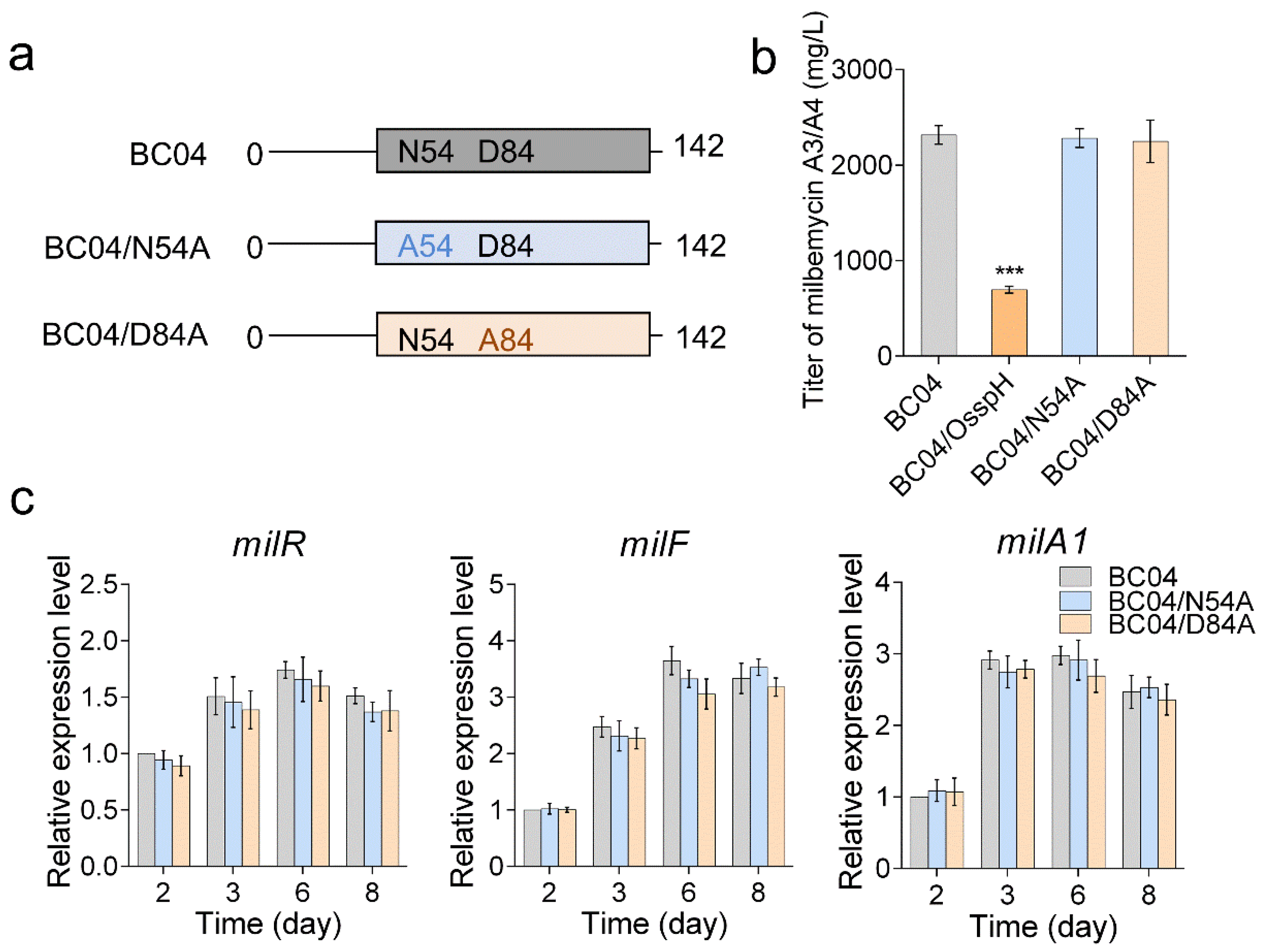
2.5. The Conserved HATPase Domain Is Essential for SspH Function
2.6. SspH Is Commonly Involved in the Control of Antibiotic Production in Streptomyces
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids and Culture Conditions
4.2. Gene Overexpression and Repression
4.3. Site-Directed Mutagenesis of sspHpH
4.4. Detection Methods of Milbemycins, Actinorhodin and Avermectins
4.5. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR) Assay
4.6. Sequence Analysis
4.7. Statistical Analysis
4.8. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nett, M.; Ikeda, H.; Moore, B.S. Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 2009, 26, 1362–1384. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Ning, S.; Luo, Y. Coordinated regulation for nature products discovery and overproduction in Streptomyces. Synth. Syst. Biotechnol. 2020, 5, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Li, X.; Li, Z.; Zhan, X.; Mao, X.; Li, Y. The Application of Regulatory Cascades in Streptomyces: Yield Enhancement and Metabolite Mining. Front. Microbiol. 2020, 11, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Agudelo, V.A.; Gómez-Ríos, D.; Ramirez-Malule, H. Clavulanic Acid Production by Streptomyces clavuligerus: Insights from Systems Biology, Strain Engineering, and Downstream Processing. Antibiotics 2021, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Bu, Q.; Li, Y.; Xie, H.; Li, J.; Lv, Z.; Su, Y.; Li, Y. Rational engineering strategies for achieving high-yield, high-quality and high-stability of natural product production in actinomycetes. Metab. Eng. 2021, 67, 198–215. [Google Scholar] [CrossRef]
- Liu, N.; Guan, H.; Niu, G.; Jiang, L.; Li, Y.; Zhang, J.; Li, J.; Tan, H. Molecular mechanism of mureidomycin biosynthesis activated by introduction of an exogenous regulatory gene ssaA into Streptomyces roseosporus. Sci. China Life Sci. 2021, 64, 1949–1963. [Google Scholar] [CrossRef]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Zhu, H.; Dang, F.; Zhang, W.; Qin, Z.; Yang, S.; Tan, H.; Lu, Y.; Jiang, W. Differential regulation of antibiotic biosynthesis by DraR-K, a novel two-component system in Streptomyces coelicolor. Mol. Microbiol. 2012, 85, 535–556. [Google Scholar] [CrossRef]
- Wang, L.; Tian, X.; Wang, J.; Yang, H.; Fan, K.; Xu, G.; Yang, K.; Tan, H. Autoregulation of antibiotic biosynthesis by binding of the end product to an atypical response regulator. Proc. Natl. Acad. Sci. USA 2009, 106, 8617–8622. [Google Scholar] [CrossRef] [Green Version]
- Dutta, R.; Inouye, M. GHKL, an emergent ATPase/kinase superfamily. Trends Biochem. Sci. 2000, 25, 24–28. [Google Scholar] [CrossRef]
- Robinson, V.L.; Buckler, D.R.; Stock, A.M. A tale of two components: A novel kinase and a regulatory switch. Nat. Struct. Biol. 2000, 7, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, H.; Liu, H.; Wang, H.; Wang, X.; Xiang, W. Characterization of a pathway-specific activator of milbemycin biosynthesis and improved milbemycin production by its overexpression in Streptomyces bingchenggensis. Microb. Cell Fact. 2016, 15, 152. [Google Scholar] [CrossRef]
- Wei, K.; Wu, Y.; Li, L.; Jiang, W.; Hu, J.; Lu, Y.; Chen, S. MilR2, a novel TetR family regulator involved in 5-oxomilbemycin A3/A4 biosynthesis in Streptomyces hygroscopicus. Appl. Microbiol. Biotechnol. 2018, 102, 8841–8853. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ye, L.; Li, C.; Wang, H.; Guo, X.; Wang, X.; Zhang, Y.; Xiang, W. SbbR/SbbA, an Important ArpA/AfsA-Like System, Regulates Milbemycin Production in Streptomyces bingchenggensis. Front. Microbiol. 2018, 9, 1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, Y.; Cheng, X.; Zhang, Y.; Li, S.; Wang, X.; Xiang, W. Titer improvement of milbemycins via coordinating metabolic competition and transcriptional co-activation controlled by Streptomyces antibiotic regulatory protein family regulator in Streptomyces bingchenggensis. Biotechnol. Bioeng. 2022, 119, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Y.; Li, S.; He, H.; Ai, G.; Wang, X.; Xiang, W. Transcriptome-guided identification of a four-component system, SbrH1-R, that modulates milbemycin biosynthesis by influencing gene cluster expression, precursor supply, and antibiotic efflux. Synth. Syst. Biotechnol. 2022, 7, 705–717. [Google Scholar] [CrossRef]
- Schumacher, M.A.; den Hengst, C.D.; Bush, M.J.; Le, T.B.K.; Tran, N.T.; Chandra, G.; Zeng, W.; Travis, B.; Brennan, R.G.; Buttner, M.J. The MerR-like protein BldC binds DNA direct repeats as cooperative multimers to regulate Streptomyces development. Nat. Commun. 2018, 9, 1139. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cheng, Y.; Lyu, M.; Wen, Y.; Song, Y.; Chen, Z.; Li, J. Redox-sensing regulator Rex regulates aerobic metabolism, morphological differentiation, and avermectin production in Streptomyces avermitilis. Sci. Rep. 2017, 7, 44567. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Andres, S.N.; Elliot, M.A. Interplay between Nucleoid-Associated Proteins and Transcription Factors in Controlling Specialized Metabolism in Streptomyces. mBio 2021, 12, e0107721. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Y.; Zhang, P.; Sheng, D.; Cao, G.; Pang, X. SCO5351 is a pleiotropic factor that impacts secondary metabolism and morphological development in Streptomyces coelicolor. FEMS Microbiol. Lett. 2018, 365, fny15. [Google Scholar] [CrossRef] [Green Version]
- Al-Bassam, M.M.; Bibb, M.J.; Bush, M.J.; Chandra, G.; Buttner, M.J. Response regulator heterodimer formation controls a key stage in Streptomyces development. PLoS Genet. 2014, 10, e1004554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, M.J. The actinobacterial WhiB-like (Wbl) family of transcription factors. Mol. Microbiol. 2018, 110, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhang, Q.; Virolle, M.J.; Xu, D. In conditions of over-expression, WblI, a WhiB-like transcriptional regulator, has a positive impact on the weak antibiotic production of Streptomyces lividans TK24. PLoS ONE 2017, 12, e0174781. [Google Scholar]
- Wang, H.; Zhang, J.; Zhang, Y.; Zhang, B.; Liu, C.; He, H.; Wang, X.; Xiang, W. Combined application of plasma mutagenesis and gene engineering leads to 5-oxomilbemycins A3/A4 as main components from Streptomyces bingchenggensis. Appl. Microbiol. Biotechnol. 2014, 98, 9703–9712. [Google Scholar] [CrossRef]
- Kim, M.S.; Cho, W.J.; Song, M.C.; Park, S.W.; Kim, K.; Kim, E.; Lee, N.; Nam, S.J.; Oh, K.H.; Yoon, Y.J. Engineered biosynthesis of milbemycins in the avermectin high-producing strain Streptomyces avermitilis. Microb. Cell Fact. 2017, 16, 9. [Google Scholar] [CrossRef] [Green Version]
- Nowak, E.; Panjikar, S.; Morth, J.P.; Jordanova, R.; Svergun, D.I.; Tucker, P.A. Structural and functional aspects of the sensor histidine kinase PrrB from Mycobacterium tuberculosis. Structure 2006, 14, 275–285. [Google Scholar] [CrossRef]
- Niu, G. Genomics-Driven Natural Product Discovery in Actinomycetes. Trends Biotechnol. 2018, 36, 238–241. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, J.; Zhuo, J.; Li, Y.; Tian, Y.; Tan, H. Activation and mechanism of a cryptic oviedomycin gene cluster via the disruption of a global regulatory gene, adpA, in Streptomyces ansochromogenes. J. Biol. Chem. 2017, 292, 19708–19720. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Lu, Y.; Chen, J.; Zhang, W.; Shu, D.; Qin, Z.; Yang, S.; Jiang, W. Characterization of a negative regulator AveI for avermectin biosynthesis in Streptomyces avermitilis NRRL8165. Appl. Microbiol. Biotechnol. 2008, 80, 277–286. [Google Scholar] [CrossRef]
- Zou, Z.; Du, D.; Zhang, Y.; Zhang, J.; Niu, G.; Tan, H. A γ-butyrolactone-sensing activator/repressor, JadR3, controls a regulatory mini-network for jadomycin biosynthesis. Mol. Microbiol. 2014, 94, 490–505. [Google Scholar] [CrossRef]
- Van Schaik, W.; Tempelaars, M.H.; Zwietering, M.H.; de Vos, W.M.; Abee, T. Analysis of the role of RsbV, RsbW, and RsbY in regulating σB activity in Bacillus cereus. J. Bacteriol. 2005, 187, 5846–5851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, D.; Jin, K.S.; Tandukar, S.; Kim, J.H.; Kwon, E.; Kim, D.Y. Structural insights into the regulation of SigB activity by RsbV and RsbW. IUCrJ 2020, 7, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Shinose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Omura, S. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Sambrock, J.; Russel, D.J.I. Molecular Cloning: A Laboratory Manual; Cold Spring Harbour Laboratory Press: New York, NY, USA, 2001; Volume 49, pp. 895–909. [Google Scholar]
- Keiser, T.; Bipp, M.; Buttner, M.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Gao, H.; Liu, M.; Liu, J.; Dai, H.; Zhou, X.; Liu, X.; Zhuo, Y.; Zhang, W.; Zhang, L. Medium optimization for the production of avermectin B1a by Streptomyces avermitilis 14-12A using response surface methodology. Bioresour. Technol. 2009, 100, 4012–4016. [Google Scholar] [CrossRef] [PubMed]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Rao, R.N.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef]
- Li, L.; Wei, K.; Zheng, G.; Liu, X.; Chen, S.; Jiang, W.; Lu, Y. CRISPR-Cpf1-Assisted Multiplex Genome Editing and Transcriptional Repression in Streptomyces. Appl. Environ. Microbiol. 2018, 84, e00827-18. [Google Scholar] [CrossRef] [Green Version]
- Strauch, E.; Takano, E.; Baylis, H.A.; Bibb, M.J. The stringent response in Streptomyces coelicolor A3(2). Mol. Microbiol. 1991, 5, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 4, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Type of Product | Reported Homolog | Predicted Function | Reference(s) |
|---|---|---|---|---|
| sbi_05734 | MerR | BldC (SCO4091) | Widespread; a repressor to maintain vegetative growth and to delay entry into development. | [17] |
| sbi_06842 | SARP | - | A cluster-situated activator of type II PKS cluster, also essential for milbemycin production. | [15] |
| sbi_03959 | Wbl | WblE (SAVERM_3016) | Widespread; an important regulator of morphological differentiation. | [18] |
| sbi_05953 | Lsr2 | Lsr2 (SVEN_3225) | Widespread; a nucleoid-associated protein that influences chromosome segregation, DNA replication, transcription and secondary metabolism. | [19] |
| sbi_03799 | TCS (RR) | SCO5351 | Widespread; a pleiotropic regulator involved in secondary metabolism and development. | [20] |
| sbi_06268 | TCS (RR) | BldM (SCO4768) | Widespread; an essential regulator for aerial hyphae formation. | [21] |
| sbi_08867 | HATPase-ol | - | Widespread; function unknown. | This work |
| sbi_03479 | TCS (HK) | SbrK | A repressor of milbemycin biosynthesis. | [16] |
| sbi_05811 | Wbl | WblA (SCO3579) | Widespread; a global regulator involved in differentiation and secondary metabolism. | [22] |
| sbi_05271 | MarR | - | Function unknown. | - |
| sbi_04164 | Wbl | WblI (SCO5046) | Widespread; a positive impact on antibiotic production. | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Zhang, Y.; Li, S.; Ye, L.; Wang, X.; Xiang, W. SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces. Antibiotics 2022, 11, 538. https://doi.org/10.3390/antibiotics11050538
Yang X, Zhang Y, Li S, Ye L, Wang X, Xiang W. SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces. Antibiotics. 2022; 11(5):538. https://doi.org/10.3390/antibiotics11050538
Chicago/Turabian StyleYang, Xue, Yanyan Zhang, Shanshan Li, Lan Ye, Xiangjing Wang, and Wensheng Xiang. 2022. "SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces" Antibiotics 11, no. 5: 538. https://doi.org/10.3390/antibiotics11050538
APA StyleYang, X., Zhang, Y., Li, S., Ye, L., Wang, X., & Xiang, W. (2022). SspH, a Novel HATPase Family Regulator, Controls Antibiotic Biosynthesis in Streptomyces. Antibiotics, 11(5), 538. https://doi.org/10.3390/antibiotics11050538