
Genomic Characterization of Clinical Acinetobacter baumannii Isolates Obtained from COVID-19 Patients in Russia

 , , and
, , and
Abstract
:1. Introduction
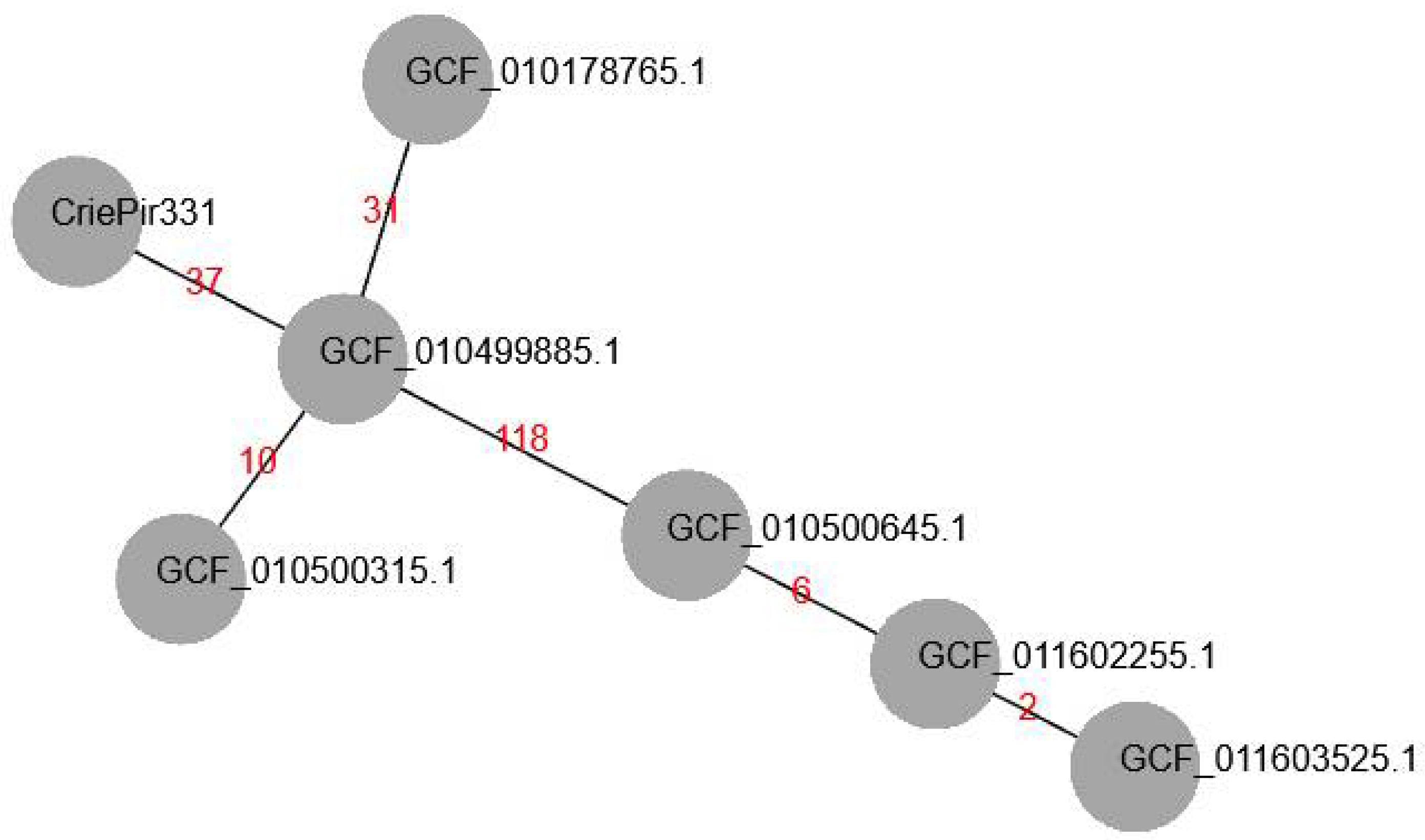
2. Results
3. Discussion
4. Materials and Methods
4.1. Determination of Antibiotic Susceptibility
4.2. DNA Isolation, Sequencing and Genome Assembly
4.3. Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Soucy, J.R.; Westwood, D.; Daneman, N.; MacFadden, D.R. Antibiotic prescribing in patients with COVID-19: Rapid review and meta-analysis. Clin. Microbiol. Infect. 2021, 27, 520–531. [Google Scholar] [CrossRef] [PubMed]
- The Pew Charitable Trusts. Could Efforts to Fight the Coronavirus Lead to Overuse of Antibiotics? 2021. Available online: https://www.pewtrusts.org/en/research-and-analysis/issue-briefs/2021/03/could-efforts-to-fight-the-coronavirus-lead-to-overuse-of-antibiotics (accessed on 25 January 2022).
- Pasero, D.; Cossu, A.P.; Terragni, P. Multi-Drug Resistance Bacterial Infections in Critically Ill Patients Admitted with COVID-19. Microorganisms 2021, 9, 1773. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. Economic burden of antibiotic resistance in ESKAPE organisms: A systematic review. Antimicrob. Resist Infect. Control 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenkov, A.Y.; Trushin, I.V.; Vinogradova, A.G.; Avramenko, A.A.; Sukhorukova, M.V.; Malhotra-Kumar, S.; Dekhnich, A.V.; Edelstein, M.V.; Kozlov, R.S. AMRmap: An Interactive Web Platform for Analysis of Antimicrobial Resistance Surveillance Data in Russia. Front. Microbiol. 2021, 12, 620002. [Google Scholar] [CrossRef] [PubMed]
- Al-Shamiri, M.M.; Zhang, S.; Mi, P.; Liu, Y.; Xun, M.; Yang, E.; Ai, L.; Han, L.; Chen, Y. Phenotypic and genotypic characteristics of Acinetobacter baumannii enrolled in the relationship among antibiotic resistance, biofilm formation and motility. Microb. Pathog. 2021, 155, 104922. [Google Scholar] [CrossRef]
- Shelenkov, A.; Petrova, L.; Zamyatin, M.; Mikhaylova, Y.; Akimkin, V. Diversity of International High-Risk Clones of Acinetobacter baumannii Revealed in a Russian Multidisciplinary Medical Center during 2017–2019. Antibiotics 2021, 10, 1009. [Google Scholar] [CrossRef]
- Hamidian, M.; Nigro, S.J. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microb. Genom. 2019, 5, e000306. [Google Scholar] [CrossRef]
- Saini, V.; Jain, C.; Singh, N.P.; Alsulimani, A.; Gupta, C.; Dar, S.A.; Haque, S.; Das, S. Paradigm Shift in Antimicrobial Resistance Pattern of Bacterial Isolates during the COVID-19 Pandemic. Antibiotics 2021, 10, 954. [Google Scholar] [CrossRef]
- Lai, C.C.; Chen, S.Y.; Ko, W.C.; Hsueh, P.R. Increased antimicrobial resistance during the COVID-19 pandemic. Int. J. Antimicrob. Agents 2021, 57, 106324. [Google Scholar] [CrossRef]
- Taylor, L. Covid-19: Antimicrobial misuse in Americas sees drug resistant infections surge, says WHO. BMJ 2021, 375, n2845. [Google Scholar] [CrossRef] [PubMed]
- Chamieh, A.; Zgheib, R.; El-Sawalhi, S.; Yammine, L.; El-Hajj, G.; Zmerli, O.; Afif, C.; Rolain, J.M.; Azar, E. Trends of Multidrug-Resistant Pathogens, Difficult to Treat Bloodstream Infections, and Antimicrobial Consumption at a Tertiary Care Center in Lebanon from 2015–2020: COVID-19 Aftermath. Antibiotics 2021, 10, 1016. [Google Scholar] [CrossRef] [PubMed]
- Guisado-Gil, A.B.; Infante-Dominguez, C.; Penalva, G.; Praena, J.; Roca, C.; Navarro-Amuedo, M.D.; Aguilar-Guisado, M.; Espinosa-Aguilera, N.; Poyato-Borrego, M.; Romero-Rodriguez, N.; et al. Impact of the COVID-19 Pandemic on Antimicrobial Consumption and Hospital-Acquired Candidemia and Multidrug-Resistant Bloodstream Infections. Antibiotics 2020, 9, 816. [Google Scholar] [CrossRef] [PubMed]
- Gandra, S.; Tseng, K.K.; Arora, A.; Bhowmik, B.; Robinson, M.L.; Panigrahi, B.; Laxminarayan, R.; Klein, E.Y. The Mortality Burden of Multidrug-resistant Pathogens in India: A Retrospective, Observational Study. Clin. Infect. Dis. 2019, 69, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colomb-Cotinat, M.; Lacoste, J.; Brun-Buisson, C.; Jarlier, V.; Coignard, B.; Vaux, S. Estimating the morbidity and mortality associated with infections due to multidrug-resistant bacteria (MDRB), France, 2012. Antimicrob. Resist Infect. Control 2016, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Alqahtani, A.; Alamer, E.; Mir, M.; Alasmari, A.; Alshahrani, M.M.; Asiri, M.; Ahmad, I.; Alhazmi, A.; Algaissi, A. Bacterial Coinfections Increase Mortality of Severely Ill COVID-19 Patients in Saudi Arabia. Int. J. Environ. Res. Public Health 2022, 19, 2424. [Google Scholar] [CrossRef]
- Silva, D.L.; Lima, C.M.; Magalhaes, V.C.R.; Baltazar, L.M.; Peres, N.T.A.; Caligiorne, R.B.; Moura, A.S.; Fereguetti, T.; Martins, J.C.; Rabelo, L.F.; et al. Fungal and bacterial coinfections increase mortality of severely ill COVID-19 patients. J. Hosp. Infect. 2021, 113, 145–154. [Google Scholar] [CrossRef]
- Rakiro, J.; Shah, J.; Waweru-Siika, W.; Wanyoike, I.; Riunga, F. Microbial coinfections and superinfections in critical COVID-19: A Kenyan retrospective cohort analysis. IJID Reg. 2021, 1, 41–46. [Google Scholar] [CrossRef]
- Paula, H.S.C.; Santiago, S.B.; Araujo, L.A.; Pedroso, C.F.; Marinho, T.A.; Goncalves, I.A.J.; Santos, T.A.P.; Pinheiro, R.S.; Oliveira, G.A.; Batista, K.A. An overview on the current available treatment for COVID-19 and the impact of antibiotic administration during the pandemic. Braz. J. Med. Biol. Res. 2021, 55, e11631. [Google Scholar] [CrossRef]
- Kim, D.H.; Choi, J.Y.; Kim, H.W.; Kim, S.H.; Chung, D.R.; Peck, K.R.; Thamlikitkul, V.; So, T.M.; Yasin, R.M.; Hsueh, P.R.; et al. Spread of carbapenem-resistant Acinetobacter baumannii global clone 2 in Asia and AbaR-type resistance islands. Antimicrob. Agents Chemother. 2013, 57, 5239–5246. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelenkov, A.; Mikhaylova, Y.; Yanushevich, Y.; Samoilov, A.; Petrova, L.; Fomina, V.; Gusarov, V.; Zamyatin, M.; Shagin, D.; Akimkin, V. Molecular Typing, Characterization of Antimicrobial Resistance, Virulence Profiling and Analysis of Whole-Genome Sequence of Clinical Klebsiella pneumoniae Isolates. Antibiotics 2020, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The population structure of Acinetobacter baumannii: Expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shashkov, A.S.; Cahill, S.M.; Arbatsky, N.P.; Westacott, A.C.; Kasimova, A.A.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; et al. Acinetobacter baumannii K116 capsular polysaccharide structure is a hybrid of the K14 and revised K37 structures. Carbohydr. Res. 2019, 484, 107774. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef] [PubMed]
- Arbatsky, N.P.; Shneider, M.M.; Dmitrenok, A.S.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; Edelstein, M.V.; Knirel, Y.A. Structure and gene cluster of the K125 capsular polysaccharide from Acinetobacter baumannii MAR13-1452. Int. J. Biol. Macromol. 2018, 117, 1195–1199. [Google Scholar] [CrossRef]
- Feijao, P.; Yao, H.T.; Fornika, D.; Gardy, J.; Hsiao, W.; Chauve, C.; Chindelevitch, L. MentaLiST-A fast MLST caller for large MLST schemes. Microb. Genom. 2018, 4, e000146. [Google Scholar] [CrossRef]
- Higgins, P.G.; Prior, K.; Harmsen, D.; Seifert, H. Development and evaluation of a core genome multilocus typing scheme for whole-genome sequence-based typing of Acinetobacter baumannii. PLoS ONE 2017, 12, e0179228. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Isolate | Patient Gender | Patient Age | Isolation Source | Ciprofloxacin | Gentamicin | Imipenem | Levofloxacin | Netilmicin | Tmp/Smz 1 |
|---|---|---|---|---|---|---|---|---|---|
| CriePir331 | Female | 75 | Blood | R | R | R | R | R | R |
| CriePir332 | Male | 78 | BAL 2 | R | R | R | R | R | R |
| CriePir333 | Male | 76 | BAL | R | R | R | R | R | R |
| CriePir345 | Male | 66 | BAL | R | R | R | R | R | R |
| Gene | Function | Affected Antimicrobials |
|---|---|---|
| arr-2 | integron-encoded ribosyltransferase | Rifampicin |
| ant(3″)-Ia | aminoglycoside nucleotidyltransferase | Aminoglycosides |
| aph(3″)-Ia | aminoglycoside phosphotransferase | Aminoglycosides |
| aph(3″)-Ib | aminoglycoside phosphotransferase | Aminoglycosides |
| aph(6)-Id | aminoglycoside phosphotransferase | Aminoglycosides |
| armA | 16S rRNA methyltransferase | Aminoglycosides |
| blaADC-30 | intrinsic ADC beta-lactamase and cephalosporinase of A. baumannii | Cephalosporines |
| blaOXA-23 | carbapenemase | Carbapenems |
| blaOXA-66 | intrinsic OXA-51-like β-lactamase of A. baumannii | Cephalosporines |
| blaPER-7 | extended-spectrum β-lactamase | Penicillins, Cephalosporines |
| catB8 | chloramphenicol acetyltransferase | Chloramphenicol |
| cmlA5 | chloramphenicol exporter | Chloramphenicol |
| mph(E) | macrolide phosphotransferase and resistance gene | Macrolides |
| msr(E) | ABC-F subfamily protein | Macrolides |
| qacE | resistance gene conferring resistance to antiseptics | Antiseptics |
| sul1, sul2 | sulfonamide resistant dihydropteroate synthase | Sulfonamides |
| adeABCRS | multidrug efflux complex and its regulators | β-lactams |
| adeFGHL | multidrug efflux complex and its regulators | Fluoroquinolones |
| ompA | porin, permeability defects | All |
| Gene Cluster | Function |
|---|---|
| abaI | virulence, motility, conjugation, biofilm formation and host-pathogen interactions |
| bap | biofilm formation |
| barAB | siderophore efflux system |
| basABCDFGHJ | iron acquisition system and acinetobactin functioning |
| bauABCDEF | iron acquisition system and acinetobactin functioning |
| bfmRS | quorum sensing-regulated two-component system involved in biofilm formation |
| csuABCDE, csuA/B | biofilm formation |
| entE | enterobactin biosynthesis (siderophore) |
| htpB | heat shock protein |
| katA | oxidative stress resistance |
| pgaABCD | biofilm formation |
| pilT | twitching motility and evasion of host immune system |
| plc, plcD | lipolytic activity for iron acquisition |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shelenkov, A.; Mikhaylova, Y.; Petrova, L.; Gaidukova, I.; Zamyatin, M.; Akimkin, V. Genomic Characterization of Clinical Acinetobacter baumannii Isolates Obtained from COVID-19 Patients in Russia. Antibiotics 2022, 11, 346. https://doi.org/10.3390/antibiotics11030346
Shelenkov A, Mikhaylova Y, Petrova L, Gaidukova I, Zamyatin M, Akimkin V. Genomic Characterization of Clinical Acinetobacter baumannii Isolates Obtained from COVID-19 Patients in Russia. Antibiotics. 2022; 11(3):346. https://doi.org/10.3390/antibiotics11030346
Chicago/Turabian StyleShelenkov, Andrey, Yulia Mikhaylova, Lyudmila Petrova, Irina Gaidukova, Mikhail Zamyatin, and Vasiliy Akimkin. 2022. "Genomic Characterization of Clinical Acinetobacter baumannii Isolates Obtained from COVID-19 Patients in Russia" Antibiotics 11, no. 3: 346. https://doi.org/10.3390/antibiotics11030346
APA StyleShelenkov, A., Mikhaylova, Y., Petrova, L., Gaidukova, I., Zamyatin, M., & Akimkin, V. (2022). Genomic Characterization of Clinical Acinetobacter baumannii Isolates Obtained from COVID-19 Patients in Russia. Antibiotics, 11(3), 346. https://doi.org/10.3390/antibiotics11030346