
Identification of Methicillin-Resistant Staphylococcus aureus (MRSA) Genetic Factors Involved in Human Endothelial Cells Damage, an Important Phenotype Correlated with Persistent Endovascular Infection

Abstract
1. Introduction
2. Results
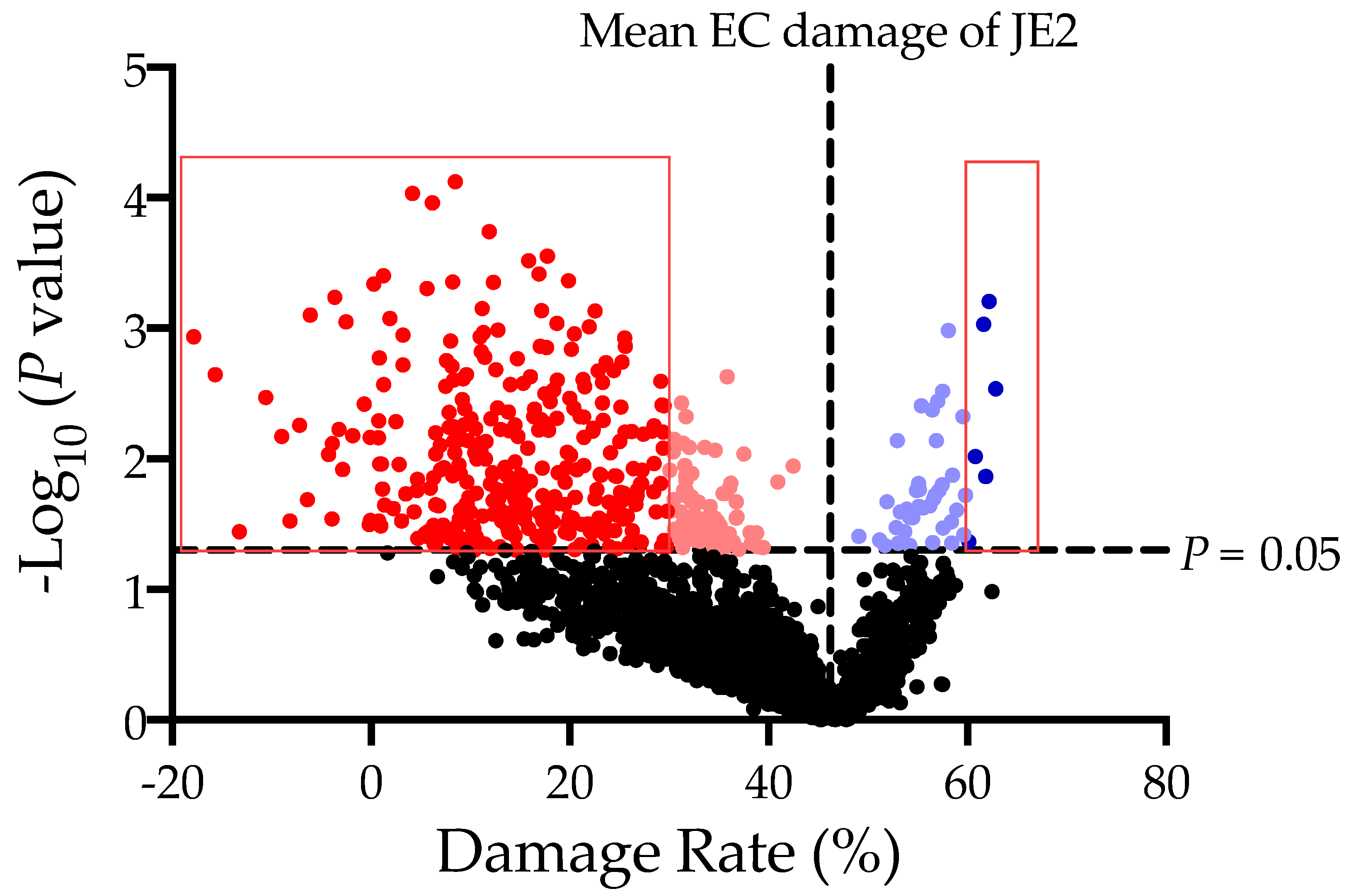
2.1. The MTT Assay Is Applicable to the High Throughput Screening of Genes Involved in HMEC-1 Damage
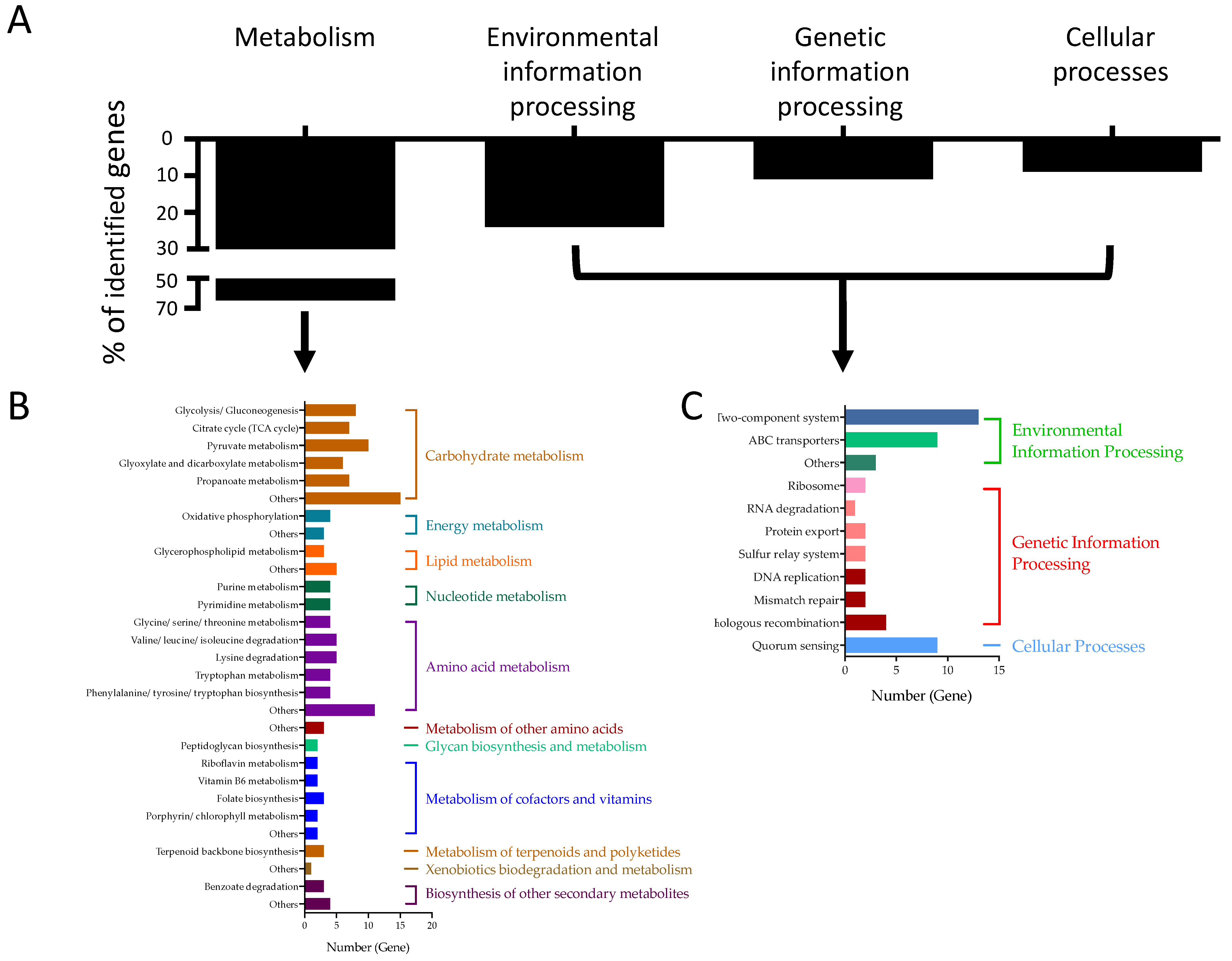
2.2. Identified Staphylococcal Genes Impacting HMEC-1 Damages
3. Discussion
4. Materials and Methods
4.1. Bacteria and Growth Conditions
4.2. Endothelial Cell (HMEC-1) Culture
4.3. HMEC-1 Damage Assay
4.4. Verification of the HMEC-1 Damage Screening Results
4.5. Statistical Analysis
4.6. KEGG Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fowler, V.G., Jr.; Miro, J.M.; Hoen, B.; Cabell, C.H.; Abrutyn, E.; Rubinstein, E.; Corey, G.R.; Spelman, D.; Bradley, S.F.; Barsic, B.; et al. Staphylococcus aureus endocarditis: A consequence of medical progress. JAMA 2005, 293, 3012–3021. [Google Scholar] [CrossRef]
- Cheung, A.L.; Bayer, A.S.; Zhang, G.; Gresham, H.; Xiong, Y.Q. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2004, 40, 1–9. [Google Scholar] [CrossRef]
- Ondusko, D.S.; Nolt, D. Staphylococcus aureus. Pediatr. Rev. 2018, 39, 287–298. [Google Scholar] [CrossRef]
- Chorianopoulos, E.; Bea, F.; Katus, H.A.; Frey, N. The role of endothelial cell biology in endocarditis. Cell Tissue Res. 2009, 335, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Bayer, A.S.; McKinnell, J.A.; Ellison, S.; Filler, S.G.; Xiong, Y.Q. In vitro endothelial cell damage is positively correlated with enhanced virulence and poor vancomycin responsiveness in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Cell Microbiol. 2011, 13, 1530–1541. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.D.; Endres, J.L.; Yajjala, V.K.; Widhelm, T.J.; Boissy, R.J.; Bose, J.L.; Bayles, K.W. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. mBio 2013, 4, e00537-12. [Google Scholar] [CrossRef] [PubMed]
- Paudel, A.; Hamamoto, H.; Panthee, S.; Matsumoto, Y.; Sekimizu, K. Large-Scale Screening and Identification of Novel Pathogenic Staphylococcus aureus Genes Using a Silkworm Infection Model. J. Infect. Dis. 2020, 221, 1795–1804. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.L.; Fey, P.D.; Bayles, K.W. Genetic tools to enhance the study of gene function and regulation in Staphylococcus aureus. Appl. Environ. Microbiol. 2013, 79, 2218–2224. [Google Scholar] [CrossRef]
- Ades, E.W.; Candal, F.J.; Swerlick, R.A.; George, V.G.; Summers, S.; Bosse, D.C.; Lawley, T.J. HMEC-1: Establishment of an immortalized human microvascular endothelial cell line. J. Investig. Dermatol. 1992, 99, 683–690. [Google Scholar] [CrossRef]
- Seidl, K.; Solis, N.V.; Bayer, A.S.; Hady, W.A.; Ellison, S.; Klashman, M.C.; Xiong, Y.Q.; Filler, S.G. Divergent responses of different endothelial cell types to infection with Candida albicans and Staphylococcus aureus. PLoS ONE 2012, 7, e39633. [Google Scholar] [CrossRef]
- Luo, X.; Zhang, X.; Zhao, T.; Zeng, T.; Liu, W.; Deng, M.; Zhao, F. A preliminary study on the proinflammatory mechanisms of Treponema pallidum outer membrane protein Tp92 in human macrophages and HMEC-1 cells. Microb. Pathog. 2017, 110, 176–183. [Google Scholar] [CrossRef]
- Ferrero, M.C.; Bregante, J.; Delpino, M.V.; Barrionuevo, P.; Fossati, C.A.; Giambartolomei, G.H.; Baldi, P.C. Proinflammatory response of human endothelial cells to Brucella infection. Microbes Infect. 2011, 13, 852–861. [Google Scholar] [CrossRef]
- Seidl, K.; Leemann, M.; Zinkernagel, A.S. The ArlRS two-component system is a regulator of Staphylococcus aureus-induced endothelial cell damage. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Bayer, A.S.; Fowler, V.G., Jr.; McKinnell, J.A.; Abdel Hady, W.; Sakoulas, G.; Yeaman, M.R.; Xiong, Y.Q. Combinatorial phenotypic signatures distinguish persistent from resolving methicillin-resistant Staphylococcus aureus bacteremia isolates. Antimicrob. Agents Chemother. 2011, 55, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bayer, A.S.; Cheung, A.; Lu, L.; Abdelhady, W.; Donegan, N.P.; Hong, J.I.; Yeaman, M.R.; Xiong, Y.Q. The Stringent Response Contributes to Persistent Methicillin-Resistant Staphylococcus aureus Endovascular Infection Through the Purine Biosynthetic Pathway. J. Infect. Dis. 2020, 222, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Haslinger-Loffler, B.; Kahl, B.C.; Grundmeier, M.; Strangfeld, K.; Wagner, B.; Fischer, U.; Cheung, A.L.; Peters, G.; Schulze-Osthoff, K.; Sinha, B. Multiple virulence factors are required for Staphylococcus aureus-induced apoptosis in endothelial cells. Cell Microbiol. 2005, 7, 1087–1097. [Google Scholar] [CrossRef]
- Braungardt, H.; Singh, V.K. Impact of Deficiencies in Branched-Chain Fatty Acids and Staphyloxanthin in Staphylococcus aureus. Biomed. Res. Int. 2019, 2019, 2603435. [Google Scholar] [CrossRef]
- Singh, V.K.; Sirobhushanam, S.; Ring, R.P.; Singh, S.; Gatto, C.; Wilkinson, B.J. Roles of pyruvate dehydrogenase and branched-chain alpha-keto acid dehydrogenase in branched-chain membrane fatty acid levels and associated functions in Staphylococcus aureus. J. Med. Microbiol. 2018, 67, 570–578. [Google Scholar] [CrossRef]
- Campbell, C.; Fingleton, C.; Zeden, M.S.; Bueno, E.; Gallagher, L.A.; Shinde, D.; Ahn, J.; Olson, H.M.; Fillmore, T.L.; Adkins, J.N.; et al. Accumulation of Succinyl Coenzyme A Perturbs the Methicillin-Resistant Staphylococcus aureus (MRSA) Succinylome and Is Associated with Increased Susceptibility to Beta-Lactam Antibiotics. mBio 2021, 12, e0053021. [Google Scholar] [CrossRef]
- Stevens, E.; Laabei, M.; Gardner, S.; Somerville, G.A.; Massey, R.C. Cytolytic toxin production by Staphylococcus aureus is dependent upon the activity of the protoheme IX farnesyltransferase. Sci. Rep. 2017, 7, 13744. [Google Scholar] [CrossRef]
- Turner, A.K.; Barber, L.Z.; Wigley, P.; Muhammad, S.; Jones, M.A.; Lovell, M.A.; Hulme, S.; Barrow, P.A. Contribution of proton-translocating proteins to the virulence of Salmonella enterica serovars Typhimurium, Gallinarum, and Dublin in chickens and mice. Infect. Immun. 2003, 71, 3392–3401. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Luqman, A.; Bitschar, K.; Hertlein, T.; Dick, J.; Ohlsen, K.; Broker, B.; Schittek, B.; Gotz, F. Staphylococcal (phospho)lipases promote biofilm formation and host cell invasion. Int. J. Med. Microbiol. 2018, 308, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Sahl, H.G. An oldie but a goodie—Cell wall biosynthesis as antibiotic target pathway. Int. J. Med. Microbiol. 2010, 300, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Bugg, T.D. Recent advances in antimicrobial nucleoside antibiotics targeting cell wall biosynthesis. Nat. Prod. Rep. 2003, 20, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Ho, Y.X.; Cowell, L.M.; Jilani, I.; Foster, S.J.; Prince, L.R. A Genome-Wide Screen Identifies Factors Involved in S. aureus-Induced Human Neutrophil Cell Death and Pathogenesis. Front. Immunol. 2019, 10, 45. [Google Scholar] [CrossRef]
- Cabrera, G.; Xiong, A.; Uebel, M.; Singh, V.K.; Jayaswal, R.K. Molecular characterization of the iron-hydroxamate uptake system in Staphylococcus aureus. Appl. Environ. Microbiol. 2001, 67, 1001–1003. [Google Scholar] [CrossRef]
- Handke, L.D.; Gribenko, A.V.; Timofeyeva, Y.; Scully, I.L.; Anderson, A.S. MntC-Dependent Manganese Transport Is Essential for Staphylococcus aureus Oxidative Stress Resistance and Virulence. mSphere 2018, 3, e00336-18. [Google Scholar] [CrossRef]
- Munzenmayer, L.; Geiger, T.; Daiber, E.; Schulte, B.; Autenrieth, S.E.; Fraunholz, M.; Wolz, C. Influence of Sae-regulated and Agr-regulated factors on the escape of Staphylococcus aureus from human macrophages. Cell Microbiol. 2016, 18, 1172–1183. [Google Scholar] [CrossRef]
- Atwood, D.N.; Beenken, K.E.; Loughran, A.J.; Meeker, D.G.; Lantz, T.L.; Graham, J.W.; Spencer, H.J.; Smeltzer, M.S. XerC Contributes to Diverse Forms of Staphylococcus aureus Infection via agr-Dependent and agr-Independent Pathways. Infect. Immun. 2016, 84, 1214–1225. [Google Scholar] [CrossRef]
- Mashruwala, A.A.; Pang, Y.Y.; Rosario-Cruz, Z.; Chahal, H.K.; Benson, M.A.; Mike, L.A.; Skaar, E.P.; Torres, V.J.; Nauseef, W.M.; Boyd, J.M. Nfu facilitates the maturation of iron-sulfur proteins and participates in virulence in Staphylococcus aureus. Mol. Microbiol. 2015, 95, 383–409. [Google Scholar] [CrossRef]
- Austin, C.M.; Garabaglu, S.; Krute, C.N.; Ridder, M.J.; Seawell, N.A.; Markiewicz, M.A.; Boyd, J.M.; Bose, J.L. Contribution of YjbIH to Virulence Factor Expression and Host Colonization in Staphylococcus aureus. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Baba, T.; Sekine, M.; Fukuda, M.; Hiramatsu, K. Beta-hemolysin promotes skin colonization by Staphylococcus aureus. J. Bacteriol. 2013, 195, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Kaatz, G.W.; McAleese, F.; Seo, S.M. Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob. Agents Chemother. 2005, 49, 1857–1864. [Google Scholar] [CrossRef]
- Liu, G.; Zhou, J.; Fu, Q.S.; Wang, J. The Escherichia coli azoreductase AzoR Is involved in resistance to thiol-specific stress caused by electrophilic quinones. J. Bacteriol. 2009, 191, 6394–6400. [Google Scholar] [CrossRef]
- Daniels, J.N.; Wuebbens, M.M.; Rajagopalan, K.V.; Schindelin, H. Crystal structure of a molybdopterin synthase-precursor Z complex: Insight into its sulfur transfer mechanism and its role in molybdenum cofactor deficiency. Biochemistry 2008, 47, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Vriesema, A.J.; Beekhuizen, H.; Hamdi, M.; Soufan, A.; Lammers, A.; Willekens, B.; Bakker, O.; Welten, A.G.; Veltrop, M.H.; van De Gevel, J.S.; et al. Altered gene expression in Staphylococcus aureus upon interaction with human endothelial cells. Infect. Immun. 2000, 68, 1765–1772. [Google Scholar] [CrossRef]
- Seidl, K.; Zinkernagel, A.S. The MTT assay is a rapid and reliable quantitative method to assess Staphylococcus aureus induced endothelial cell damage. J. Microbiol. Methods 2013, 92, 307–309. [Google Scholar] [CrossRef]
- Quiblier, C.; Seidl, K.; Roschitzki, B.; Zinkernagel, A.S.; Berger-Bachi, B.; Senn, M.M. Secretome analysis defines the major role of SecDF in Staphylococcus aureus virulence. PLoS ONE 2013, 8, e63513. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locus | Gene Name | Description | % EC Damage (Mean ± SD) |
|---|---|---|---|
| SAUSA300_0261 | hypothetical | conserved hypothetical protein | 29.83 ± 8.34 |
| SAUSA300_1172 | hypothetical | M16 family peptidase | 29.74 ± 4.80 |
| SAUSA300_0083 | hypothetical | hypothetical protein | 29.70 ± 10.14 |
| SAUSA300_1386 | hypothetical | phiETA ORF59-like protein | 29.57 ± 1.07 |
| SAUSA300_0076 | hypothetical | ABC transporter ATP-binding protein | 29.57 ± 4.10 |
| SAUSA300_1712 | ribH | 6,7-dimethyl-8-ribityllumazine synthase | 29.49 ± 9.83 |
| SAUSA300_1457 | malR | maltose operon transcriptional repressor | 29.46 ± 2.79 |
| SAUSA300_1309 | hypothetical | IS200 family transposase | 29.41 ± 8.13 |
| SAUSA300_1253 | glcT | transcription antiterminator | 29.37 ± 4.04 |
| SAUSA300_1797 | hypothetical | conserved hypothetical protein | 29.37 ± 4.79 |
| SAUSA300_1759 | hypothetical | hypothetical protein | 29.25 ± 2.85 |
| SAUSA300_2386 | hypothetical | beta-lactamase | 29.13 ± 1.62 |
| SAUSA300_2434 | hypothetical | transporter protein | 29.13 ± 5.28 |
| SAUSA300_2037 | hypothetical | ATP-dependent RNA helicase | 28.67 ± 8.90 |
| SAUSA300_1654 | hypothetical | proline dipeptidase | 28.46 ± 4.20 |
| SAUSA300_0615 | hypothetical | putative monovalent cation/H+ antiporter subunit F | 28.45 ± 4.24 |
| SAUSA300_1659 | tpx | thiol peroxidase | 28.41 ± 7.42 |
| SAUSA300_1478 | hypothetical | putative lipoprotein | 28.28 ± 4.37 |
| SAUSA300_2455 | hypothetical | putative fructose-1,6-bisphosphatase | 28.27 ± 5.83 |
| SAUSA300_1297 | acyP | acylphosphatase | 28.23 ± 4.50 |
| SAUSA300_2606 | hisF | imidazole glycerol phosphate synthase subunit HisF | 27.62 ± 4.01 |
| SAUSA300_0795 | hypothetical | hypothetical protein | 27.38 ± 6.00 |
| SAUSA300_1683 | hypothetical | bifunctional 3-deoxy-7-phosphoheptulonate synthase/chorismate mutase | 27.26 ± 6.86 |
| SAUSA300_2618 | hypothetical | hypothetical protein | 27.23 ± 7.65 |
| SAUSA300_1398 | hypothetical | phiSLT ORF123-like protein | 27.16 ± 11.43 |
| SAUSA300_0059 | hypothetical | conserved hypothetical protein | 27.07 ± 7.67 |
| SAUSA300_1764 | epiD | lantibiotic epidermin biosynthesis protein EpiD | 26.84 ± 3.46 |
| SAUSA300_2332 | hypothetical | heat shock protein | 26.78 ± 8.46 |
| SAUSA300_1040 | hypothetical | hypothetical protein | 26.74 ± 8.21 |
| SAUSA300_2280 | fosB | fosfomycin resistance protein FosB | 26.67 ± 8.68 |
| SAUSA300_1750 | hypothetical | conserved hypothetical protein | 26.62 ± 9.44 |
| SAUSA300_0883 | hypothetical | putative surface protein | 26.40 ± 12.90 |
| SAUSA300_1964 | hypothetical | hypothetical protein | 26.38 ± 7.19 |
| SAUSA300_0290 | hypothetical | putative lipoprotein | 26.29 ± 8.56 |
| SAUSA300_1672 | nagE | phosphotransferase system, N-acetylglucosamine-specific IIBC component | 26.21 ± 5.46 |
| SAUSA300_2023 | rsbW | anti-sigma-B factor, serine-protein kinase | 26.01 ± 0.14 |
| SAUSA300_0190 | ipdC | indole-3-pyruvate decarboxylase | 25.81 ± 7.93 |
| SAUSA300_2413 | hypothetical | hypothetical protein | 25.79 ± 4.70 |
| SAUSA300_0798 | hypothetical | ABC transporter substrate-binding protein | 25.59 ± 3.93 |
| SAUSA300_0489 | ftsH | putative cell division protein FtsH | 25.55 ± 5.76 |
| SAUSA300_1093 | pyrB | aspartate carbamoyltransferase catalytic subunit | 25.49 ± 1.23 |
| SAUSA300_0517 | hypothetical | RNA methyltransferase | 25.39 ± 8.18 |
| SAUSA300_1740 | hypothetical | hypothetical protein | 25.37 ± 9.05 |
| SAUSA300_0540 | hypothetical | HAD family hydrolase | 25.26 ± 9.24 |
| SAUSA300_2272 | hypothetical | hypothetical protein | 25.25 ± 4.80 |
| SAUSA300_1968 | hypothetical | putative phage transcriptional regulator | 25.23 ± 9.97 |
| SAUSA300_0642 | hypothetical | hypothetical protein | 25.21 ± 4.58 |
| SAUSA300_2358 | hypothetical | ABC transporter permease | 25.11 ± 6.08 |
| SAUSA300_1984 | mroQ | hypothetical protein | 25.07 ± 9.15 |
| SAUSA300_1266 | trpF | N-(5′-phosphoribosyl)anthranilate isomerase | 25.05 ± 7.12 |
| SAUSA300_2251 | hypothetical | dehydrogenase family protein | 25.00 ± 3.65 |
| SAUSA300_0706 | hypothetical | putative osmoprotectant ABC transporter ATP-binding protein | 24.95 ± 11.00 |
| SAUSA300_0941 | hypothetical | putative ferrichrome ABC transporter | 24.69 ± 6.43 |
| SAUSA300_0951 | sspA | V8 protease | 24.55 ± 8.41 |
| SAUSA300_1875 | hypothetical | exonuclease | 24.52 ± 10.68 |
| SAUSA300_0566 | hypothetical | amino acid permease | 24.49 ± 5.06 |
| SAUSA300_0871 | hypothetical | hypothetical protein | 24.49 ± 12.19 |
| SAUSA300_0565 | hypothetical | conserved hypothetical protein | 24.43 ± 5.34 |
| SAUSA300_0391 | hypothetical | hypothetical protein | 24.38 ± 0.45 |
| SAUSA300_1328 | hypothetical | putative drug transporter | 24.10 ± 7.38 |
| SAUSA300_2279 | hypothetical | LysR family regulatory protein | 23.92 ± 10.37 |
| SAUSA300_0505 | hypothetical | glutamine amidotransferase subunit PdxT | 23.61 ± 3.46 |
| SAUSA300_0470 | ksgA | dimethyladenosine transferase | 23.56 ± 7.13 |
| SAUSA300_1106 | hypothetical | putative lipoprotein | 23.45 ± 8.92 |
| SAUSA300_1991 | agrC | accessory gene regulator protein C | 23.44 ± 9.71 |
| SAUSA300_0108 | hypothetical | antigen, 67 kDa | 23.33 ± 6.80 |
| SAUSA300_2326 | araC | transcription regulatory protein | 23.30 ± 5.35 |
| SAUSA300_1399 | hypothetical | phiSLT ORF110-like protein | 23.29 ± 0.65 |
| SAUSA300_1942 | hypothetical | hypothetical protein | 23.29 ± 11.27 |
| SAUSA300_0079 | hypothetical | putative lipoprotein | 23.27 ± 6.02 |
| SAUSA300_1384 | hypothetical | phiSLT ORF100b-like protein, holin | 23.25 ± 6.98 |
| SAUSA300_1950 | hypothetical | hypothetical protein | 23.24 ± 9.64 |
| SAUSA300_0320 | gehB | triacylglycerol lipase | 23.13 ± 9.02 |
| SAUSA300_0370 | hypothetical | putative enterotoxin | 23.06 ± 9.01 |
| SAUSA300_1224 | hypothetical | conserved hypothetical protein | 22.85 ± 4.12 |
| SAUSA300_1925 | hypothetical | phiPVL ORF17-like protein | 22.72 ± 9.85 |
| SAUSA300_1271 | hypothetical | hydrolase-like protein | 22.57 ± 5.67 |
| SAUSA300_0547 | sdrD | sdrD protein | 22.52 ± 1.23 |
| SAUSA300_0561 | hypothetical | hypothetical protein | 22.37 ± 6.87 |
| SAUSA300_2367 | hlgB | gamma-hemolysin component B | 22.27 ± 7.70 |
| SAUSA300_1671 | hypothetical | hypothetical protein | 22.15 ± 10.08 |
| SAUSA300_2341 | narJ | respiratory nitrate reductase, subunit delta | 22.11 ± 4.50 |
| SAUSA300_0420 | hypothetical | hypothetical protein | 22.10 ± 8.19 |
| SAUSA300_2281 | hutG | formimidoylglutamase | 22.05 ± 12.63 |
| SAUSA300_1427 | hypothetical | phiSLT ORF86-like protein | 21.94 ± 2.49 |
| SAUSA300_0691 | saeR | DNA-binding response regulator SaeR | 21.93 ± 10.56 |
| SAUSA300_1519 | hypothetical | hypothetical protein | 21.86 ± 0.84 |
| SAUSA300_0253 | scdA | cell wall biosynthesis protein ScdA | 21.83 ± 12.24 |
| SAUSA300_2459 | hypothetical | MarR family transcriptional regulator | 21.58 ± 6.37 |
| SAUSA300_2505 | hypothetical | acetyltransferase | 21.48 ± 5.28 |
| SAUSA300_0652 | hypothetical | hypothetical protein | 21.46 ± 9.86 |
| SAUSA300_1213 | hypothetical | hypothetical protein | 21.42 ± 8.18 |
| SAUSA300_1216 | hypothetical | cardiolipin synthetase | 21.40 ± 13.46 |
| SAUSA300_0395 | hypothetical | superantigen-like protein | 21.39 ± 9.28 |
| SAUSA300_1016 | cyoE | protoheme IX farnesyltransferase | 21.38 ± 6.70 |
| SAUSA300_1126 | rnc | ribonuclease III | 21.34 ± 5.04 |
| SAUSA300_1437 | hypothetical | phiSLT ORF204-like protein | 21.26 ± 3.02 |
| SAUSA300_2145 | hypothetical | glycine betaine transporter | 21.18 ± 9.85 |
| SAUSA300_2288 | hypothetical | ABC transporter ATP-binding protein | 21.10 ± 15.49 |
| SAUSA300_0698 | pabA | para-aminobenzoate synthase, glutamine amidotransferase, component II | 21.05 ± 4.75 |
| SAUSA300_0519 | hypothetical | hypothetical protein | 20.86 ± 6.93 |
| SAUSA300_2330 | hypothetical | hypothetical protein | 20.82 ± 4.02 |
| SAUSA300_0141 | deoB | phosphopentomutase | 20.69 ± 9.71 |
| SAUSA300_1684 | hypothetical | hypothetical protein | 20.53 ± 11.18 |
| SAUSA300_1595 | tgt | queuine tRNA-ribosyltransferase | 20.53 ± 9.07 |
| SAUSA300_0442 | hypothetical | hypothetical protein | 20.45 ± 3.70 |
| SAUSA300_0744 | lgt | prolipoprotein diacylglyceryl transferase | 20.44 ± 5.61 |
| SAUSA300_1576 | recD2 | helicase, RecD/TraA family | 20.41 ± 6.63 |
| SAUSA300_2088 | luxS | S-ribosylhomocysteinase | 20.40 ± 2.33 |
| SAUSA300_0131 | hypothetical | putative Bacterial sugar transferase | 20.28 ± 13.49 |
| SAUSA300_0649 | hypothetical | hypothetical protein | 20.23 ± 0.89 |
| SAUSA300_2550 | nrdG | anaerobic ribonucleotide reductase, small subunit | 20.22 ± 10.12 |
| SAUSA300_2168 | hypothetical | hypothetical protein | 20.16 ± 4.12 |
| SAUSA300_2587 | hypothetical | accessory secretory protein Asp1 | 20.06 ± 9.42 |
| SAUSA300_2548 | hypothetical | hypothetical protein | 19.98 ± 7.37 |
| SAUSA300_1021 | hypothetical | hypothetical protein | 19.92 ± 15.09 |
| SAUSA300_0456 | rrlA | 23S ribosomal RNA | 19.91 ± 0.15 |
| SAUSA300_0431 | hypothetical | hypothetical protein | 19.86 ± 4.23 |
| SAUSA300_1247 | hypothetical | conserved hypothetical protein | 19.79 ± 10.23 |
| SAUSA300_2108 | mtlD | mannitol-1-phosphate 5-dehydrogenase | 19.74 ± 9.18 |
| SAUSA300_2516 | hypothetical | short chain dehydrogenase/reductase family oxidoreductase | 19.65 ± 10.14 |
| SAUSA300_0450 | treR | trehalose operon repressor | 19.59 ± 13.38 |
| SAUSA300_0422 | hypothetical | hypothetical protein | 19.54 ± 2.66 |
| SAUSA300_1739 | hypothetical | hypothetical protein | 19.47 ± 8.56 |
| SAUSA300_0257 | lrgB | antiholin-like protein LrgB | 19.47 ± 17.61 |
| SAUSA300_0056 | hypothetical | hypothetical protein | 19.05 ± 4.22 |
| SAUSA300_2352 | hypothetical | addiction module antitoxin | 18.95 ± 11.82 |
| SAUSA300_2236 | hypothetical | hypothetical protein | 18.82 ± 4.26 |
| SAUSA300_1409 | hypothetical | hypothetical protein | 18.77 ± 11.78 |
| SAUSA300_1304 | hypothetical | hypothetical protein | 18.73 ± 5.92 |
| SAUSA300_1934 | hypothetical | phi77 ORF020-like protein, phage major tail protein | 18.68 ± 3.51 |
| SAUSA300_1279 | phoU | phosphate transport system regulatory protein PhoU | 18.68 ± 7.74 |
| SAUSA300_1217 | hypothetical | ABC transporter ATP-binding protein | 18.66 ± 8.42 |
| SAUSA300_0468 | hypothetical | TatD family hydrolase | 18.62 ± 0.90 |
| SAUSA300_2132 | hypothetical | hypothetical protein | 18.54 ± 17.28 |
| SAUSA300_0288 | essD/esaD | hypothetical protein | 18.50 ± 12.03 |
| SAUSA300_2461 | hypothetical | glyoxalase family protein | 18.38 ± 6.48 |
| SAUSA300_1349 | bshA | glycosyl transferase, group 1 family protein | 18.26 ± 11.03 |
| SAUSA300_1009 | typA | GTP-binding protein | 18.22 ± 6.42 |
| SAUSA300_1755 | splD | serine protease SplD | 18.20 ± 6.01 |
| SAUSA300_1966 | hypothetical | phi77 ORF014-like protein, phage anti-repressor protein | 18.04 ± 5.61 |
| SAUSA300_1307 | arlS | sensor histidine kinase protein | 18.01 ± 7.14 |
| SAUSA300_1918 | hlb | truncated beta-hemolysin | 17.91 ± 11.34 |
| SAUSA300_1569 | hypothetical | U32 family peptidase | 17.90 ± 6.37 |
| SAUSA300_1397 | hypothetical | phiSLT ORF213-like protein, major tail protein | 17.88 ± 16.40 |
| SAUSA300_1032 | hypothetical | putative iron compound ABC transporter iron compound-binding protein | 17.87 ± 9.01 |
| SAUSA300_0259 | hypothetical | PTS system, IIA component | 17.72 ± 4.08 |
| SAUSA300_1070 | hypothetical | hypothetical protein | 17.66 ± 6.61 |
| SAUSA300_1474 | hypothetical | hypothetical protein | 17.57 ± 3.84 |
| SAUSA300_1451 | hypothetical | short chain dehydrogenase/reductase family oxidoreductase | 17.47 ± 4.46 |
| SAUSA300_0769 | hypothetical | hypothetical protein | 17.42 ± 7.43 |
| SAUSA300_2098 | arsR | ArsR family transcriptional regulator | 17.36 ± 8.42 |
| SAUSA300_0094 | hypothetical | hypothetical protein | 17.32 ± 9.77 |
| SAUSA300_1470 | ispA | geranyltranstransferase | 17.29 ± 13.19 |
| SAUSA300_1403 | hypothetical | phiSLT ORF412-like protein, portal protein | 17.28 ± 10.80 |
| SAUSA300_2432 | hypothetical | MutT/NUDIX family hydrolase | 17.26 ± 15.82 |
| SAUSA300_0631 | hypothetical | putative nucleoside transporter | 17.25 ± 11.20 |
| SAUSA300_1000 | potB | spermidine/putrescine ABC transporter permease | 17.14 ± 5.86 |
| SAUSA300_2559 | hypothetical | DNA-binding response regulator | 17.10 ± 8.85 |
| SAUSA300_2467 | srtA | sortase | 17.01 ± 6.72 |
| SAUSA300_2300 | hypothetical | transcriptional regulator, TetR family | 16.92 ± 5.04 |
| SAUSA300_0916 | hypothetical | hypothetical protein | 16.89 ± 2.85 |
| SAUSA300_1444 | scpB | segregation and condensation protein B | 16.85 ± 6.40 |
| SAUSA300_0995 | hypothetical | branched-chain alpha-keto acid dehydrogenase subunit E2 | 16.83 ± 18.68 |
| SAUSA300_0419 | hypothetical | tandem lipoprotein | 16.78 ± 3.58 |
| SAUSA300_1563 | accC | acetyl-CoA carboxylase, biotin carboxylase | 16.73 ± 11.04 |
| SAUSA300_2027 | alr | alanine racemase | 16.70 ± 16.05 |
| SAUSA300_2607 | hisA | phoribosyl)-5-((5-phosphoribosylamino)methylideneamino) imidazole-4-carboxamide | 16.70 ± 11.46 |
| SAUSA300_0023 | hypothetical | hypothetical protein | 16.69 ± 16.09 |
| SAUSA300_1622 | tig | trigger factor | 16.44 ± 5.67 |
| SAUSA300_0011 | hypothetical | hypothetical protein | 16.37 ± 4.02 |
| SAUSA300_1097 | pyrF | orotidine 5′-phosphate decarboxylase | 16.34 ± 8.94 |
| SAUSA300_1339 | hypothetical | hypothetical protein | 16.25 ± 5.49 |
| SAUSA300_0585 | hypothetical | hypothetical protein | 16.24 ± 13.38 |
| SAUSA300_0839 | nfu | hypothetical protein | 16.23 ± 12.30 |
| SAUSA300_0071 | hypothetical | ISSep1-like transposase | 16.19 ± 3.17 |
| SAUSA300_0651 | hypothetical | CHAP domain-contain protein | 16.09 ± 6.91 |
| SAUSA300_1599 | hypothetical | hypothetical protein | 16.02 ± 7.75 |
| SAUSA300_1607 | hypothetical | hypothetical protein | 16.02 ± 8.76 |
| SAUSA300_0588 | hypothetical | hypothetical protein | 15.86 ± 15.72 |
| SAUSA300_2276 | hypothetical | peptidase, M20/M25/M40 family | 15.84 ± 1.33 |
| SAUSA300_2055 | murA | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | 15.79 ± 10.49 |
| SAUSA300_0808 | hypothetical | hypothetical protein | 15.69 ± 12.88 |
| SAUSA300_0759 | gpmI | phosphoglyceromutase | 15.68 ± 9.84 |
| SAUSA300_0857 | ppiB | hypothetical protein | 15.66 ± 4.76 |
| SAUSA300_1051 | hypothetical | hypothetical protein | 15.51 ± 14.05 |
| SAUSA300_1383 | hypothetical | phiSLT ORF484-like protein, lysin | 15.46 ± 15.13 |
| SAUSA300_1566 | hypothetical | hypothetical protein | 15.42 ± 14.25 |
| SAUSA300_2040 | hypothetical | hypothetical protein | 15.42 ± 12.63 |
| SAUSA300_1145 | xerC | tyrosine recombinase xerC | 15.33 ± 4.57 |
| SAUSA300_0687 | hypothetical | putative hemolysin | 15.14 ± 12.23 |
| SAUSA300_0630 | hypothetical | ABC transporter ATP-binding protein | 15.07 ± 10.45 |
| SAUSA300_1577 | hypothetical | TPR domain-containing protein | 14.93 ± 1.75 |
| SAUSA300_1288 | dapA | dihydrodipicolinate synthase | 14.75 ± 7.53 |
| SAUSA300_1937 | hypothetical | phi77 ORF045-like protein | 14.69 ± 8.83 |
| SAUSA300_1419 | hypothetical | phiSLT ORF80-like protein | 14.65 ± 9.06 |
| SAUSA300_2345 | nirD | nitrite reductase (NAD(P)H), small subunit | 14.54 ± 4.64 |
| SAUSA300_1365 | rpsA | 30S ribosomal protein S1 | 14.53 ± 3.46 |
| SAUSA300_0029 | hypothetical | hypothetical protein | 14.39 ± 3.30 |
| SAUSA300_2575 | hypothetical | BglG family transcriptional antiterminator | 14.12 ± 4.67 |
| SAUSA300_1497 | hypothetical | glycine dehydrogenase subunit 1 | 14.08 ± 4.09 |
| SAUSA300_1682 | ccpA | catabolite control protein A | 14.04 ± 8.43 |
| SAUSA300_0657 | hypothetical | hypothetical protein | 14.02 ± 7.45 |
| SAUSA300_1955 | hypothetical | putative endodeoxyribonuclease RusA | 13.92 ± 10.12 |
| SAUSA300_0924 | ktrD | sodium transport family protein | 13.85 ± 14.78 |
| SAUSA300_0077 | hypothetical | ABC transporter ATP-binding protein | 13.80 ± 6.67 |
| SAUSA300_0504 | pdxS | pyridoxal biosynthesis lyase PdxS | 13.58 ± 7.70 |
| SAUSA300_0195 | hypothetical | transcriptional regulator | 13.06 ± 13.37 |
| SAUSA300_1308 | arlR | DNA-binding response regulator | 13.05 ± 5.02 |
| SAUSA300_0859 | hypothetical | NADH-dependent flavin oxidoreductase | 12.99 ± 7.37 |
| SAUSA300_1721 | hypothetical | hypothetical protein | 12.97 ± 3.93 |
| SAUSA300_0186 | argC | N-acetyl-gamma-glutamyl-phosphate reductase | 12.92 ± 16.00 |
| SAUSA300_2641 | hypothetical | hypothetical protein | 12.90 ± 8.36 |
| SAUSA300_0987 | hypothetical | cytochrome D ubiquinol oxidase, subunit II | 12.85 ± 10.22 |
| SAUSA300_1696 | dat | D-alanine aminotransferase | 12.74 ± 5.48 |
| SAUSA300_1283 | hypothetical | phosphate ABC transporter, phosphate-binding protein PstS | 12.73 ± 9.23 |
| SAUSA300_1185 | miaB | (dimethylallyl)adenosine tRNA methylthiotransferase | 12.62 ± 10.40 |
| SAUSA300_2365 | hlgA | gamma-hemolysin component A | 12.56 ± 10.54 |
| SAUSA300_1394 | hypothetical | hypothetical protein | 12.34 ± 12.26 |
| SAUSA300_0115 | sirC | iron compound ABC transporter permease SirC | 12.30 ± 6.17 |
| SAUSA300_2284 | hypothetical | hypothetical protein | 12.20 ± 10.36 |
| SAUSA300_2225 | moaC | molybdenum cofactor biosynthesis protein MoaC | 12.08 ± 9.05 |
| SAUSA300_0244 | hypothetical | zinc-binding dehydrogenase family oxidoreductase | 12.05 ± 9.79 |
| SAUSA300_2022 | rpoF | RNA polymerase sigma factor SigB | 12.05 ± 6.83 |
| SAUSA300_1089 | lspA | lipoprotein signal peptidase | 11.97 ± 6.81 |
| SAUSA300_1618 | hemX | hemA concentration negative effector hemX | 11.88 ± 1.05 |
| SAUSA300_0117 | sirA | iron compound ABC transporter iron compound-binding protein SirA | 11.83 ± 7.84 |
| SAUSA300_0899 | mecA | adaptor protein | 11.58 ± 10.37 |
| SAUSA300_2492 | hypothetical | acetyltransferase family protein | 11.55 ± 7.80 |
| SAUSA300_1433 | hypothetical | putative phage regulatory protein | 11.41 ± 8.17 |
| SAUSA300_1244 | mscL | large conductance mechanosensitive channel protein | 11.32 ± 7.21 |
| SAUSA300_0049 | hypothetical | hypothetical protein | 11.30 ± 0.62 |
| SAUSA300_1667 | hypothetical | putative glycerophosphoryl diester phosphodiesterase | 11.30 ± 7.51 |
| SAUSA300_0994 | pdhB | pyruvate dehydrogenase E1 component, beta subunit | 11.20 ± 8.12 |
| SAUSA300_0974 | purN | phosphoribosylglycinamide formyltransferase | 11.07 ± 8.08 |
| SAUSA300_0067 | hypothetical | universal stress protein | 11.02 ± 9.02 |
| SAUSA300_1590 | rsh (relA) | GTP pyrophosphokinase | 10.95 ± 7.18 |
| SAUSA300_0526 | hypothetical | methyltransferase small subunit | 10.80 ± 10.78 |
| SAUSA300_0952 | hypothetical | aminotransferase, class I | 10.57 ± 6.79 |
| SAUSA300_1694 | trmB | tRNA (guanine-N(7)-)-methyltransferase | 10.55 ± 16.08 |
| SAUSA300_0041 | hypothetical | hypothetical protein | 10.41 ± 2.09 |
| SAUSA300_1449 | hypothetical | MutT/nudix family protein | 10.11 ± 13.24 |
| SAUSA300_0724 | hypothetical | hypothetical protein | 10.06 ± 2.60 |
| SAUSA300_1757 | splB | serine protease SplB | 9.41 ± 4.17 |
| SAUSA300_0476 | hypothetical | hypothetical protein | 9.18 ± 8.05 |
| SAUSA300_2052 | hypothetical | single-stranded DNA- binding protein family | 9.11 ± 18.19 |
| SAUSA300_2176 | cbiO | cobalt transporter ATP-binding subunit | 9.03 ± 9.11 |
| SAUSA300_1112 | stp1 | protein phosphatase 2C domain-containing protein | 8.98 ± 14.19 |
| SAUSA300_0789 | hypothetical | putative thioredoxin | 8.89 ± 18.33 |
| SAUSA300_0379 | ahpF | alkyl hydroperoxide reductase subunit F | 8.46 ± 4.49 |
| SAUSA300_0348 | tatA | twin arginine-targeting protein translocase | 8.36 ± 5.53 |
| SAUSA300_0469 | rnmV | hypothetical protein | 8.35 ± 0.35 |
| SAUSA300_1792 | hypothetical | hypothetical protein | 8.20 ± 4.58 |
| SAUSA300_2061 | atpH | F0F1 ATP synthase subunit delta | 7.98 ± 1.29 |
| SAUSA300_1092 | pyrP | uracil permease | 7.85 ± 2.60 |
| SAUSA300_0905 | hypothetical | hypothetical protein | 7.61 ± 3.76 |
| SAUSA300_0444 | gltC | LysR family regulatory protein | 7.59 ± 2.70 |
| SAUSA300_2646 | trmE | tRNA modification GTPase TrmE | 7.41 ± 8.81 |
| SAUSA300_2105 | mtlF | PTS system, mannitol specific IIBC component | 6.95 ± 0.84 |
| SAUSA300_2486 | clpL | putative ATP-dependent Clp proteinase | 6.73 ± 0.02 |
| SAUSA300_1887 | pcrB | geranylgeranylglyceryl phosphate synthase-like protein | 6.58 ± 3.46 |
| SAUSA300_1653 | hypothetical | metal-dependent hydrolase | 6.25 ± 8.63 |
| SAUSA300_2393 | opuCa | glycine betaine/carnitine/choline ABC transporter ATP-binding protein | 6.25 ± 7.87 |
| SAUSA300_1183 | hypothetical | 2-oxoglutarate ferredoxin oxidoreductase subunit beta | 6.19 ± 1.88 |
| SAUSA300_0393 | hypothetical | hypothetical protein | 6.18 ± 2.30 |
| SAUSA300_0174 | hypothetical | hypothetical protein | 6.15 ± 1.39 |
| SAUSA300_0841 | hypothetical | hypothetical protein | 5.97 ± 2.99 |
| SAUSA300_1096 | carB | carbamoyl phosphate synthase large subunit | 5.89 ± 2.89 |
| SAUSA300_2593 | hypothetical | hypothetical protein | 5.84 ± 3.04 |
| SAUSA300_0221 | pflA | pyruvate formate-lyase activating enzyme | 5.68 ± 18.96 |
| SAUSA300_0996 | lpdA | dihydrolipoamide dehydrogenase | 5.49 ± 2.87 |
| SAUSA300_1992 | agrA | accessory gene regulator protein A | 5.34 ± 14.81 |
| SAUSA300_1147 | hslU | ATP-dependent protease ATP-binding subunit HslU | 4.99 ± 6.72 |
| SAUSA300_1120 | recG | ATP-dependent DNA helicase RecG | 4.60 ± 0.15 |
| SAUSA300_2078 | murA | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | 3.18 ± 3.15 |
| SAUSA300_1583 | cymR | hypothetical protein | 2.48 ± 0.46 |
| SAUSA300_0992 | hypothetical | hypothetical protein | 2.24 ± 20.30 |
| SAUSA300_0634 | fhuB | ferrichrome transport permease fhuB | 2.22 ± 4.57 |
| SAUSA300_0750 | whiA | hypothetical protein | 1.88 ± 4.32 |
| SAUSA300_2485 | hypothetical | methylated DNA-protein cysteine methyltransferase | 1.78 ± 9.18 |
| SAUSA300_0426 | hypothetical | hypothetical protein | 0.95 ± 5.09 |
| SAUSA300_2598 | capA | capsular polysaccharide biosynthesis protein Cap1A | 0.85 ± 1.11 |
| SAUSA300_2246 | hypothetical | hypothetical protein | 0.51 ± 16.59 |
| SAUSA300_2518 | hypothetical | hydrolase family protein | 0.42 ± 7.59 |
| SAUSA300_0355 | hypothetical | acetyl-CoA acetyltransferase | −0.68 a ± 4.44 |
| SAUSA300_0398 | hypothetical | superantigen-like protein | −0.83 a ± 2.42 |
| SAUSA300_2226 | moaB | molybdenum cofactor biosynthesis protein B | −1.15 a ± 5.09 |
| SAUSA300_0945 | hypothetical | isochorismate synthase family protein | −1.17 a ± 14.05 |
| SAUSA300_0904 | yjbI | hypothetical protein | −1.32 a ± 9.61 |
| SAUSA300_0423 | hypothetical | hypothetical protein | −2.20 a ± 9.08 |
| SAUSA300_1422 | hypothetical | phiSLT ORF65-like protein | −2.77 a ± 6.35 |
| SAUSA300_0068 | hypothetical | cadmium-exporting ATPase, truncation | −2.79 a ± 8.85 |
| SAUSA300_1870 | hypothetical | hypothetical protein | −2.92 a ± 15.58 |
| SAUSA300_1139 | sucD | succinyl-CoA synthetase subunit alpha | −2.94 a ± 8.32 |
| SAUSA300_0918 | ugtP | diacylglycerol glucosyltransferase | −3.09 a ± 8.63 |
| SAUSA300_0597 | hypothetical | putative endonuclease III | −3.15 a ± 14.78 |
| SAUSA300_0326 | hypothetical | hypothetical protein | −3.64 a ± 2.40 |
| SAUSA300_0690 | saeS | sensor histidine kinase SaeS | −4.88 a ± 14.01 |
| SAUSA300_0560 | vraB | acetyl-CoA c-acetyltransferase | −5.06 a ± 6.53 |
| SAUSA300_2334 | hypothetical | hypothetical protein | -5.12 a ± 7.55 |
| SAUSA300_2025 | rsbU | sigma-B regulation protein | −5.19 a ± 6.08 |
| SAUSA300_2152 | lacD | tagatose 1,6-diphosphate aldolase | −5.59 a ± 11.59 |
| SAUSA300_1680 | acuA | acetoin utilization protein AcuA | −5.94 a ± 10.87 |
| SAUSA300_2024 | rsbV | anti-sigma-B factor, antagonist | −6.77 a ± 14.71 |
| SAUSA300_0618 | mntC | ABC transporter substrate-binding protein | −6.85 a ± 4.61 |
| SAUSA300_1876 | hypothetical | DNA polymerase IV | −6.91 a ± 9.59 |
| SAUSA300_1465 | hypothetical | 2-oxoisovalerate dehydrogenase, E1 component, beta subunit | −7.15 a ± 6.73 |
| SAUSA300_1573 | hypothetical | Holliday junction resolvase-like protein | −10.10 a ± 6.88 |
| SAUSA300_1473 | nusB | transcription antitermination protein NusB | −10.84 a ± 10.00 |
| SAUSA300_1357 | aroC | chorismate synthase | −11.88 a ± 0.89 |
| SAUSA300_1095 | carA | carbamoyl phosphate synthase small subunit | −14.12 a ± 10.52 |
| SAUSA300_1469 | argR | arginine repressor | −14.16 a ± 8.61 |
| SAUSA300_1615 | hemB | delta-aminolevulinic acid dehydratase | −14.95 a ± 14.12 |
| SAUSA300_1467 | lpdA | dihydrolipoamide dehydrogenase | −15.68 a ± 14.07 |
| SAUSA300_0993 | pdhA | pyruvate dehydrogenase E1 component, alpha subunit | −17.05 a ± 10.66 |
| SAUSA300_0752 | clpP | ATP-dependent Clp protease proteolytic subunit | −17.66 a ± 11.34 |
| SAUSA300_1715 | ribD | riboflavin biosynthesis protein | −23.78 a ± 4.28 |
| Locus | Gene Name | Description | % EC Damage (Mean ± SD) |
|---|---|---|---|
| SAUSA300_1197 | ND a | glutathione peroxidase | 62.86 ± 5.67 |
| SAUSA300_1333 | hypothetical | conserved hypothetical protein | 62.17 ± 3.05 |
| SAUSA300_1485 | hypothetical | conserved hypothetical protein | 61.86 ± 6.12 |
| SAUSA300_2221 | moaD | molybdopterin converting factor, subunit 1 | 61.64 ± 3.61 |
| SAUSA300_0206 | azoR | flavodoxin family protein | 60.82 ± 6.24 |
| SAUSA300_0335 | mepA | MATE efflux family protein | 60.15 ± 8.13 |
| Locus | Group | Gene Name | % EC Damage (Mean ± SD) | |
|---|---|---|---|---|
| 384-Well Plates | 24-Well Plates | |||
| JE2 | Wildtype | 46.19 ± 2.97 | 42.43 ± 6.44 | |
| SAUSA300_1197 | EC damage ≥ 60% in 384-well plates | hypothetical | 62.86 ± 5.67 | 59.40 ± 1.50 |
| SAUSA300_1333 | hypothetical | 62.17 ± 3.05 | 66.92 ± 0.84 | |
| SAUSA300_1485 | hypothetical | 61.86 ± 6.12 | 61.75 a | |
| SAUSA300_2221 | moaD | 61.64 ± 3.61 | 59.90 ± 1.08 | |
| SAUSA300_0206 | hypothetical | 60.82 ± 6.24 | 69.33 ± 0.48 | |
| SAUSA300_0335 | hypothetical | 60.15 ± 8.31 | 63.35 ± 2.06 | |
| SAUSA300_1040 | EC damage ≤ 30% in 384-well plates | hypothetical | 26.74 ± 8.21 | 30.92 a |
| SAUSA300_1875 | hypothetical | 24.52 ± 10.68 | 30.51 a | |
| SAUSA300_0871 | hypothetical | 24.49 ± 12.19 | 28.60 a | |
| SAUSA300_1950 | hypothetical | 23.24 ± 9.64 | 25.87 a | |
| SAUSA300_0253 | scdA | 21.83 ± 12.24 | 22.52 a | |
| SAUSA300_0649 | hypothetical | 20.24 ± 0.89 | 22.65 a | |
| SAUSA300_2587 | hypothetical | 20.06 ± 9.42 | 26.45 a | |
| SAUSA300_0631 | hypothetical | 17.25 ± 11.20 | 23.00 a | |
| SAUSA300_2027 | alr | 16.70 ± 16.05 | 3.28 ± 1.38 | |
| SAUSA300_2055 | murA | 15.79 ± 10.49 | 7.62 ± 0.59 | |
| SAUSA300_1682 | ccpA | 14.04 ± 8.43 | 13.43 a | |
| SAUSA300_1696 | dat | 12.74 ± 5.48 | 14.99 ± 1.34 | |
| SAUSA300_0974 | purN | 11.07 ± 8.08 | 20.58 a | |
| SAUSA300_1563 | accC | 16.73 ± 11.04 | 11.82 ± 0.72 | |
| SAUSA300_0041 | hypothetical | 10.41 ± 2.09 | 3.30 a | |
| SAUSA300_0994 | pdhB | 11.20 ± 8.12 | 19.36 a | |
| SAUSA300_0186 | argC | 12.92 ± 16.00 | 15.20 ± 2.13 | |
| SAUSA300_1992 | agrA | 5.34 ± 14.81 | −3.82 ± 1.77 | |
| SAUSA300_0355 | hypothetical | −0.68 ± 4.44 | −1.20 a | |
| SAUSA300_0690 | saeS | −4.89 ± 14.01 | −12.80 ± 1.77 | |
| Categories | Sub-Groups | No. of Mutants with Decreased HMEC-1 Damage | No. of Mutants with Increased HMEC-1 Damage |
|---|---|---|---|
| Metabolism | Carbohydrate metabolism | 53 | |
| Amino acid metabolism | 33 | ||
| Metabolism of cofactors and vitamins | 11 | ||
| Lipid metabolism | 8 | 1 | |
| Nucleotide metabolism | 8 | ||
| Biosynthesis of other secondary metabolites | 7 | ||
| Energy metabolism | 7 | ||
| Metabolism of other amino acids | 3 | 1 | |
| Metabolism of terpenoids and polyketides | 3 | ||
| Glycan biosynthesis and metabolism | 2 | ||
| Xenobiotics biodegradation and metabolism | 1 | ||
| Genetic information processing | Homologous recombination | 4 | |
| DNA replication | 2 | ||
| Mismatch repair | 2 | ||
| Protein export | 2 | ||
| Ribosome | 2 | ||
| Sulfur relay system | 2 | 1 | |
| RNA degradation | 1 | ||
| Environmental information processing | Two-component system | 13 | |
| ABC transporters | 9 | ||
| Other | 3 | ||
| Cellular processes | Quorum sensing | 9 | |
| Total | 185 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Li, Y.; Li, L.; Xiong, Y.Q. Identification of Methicillin-Resistant Staphylococcus aureus (MRSA) Genetic Factors Involved in Human Endothelial Cells Damage, an Important Phenotype Correlated with Persistent Endovascular Infection. Antibiotics 2022, 11, 316. https://doi.org/10.3390/antibiotics11030316
Xiao X, Li Y, Li L, Xiong YQ. Identification of Methicillin-Resistant Staphylococcus aureus (MRSA) Genetic Factors Involved in Human Endothelial Cells Damage, an Important Phenotype Correlated with Persistent Endovascular Infection. Antibiotics. 2022; 11(3):316. https://doi.org/10.3390/antibiotics11030316
Chicago/Turabian StyleXiao, Xia, Yi Li, Liang Li, and Yan Q. Xiong. 2022. "Identification of Methicillin-Resistant Staphylococcus aureus (MRSA) Genetic Factors Involved in Human Endothelial Cells Damage, an Important Phenotype Correlated with Persistent Endovascular Infection" Antibiotics 11, no. 3: 316. https://doi.org/10.3390/antibiotics11030316
APA StyleXiao, X., Li, Y., Li, L., & Xiong, Y. Q. (2022). Identification of Methicillin-Resistant Staphylococcus aureus (MRSA) Genetic Factors Involved in Human Endothelial Cells Damage, an Important Phenotype Correlated with Persistent Endovascular Infection. Antibiotics, 11(3), 316. https://doi.org/10.3390/antibiotics11030316