
Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using Octyl Gallate, a Food-Grade Antioxidant



Abstract
1. Introduction
2. Results
2.1. Synergy between OG and Antibiotics against MRSA and Methicillin-Sensitive Staphylococcus aureus (MSSA)
2.2. Antimicrobial Synergy of OG with β-Lactams in Clinical Isolates of MRSA
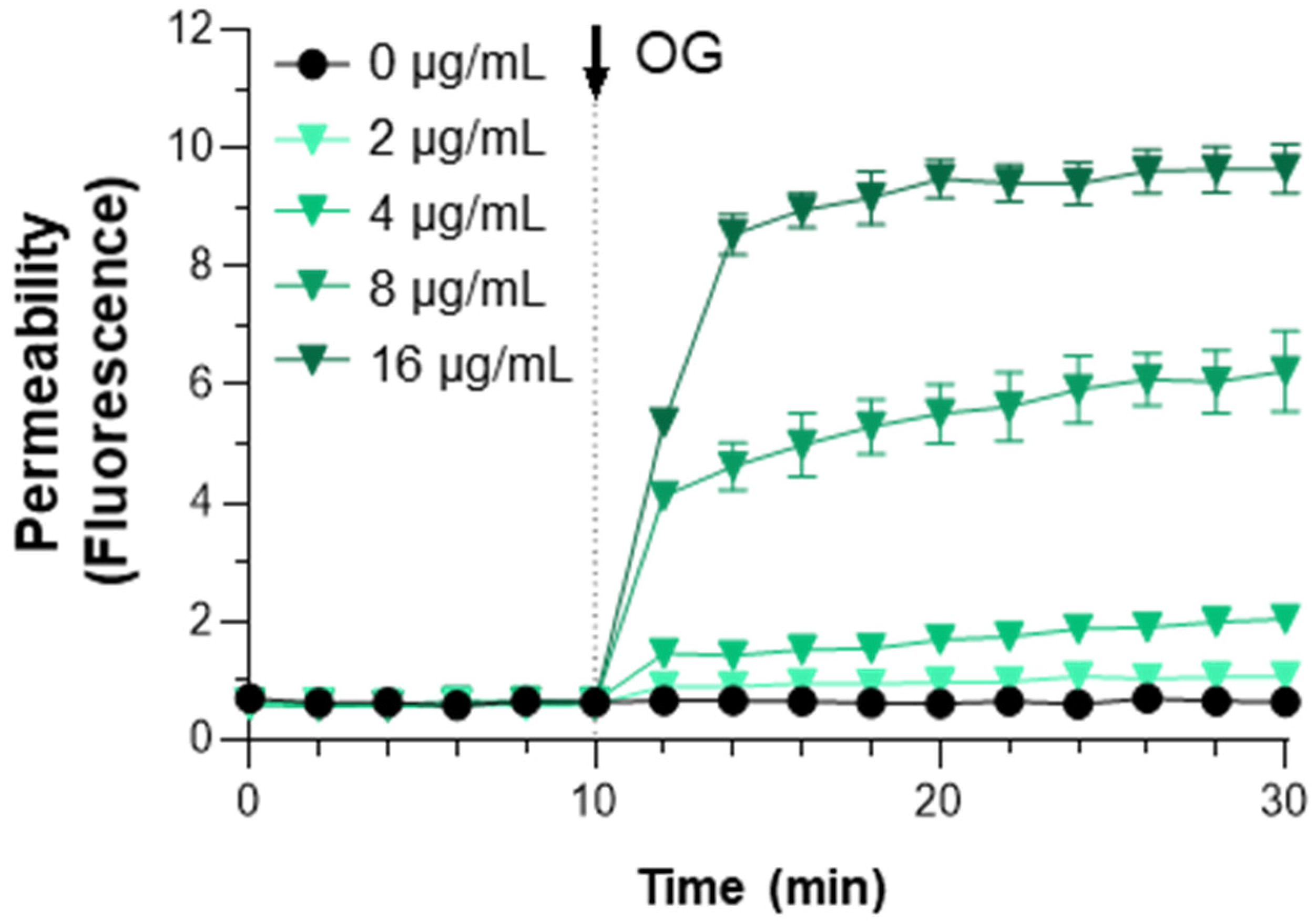
2.3. OG Increased Cell Wall Permeability in MRSA
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture
4.2. Antimicrobial Susceptibility Testing
4.3. Checkerboard Titration Assay
4.4. Calculation of FIC and FBC Indices
4.5. Cell Wall Permeability Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nathan, C.; Cars, O. Antibiotic Resistance—Problems, Progress, and Prospects. N. Engl. J. Med. 2014, 371, 1761–1763. [Google Scholar] [CrossRef]
- Tornimbene, B.; Eremin, S.; Escher, M.; Griskeviciene, J.; Manglani, S.; Pessoa-Silva, C.L. WHO Global Antimicrobial Resistance Surveillance System early implementation 2016–2017. Lancet Infect. Dis. 2018, 18, 241–242. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, S.; Mizutani, T.; Hiramatsu, K.; Kikuchi, K. Rapid Acquisition of linezolid resistance in methicillin-resistant Staphylococcus aureus: Role of hypermutation and homologous recombination. PLoS ONE 2016, 11, e0155512. [Google Scholar] [CrossRef]
- Carrel, M.; Perencevich, E.N.; David, M.Z. USA300 Methicillin-resistant Staphylococcus aureus, United States, 2000–2013. Emerg. Infect. Dis. 2015, 21, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Ghahremani, M.; Jazani, N.H.; Sharifi, Y. Emergence of vancomycin-intermediate and -resistant Staphylococcus aureus among methicillin-resistant S. aureus isolated from clinical specimens in the northwest of Iran. J. Glob. Antimicrob. Resist. 2018, 14, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, P.C. Reduced glycopeptide susceptibility in methicillin-resistant Staphylococcus aureus (MRSA). Int. J. Antimicrob. Agents 2007, 30, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Epstein, L.; Mu, Y.; Belflower, R.; Scott, J.; Ray, S.; Dumyati, G.; Felsen, C.; Petit, S.; Yousey-Hindes, K.; Nadle, J.; et al. Risk factors for invasive methicillin-resistant Staphylococcus aureus infection after recent discharge from an acute-care hospitalization, 2011–2013. Clin. Infect. Dis. 2016, 62, 45–52. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis 2009, 48, 1–12. [Google Scholar] [CrossRef]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2018, 53, 371–382. [Google Scholar] [CrossRef]
- Wright, G.D. Antibiotic adjuvants: Rescuing antibiotics from resistance. Trends Microbiol. 2016, 24, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Uto, L.R.; Gerriets, V. Clavulanic acid. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK545273/ (accessed on 18 January 2022).
- Kim, J.-C.; Jeon, B. Novel adjuvant strategy to potentiate bacitracin against multidrug-resistant methicillin-resistant Staphylococcus aureus (MRSA). J. Antimicrob. Chemother. 2016, 71, 1260–1263. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oh, E.; Bae, J.; Kumar, A.; Choi, H.J.; Jeon, B. Antioxidant-based synergistic eradication of methicillin-resistant Staphylococcus aureus (MRSA) biofilms with bacitracin. Int. J. Antimicrob. Agents 2018, 52, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Wu, S.C.; Han, F.; Song, M.R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.Z. Natural flavones from Morus alba against methicillin-resistant Staphylococcus aureus via targeting the proton motive force and membrane permeability. J. Agric. Food Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiang, Y.; Xu, M. From red to green: The propidium iodide-permeable membrane of Shewanella decolorationis S12 is repairable. Sci. Rep. 2015, 5, 18583. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Resisting resistance: New chemical strategies for battling superbugs. Chem. Biol. 2000, 7, R127–R132. [Google Scholar] [CrossRef]
- Pages, J.M.; Amaral, L. Mechanisms of drug efflux and strategies to combat them: Challenging the efflux pump of Gram-negative bacteria. Biochim. Biophys. Acta 2009, 1794, 826–833. [Google Scholar] [CrossRef]
- Kim, W.; Zou, G.; Hari, T.P.A.; Wilt, I.K.; Zhu, W.; Galle, N.; Faizi, H.A.; Hendricks, G.L.; Tori, K.; Pan, W.; et al. A selective membrane-targeting repurposed antibiotic with activity against persistent methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2019, 116, 16529–16534. [Google Scholar] [CrossRef]
- Yarlagadda, V.; Medina, R.; Wright, G.D. Venturicidin A, a Membrane-active natural product inhibitor of ATP synthase potentiates aminoglycoside antibiotics. Sci. Rep. 2020, 10, 8134. [Google Scholar] [CrossRef]
- FDA. Food Additive Status List. Available online: http://www.fda.gov/Food/IngredientsPackagingLabeling/FoodAdditivesIngredients/ucm091048.htm (accessed on 18 January 2022).
- Shi, Y.-G.; Li, D.-H.; Kong, Y.-M.; Zhang, R.-R.; Gu, Q.; Hu, M.-X.; Tian, S.-Y.; Jin, W.-G. Enhanced antibacterial efficacy and mechanism of octyl gallate/beta-cyclodextrins against Pseudomonas fluorescens and Vibrio parahaemolyticus and incorporated electrospun nanofibers for Chinese giant salamander fillets preservation. Int. J. Food Microbiol. 2022, 361, 109460. [Google Scholar] [CrossRef]
- Stapleton, P.D.; Shah, S.; Anderson, J.C.; Hara, Y.; Hamilton-Miller, J.M.; Taylor, P.W. Modulation of beta-lactam resistance in Staphylococcus aureus by catechins and gallates. Int. J. Antimicrob. Agents 2004, 23, 462–467. [Google Scholar] [CrossRef]
- Stone, K.J.; Strominger, J.L. Mechanism of action of bacitracin: Complexation with metal ion and C 55-isoprenyl pyrophosphate. Proc. Natl. Acad. Sci. USA 1971, 68, 3223–3227. [Google Scholar] [CrossRef] [PubMed]
- M07-A10. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 10th ed.; 267 Approved Standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Hsieh, M.H.; Chen, M.Y.; Victor, L.Y.; Chow, J.W. Synergy assessed by checkerboard a critical analysis. Diagn. Microbiol. Infect. Dis. 1993, 16, 343–349. [Google Scholar] [CrossRef]
- Rand, K.H.; Houck, H.J.; Brown, P.; Bennett, D. Reproducibility of the microdilution checkerboard method for antibiotic synergy. Antimicrob. Agents Chemother. 1993, 37, 613–615. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antibiotic | MIC (µg/mL) ⱡ | MBC (µg/mL) ⱡ | ||||
|---|---|---|---|---|---|---|
| Antibiotic Alone | Antibiotic Plus OG | FIC Index | Antibiotic Alone | Antibiotic Plus OG | FBC Index | |
| MSSA | ||||||
| Penicillin | 1 | 0.25 (4) | 0.094 | 1 | 0.25 (4) | 0.281 |
| Ampicillin | 2 | 0.125 (16) | 0.125 | 2 | 0.125 (16) | 0.094 |
| Cephalothin | 0.5 | 0.125 (4) | 0.5 | 0.5 | 0.125 (4) | 0.258 |
| Gentamicin | 1 | 0.125 (8) | 0.129 | 1 | 0.125 (8) | 0.144 |
| Chloramphenicol | 16 | 2 (8) | 0.329 | 32 | 2 (16) | 0.082 |
| Tetracycline | 1 | 0.125 (8) | 0.129 | 2 | 0.063 (32) | 0.051 |
| Erythromycin | 0.5 | 0.25 (2) | 0.539 | 2 | 0.25 (8) | 0.144 |
| Lincomycin | 1 | 0.5 (2) | 0.539 | 2 | 0.25 (8) | 0.144 |
| MRSA | ||||||
| Penicillin | >64 | 0.25 (>256) | 0.127 | >64 | 8 (>8) | 0.126 |
| Ampicillin | >64 | 0.5 (>128) | 0.129 | >64 | 1 (>64) | 0.066 |
| Cephalothin | >64 | 1 (>64) | 0.133 | 64 | 2 (32) | 0.070 |
| Gentamicin | >64 | 8 (>8) | 0.156 | >64 | 16 (>4) | 0.047 |
| Chloramphenicol | 16 | 1 (16) | 0.188 | 32 | 2 (16) | 0.125 |
| Tetracycline | >64 | 16 (>4) | 0.188 | >64 | 64 (>1) | 0.375 |
| Erythromycin | >64 | >4 (>16) | >2 | >64 | >4 (>16) | >2 |
| Lincomycin | >64 | >16 (>4) | >2 | >64 | >16 (>4) | >2 |
| Strain | MIC (μg/mL) | MIC of Combinations (μg/mL) ⱡ | FIC Index | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PEN | AMP | CEP | PEN + OG | AMP + OG | CEP + OG | PEN + OG | AMP + OG | CEP + OG | |
| MRSA 3795 | >64 | >64 | >64 | 0.063 (>1024) | 0.125 (>512) | 1 (>64) | 0.032 | 0.032 | 0.039 |
| MRSA 3823 | >64 | >64 | >64 | 0.063 (>1024) | 0.125 (>512) | 1 (>64) | 0.126 | 0.126 | 0.133 |
| MRSA 3865 | 64 | >64 | >64 | 0.5 (128) | 1 (>64) | 4 (>16) | 0.070 | 0.070 | 0.156 |
| MRSA 3903 | >64 | >64 | >64 | 0.125 (>512) | 0.125 (>512) | 1 (>64) | 0.126 | 0.126 | 0.133 |
| Strain | MBC (μg/mL) | MBC of Combinations (μg/mL) ⱡ | FBC Index | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PEN | AMP | CEP | PEN + OG | AMP + OG | CEP + OG | PEN + OG | AMP + OG | CEP + OG | |
| MRSA 3795 | >64 | >64 | >64 | 0.063 (>1024) | 0.25 (>256) | 1 (>64) | 0.063 | 0.063 | 0.035 |
| MRSA 3823 | >64 | >64 | >64 | 0.5 (>128) | 0.5 (>128) | 2 (>32) | 0.033 | 0.127 | 0.039 |
| MRSA 3865 | 64 | >64 | >64 | 4 (16) | 8 (>8) | 8 (>8) | 0.188 | 0.156 | 0.156 |
| MRSA 3903 | >64 | >64 | >64 | 0.5 (>128) | 0.5 (>128) | 2 (>32) | 0.033 | 0.033 | 0.039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamang, M.D.; Bae, J.; Park, M.; Jeon, B. Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using Octyl Gallate, a Food-Grade Antioxidant. Antibiotics 2022, 11, 266. https://doi.org/10.3390/antibiotics11020266
Tamang MD, Bae J, Park M, Jeon B. Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using Octyl Gallate, a Food-Grade Antioxidant. Antibiotics. 2022; 11(2):266. https://doi.org/10.3390/antibiotics11020266
Chicago/Turabian StyleTamang, Migma Dorji, Junghee Bae, Myungseo Park, and Byeonghwa Jeon. 2022. "Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using Octyl Gallate, a Food-Grade Antioxidant" Antibiotics 11, no. 2: 266. https://doi.org/10.3390/antibiotics11020266
APA StyleTamang, M. D., Bae, J., Park, M., & Jeon, B. (2022). Potentiation of β-Lactams against Methicillin-Resistant Staphylococcus aureus (MRSA) Using Octyl Gallate, a Food-Grade Antioxidant. Antibiotics, 11(2), 266. https://doi.org/10.3390/antibiotics11020266