
Genetic Mechanisms of Vancomycin Resistance in Clostridioides difficile: A Systematic Review


Abstract
1. Introduction
2. Results
2.1. Binding Site Alterations
2.1.1. Van Genes

{kind=link}
| Ref. | Gene | Genetic Mechanism (If Described) | Encoded Protein | Function | Proposed Mechanism of Resistance |
|---|---|---|---|---|---|
| [29] | agrD1 | Overexpression | Accessory gene regulator D1 (AgrD1) | Quorum sensing generation/modulation | Unknown |
| [27] | cd2068 | Upregulation | CD2068 | ABC transporter pump | Drug efflux* |
| [25] | cd3659 | Mutation (Gly982Thr) | DNA exonuclease/phosphodiesterase | -- | Unknown |
| [28] | cdtA | Presence ** | Toxin A (TcdA) | Enterotoxin | No impact |
| [28] | cdtB | Presence ** | Toxin B (TcdB) | Cytotoxin | No impact |
| [28] | cwlD | Presence ** | N-acetylmuramoyl-L-alanine amidase | Spore germination | No impact |
| [24,28,29] | cwp84 | Overexpression | Cysteine protease (Cwp84) | Surface layer protein maturation/adhesion/biofilm regulation | Biofilm production |
| [28] | fliC | Presence ** | Flagellin (FliC) | Adhesion/biofilm regulation | Biofilm production |
| [28,29] | luxS | Presence ** | LuxS | Autoinducer-2 synthesis/quorum sensing modulation/biofilm regulation | Biofilm production |
| [26] | pX18-498_006 *** | Presence | N-acetylmuramoyl-L-alanine amidase | Cell wall integrity | Unknown |
| [25] | sdaB/cd3222 | Mutation (Ala295 deletion) | L-serine deaminase | -- | Unknown |
| [29] | sigH | Overexpression | Sigma factor (SigH) | Sporulation regulation | Biofilm production |
| [28] | sleC | Presence ** | SleC | Spore germination | Biofilm production |
| [29] | slpA | Presence | S-layer protein (SlpA) | Surface layer composition/ adhesion/biofilm regulation | No impact |
| [28,29] | spo0A | Overexpression | Spo0A | Sporulation/biofilm regulation | Biofilm production |
2.1.2. MurG
2.2. Plasmid Acquisition of N-Acetylmuramoyl-L-Alanine
2.3. Efflux Pumps
2.4. RNA Polymerase Mutations
2.5. Biofilm Production
2.5.1. Surface Factors Involved in Biofilm Formation
2.5.2. Sporulation and Germination
2.5.3. Quorum Sensing
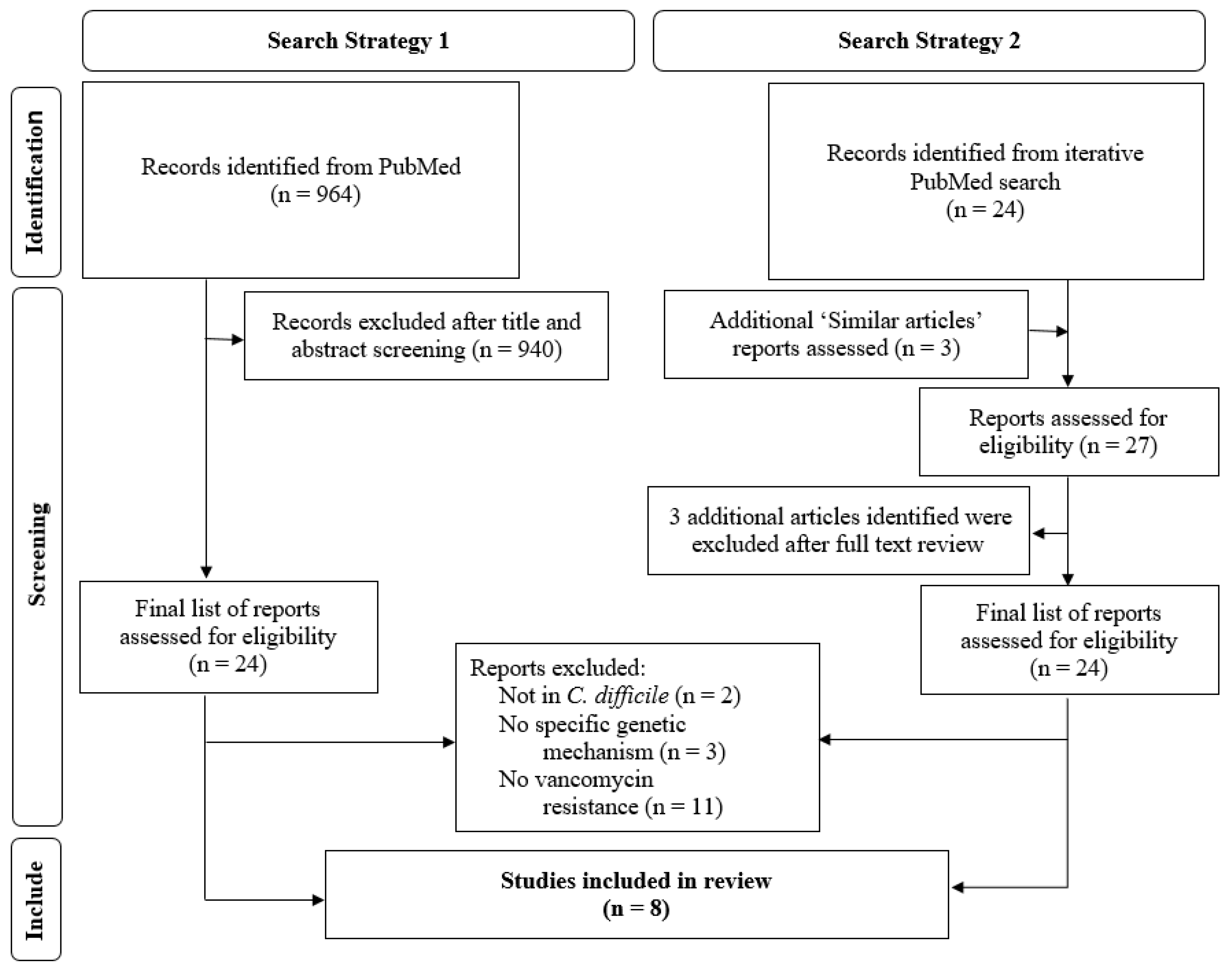
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Magill, S.S.; O’Leary, E.; Janelle, S.J.; Thompson, D.L.; Dumyati, G.; Nadle, J.; Wilson, L.E.; Kainer, M.A.; Lynfield, R.; Greissman, S.; et al. Changes in Prevalence of Health Care-Associated Infections in U.S. Hospitals. N. Engl. J. Med. 2018, 379, 1732–1744. [Google Scholar] [CrossRef] [PubMed]
- Antibiotic Resistance Threats in the United States. 2019. Available online: https;//www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 13 January 2022).
- Gonzales, M.; Pepin, J.; Frost, E.H.; Carrier, J.C.; Sirard, S.; Fortier, L.C.; Valiquette, L. Faecal pharmacokinetics of orally administered vancomycin in patients with suspected Clostridium difficile infection. BMC Infect. Dis. 2010, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 5 January 2022).
- CLSI M100-ED31: 2021 Performance Standards for Antimicrobial Susceptibility Testing, 31st Edition. Available online: http://em100.edaptivedocs.net/GetDoc.aspx?doc=CLSI%20M100%20ED31:2021&scope=user (accessed on 13 January 2022).
- Gonzales-Luna, A.J.; Olaitan, A.O.; Shen, W.J.; Deshpande, A.; Carlson, T.J.; Dotson, K.M.; Lancaster, C.; Begum, K.; Alam, M.J.; Hurdle, J.G.; et al. Reduced Susceptibility to Metronidazole Is Associated With Initial Clinical Failure in Clostridioides difficile Infection. Open Forum Infect. Dis. 2021, 8, ofab365. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Louie, T.J.; Gerding, D.N.; Cornely, O.A.; Chasan-Taber, S.; Fitts, D.; Gelone, S.P.; Broom, C.; Davidson, D.M. Vancomycin, Metronidazole, or Tolevamer for Clostridium difficile Infection: Results From Two Multinational, Randomized, Controlled Trials. Clin. Infect. Dis. 2014, 59, 345–354. [Google Scholar] [CrossRef]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, 987–994. [Google Scholar] [CrossRef]
- Clancy, C.J.; Buehrle, D.; Vu, M.; Wagener, M.M.; Nguyen, M.H. Impact of Revised Infectious Diseases Society of America and Society for Healthcare Epidemiology of America Clinical Practice Guidelines on the Treatment of Clostridium difficile Infections in the United States. Clin. Infect. Dis. 2021, 72, 1944–1949. [Google Scholar] [CrossRef]
- Louie, T.J.; Miller, M.A.; Mullane, K.M.; Weiss, K.; Lentnek, A.; Golan, Y.; Gorbach, S.; Sears, P.; Shue, Y.K.; Group, O.P.T.C.S. Fidaxomicin versus vancomycin for Clostridium difficile infection. N. Engl. J. Med. 2011, 364, 422–431. [Google Scholar] [CrossRef]
- Cornely, O.A.; Crook, D.W.; Esposito, R.; Poirier, A.; Somero, M.S.; Weiss, K.; Sears, P.; Gorbach, S. Fidaxomicin versus vancomycin for infection with Clostridium difficile in Europe, Canada, and the USA: A double-blind, non-inferiority, randomised controlled trial. Lancet Infect. Dis. 2012, 12, 281–289. [Google Scholar] [CrossRef]
- Mikamo, H.; Tateda, K.; Yanagihara, K.; Kusachi, S.; Takesue, Y.; Miki, T.; Oizumi, Y.; Gamo, K.; Hashimoto, A.; Toyoshima, J.; et al. Efficacy and safety of fidaxomicin for the treatment of Clostridioides (Clostridium) difficile infection in a randomized, double-blind, comparative Phase III study in Japan. J. Infect. Chemother. 2018, 24, 744–752. [Google Scholar] [CrossRef]
- Guery, B.; Menichetti, F.; Anttila, V.J.; Adomakoh, N.; Aguado, J.M.; Bisnauthsing, K.; Georgopali, A.; Goldenberg, S.D.; Karas, A.; Kazeem, G.; et al. Extended-pulsed fidaxomicin versus vancomycin for Clostridium difficile infection in patients 60 years and older (EXTEND): A randomised, controlled, open-label, phase 3b/4 trial. Lancet Infect. Dis. 2018, 18, 296–307. [Google Scholar] [CrossRef]
- Johnson, S.; Lavergne, V.; Skinner, A.M.; Gonzales-Luna, A.J.; Garey, K.W.; Kelly, C.P.; Wilcox, M.H. Clinical Practice Guideline by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA): 2021 Focused Update Guidelines on Management of Clostridioides difficile Infection in Adults. Clin. Infect. Dis. 2021, 73, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Wenisch, C.; Parschalk, B.; Hasenhundl, M.; Hirschl, A.M.; Graninger, W. Comparison of vancomycin, teicoplanin, metronidazole, and fusidic acid for the treatment of Clostridium difficile-associated diarrhea. Clin. Infect. Dis. 1996, 22, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Zar, F.A.; Bakkanagari, S.R.; Moorthi, K.M.L.S.T.; Davis, M.B. A Comparison of Vancomycin and Metronidazole for the Treatment of Clostridium difficile-Associated Diarrhea, Stratified by Disease Severity. Clin. Infect. Dis. 2007, 45, 302–307. [Google Scholar] [CrossRef]
- Louie, T.J.; Peppe, J.; Watt, C.K.; Johnson, D.; Mohammed, R.; Dow, G.; Weiss, K.; Simon, S.; John, J.F., Jr.; Garber, G.; et al. Tolevamer, a novel nonantibiotic polymer, compared with vancomycin in the treatment of mild to moderately severe Clostridium difficile-associated diarrhea. Clin. Infect. Dis. 2006, 43, 411–420. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dudley, M.N.; McLaughlin, J.C.; Carrington, G.; Frick, J.; Nightingale, C.H.; Quintiliani, R. Oral bacitracin vs vancomycin therapy for Clostridium difficile-induced diarrhea. A randomized double-blind trial. Arch. Intern. Med. 1986, 146, 1101–1104. [Google Scholar] [CrossRef]
- Jon, J.V.; Mark, H.W.; Jane, F. Antimicrobial resistance progression in the United Kingdom: A temporal comparison of Clostridioides difficile antimicrobial susceptibilities. Anaerobe 2021, 70, 102385. [Google Scholar] [CrossRef]
- Adler, A.; Miller-Roll, T.; Bradenstein, R.; Block, C.; Mendelson, B.; Parizade, M.; Paitan, Y.; Schwartz, D.; Peled, N.; Carmeli, Y.; et al. A national survey of the molecular epidemiology of Clostridium difficile in Israel: The dissemination of the ribotype 027 strain with reduced susceptibility to vancomycin and metronidazole. Diagn. Microbiol. Infect. Dis. 2015, 83, 21–24. [Google Scholar] [CrossRef]
- Jarrad, A.M.; Blaskovich, M.A.T.; Prasetyoputri, A.; Karoli, T.; Hansford, K.A.; Cooper, M.A. Detection and Investigation of Eagle Effect Resistance to Vancomycin in Clostridium difficile With an ATP-Bioluminescence Assay. Front. Microbiol. 2018, 9, 1420. [Google Scholar] [CrossRef]
- Ramirez-Vargas, G.; Quesada-Gomez, C.; Acuna-Amador, L.; Lopez-Urena, D.; Murillo, T.; Del Mar Gamboa-Coronado, M.; Chaves-Olarte, E.; Thomson, N.; Rodriguez-Cavallini, E.; Rodriguez, C. A Clostridium difficile Lineage Endemic to Costa Rican Hospitals Is Multidrug Resistant by Acquisition of Chromosomal Mutations and Novel Mobile Genetic Elements. Antimicrob. Agents Chemother. 2017, 61, e02054-16. [Google Scholar] [CrossRef]
- Shen, W.J.; Deshpande, A.; Hevener, K.E.; Endres, B.T.; Garey, K.W.; Palmer, K.L.; Hurdle, J.G. Constitutive expression of the cryptic vanGCd operon promotes vancomycin resistance in Clostridioides difficile clinical isolates. J. Antimicrob. Chemother. 2020, 75, 859–867. [Google Scholar] [CrossRef]
- Saldanha, G.Z.; Pires, R.N.; Rauber, A.P.; de Lima-Morales, D.; Falci, D.R.; Caierao, J.; Pasqualotto, A.C.; Martins, A.F. Genetic relatedness, Virulence factors and Antimicrobial Resistance of C. difficile strains from hospitalized patients in a multicentric study in Brazil. J. Glob. Antimicrob. Resist. 2020, 22, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Leeds, J.A.; Sachdeva, M.; Mullin, S.; Barnes, S.W.; Ruzin, A. In vitro selection, via serial passage, of Clostridium difficile mutants with reduced susceptibility to fidaxomicin or vancomycin. J. Antimicrob. Chemother. 2014, 69, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Pu, M.; Cho, J.M.; Cunningham, S.A.; Behera, G.K.; Becker, S.; Amjad, T.; Greenwood-Quaintance, K.E.; Mendes-Soares, H.; Jones-Hall, Y.; Jeraldo, P.R.; et al. Plasmid Acquisition Alters Vancomycin Susceptibility in Clostridioides difficile. Gastroenterology 2021, 160, 941–945.e8. [Google Scholar] [CrossRef]
- Ngernsombat, C.; Sreesai, S.; Harnvoravongchai, P.; Chankhamhaengdecha, S.; Janvilisri, T. CD2068 potentially mediates multidrug efflux in Clostridium difficile. Sci. Rep. 2017, 7, 9982. [Google Scholar] [CrossRef] [PubMed]
- Ðapa, T.; Leuzzi, R.; Ng, Y.K.; Baban, S.T.; Adamo, R.; Kuehne, S.A.; Scarselli, M.; Minton, N.P.; Serruto, D.; Unnikrishnan, M. Multiple factors modulate biofilm formation by the anaerobic pathogen Clostridium difficile. J. Bacteriol. 2013, 195, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Tijerina-Rodriguez, L.; Villarreal-Trevino, L.; Baines, S.D.; Morfin-Otero, R.; Camacho-Ortiz, A.; Flores-Trevino, S.; Maldonado-Garza, H.; Rodriguez-Noriega, E.; Garza-Gonzalez, E. High sporulation and overexpression of virulence factors in biofilms and reduced susceptibility to vancomycin and linezolid in recurrent Clostridium [Clostridioides] difficile infection isolates. PLoS ONE 2019, 14, e0220671. [Google Scholar] [CrossRef] [PubMed]
- Depardieu, F.; Mejean, V.; Courvalin, P. Competition between VanU(G) repressor and VanR(G) activator leads to rheostatic control of vanG vancomycin resistance operon expression. PLoS Genet. 2015, 11, e1005170. [Google Scholar] [CrossRef]
- O’Grady, K.; Knight, D.R.; Riley, T.V. Antimicrobial resistance in Clostridioides difficile. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2459–2478. [Google Scholar] [CrossRef]
- Woods, E.C.; Wetzel, D.; Mukerjee, M.; McBride, S.M. Examination of the Clostridioides (Clostridium) difficile VanZ ortholog, CD1240. Anaerobe 2018, 53, 108–115. [Google Scholar] [CrossRef]
- Knight, D.R.; Androga, G.O.; Ballard, S.A.; Howden, B.P.; Riley, T.V. A Phenotypically Silent vanB2 Operon Carried on a Tn1549-Like Element in Clostridium difficile. mSphere 2016, 1, e00177-16. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, C.; Li, W.G.; Xu, J.L.; Zhang, W.Z.; Dai, Y.F.; Lu, J.X. Independent Microevolution Mediated by Mobile Genetic Elements of Individual Clostridium difficile Isolates from Clade 4 Revealed by Whole-Genome Sequencing. mSystems 2019, 4, e00252-28. [Google Scholar] [CrossRef]
- Ammam, F.; Meziane-Cherif, D.; Mengin-Lecreulx, D.; Blanot, D.; Patin, D.; Boneca, I.G.; Courvalin, P.; Lambert, T.; Candela, T. The functional vanGCd cluster of Clostridium difficile does not confer vancomycin resistance. Mol. Microbiol. 2013, 89, 612–625. [Google Scholar] [CrossRef]
- Roxas, B.A.P.; Roxas, J.L.; Claus-Walker, R.; Harishankar, A.; Mansoor, A.; Anwar, F.; Jillella, S.; Williams, A.; Lindsey, J.; Elliott, S.P.; et al. Phylogenomic analysis of Clostridioides difficile ribotype 106 strains reveals novel genetic islands and emergent phenotypes. Sci. Rep. 2020, 10, 22135. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Liu, S.; Meng, X.; Liang, W.; Xu, Z.; Tang, B.; Wang, Y.; Duan, J.; Fu, C.; Wu, B.; et al. Genome characterization of a novel binary toxin-positive strain of Clostridium difficile and comparison with the epidemic 027 and 078 strains. Gut Pathog. 2017, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Ammam, F.; Marvaud, J.C.; Lambert, T. Distribution of the vanG-like gene cluster in Clostridium difficile clinical isolates. Can. J. Microbiol. 2012, 58, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, K.; Marvaud, J.C.; Lambert, T. Analysis of the mobilization functions of the vancomycin resistance transposon Tn1549, a member of a new family of conjugative elements. J. Bacteriol. 2010, 192, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.; Gross, B.; Walker, S. E. coli MurG: A paradigm for a superfamily of glycosyltransferases. Curr. Drug Targets Infect. Disord. 2001, 1, 201–213. [Google Scholar] [CrossRef]
- Liu, X. ABC Family Transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar] [CrossRef]
- Rafii, F.; Park, M. Detection and characterization of an ABC transporter in Clostridium hathewayi. Arch. Microbiol. 2008, 190, 417–426. [Google Scholar] [CrossRef]
- Rafii, F.; Park, M.; Carman, R.J. Characterization of an ATP-binding cassette from Clostridium perfringens with homology to an ABC transporter from Clostridium hathewayi. Anaerobe 2009, 15, 116–121. [Google Scholar] [CrossRef]
- McBride, S.M.; Sonenshein, A.L. The dlt operon confers resistance to cationic antimicrobial peptides in Clostridium difficile. Microbiology (Reading) 2011, 157, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Dannheim, H.; Riedel, T.; Neumann-Schaal, M.; Bunk, B.; Schober, I.; Sproer, C.; Chibani, C.M.; Gronow, S.; Liesegang, H.; Overmann, J.; et al. Manual curation and reannotation of the genomes of Clostridium difficile 630Deltaerm and C. difficile 630. J. Med. Microbiol. 2017, 66, 286–293. [Google Scholar] [CrossRef]
- Curry, S.R.; Marsh, J.W.; Shutt, K.A.; Muto, C.A.; O’Leary, M.M.; Saul, M.I.; Pasculle, A.W.; Harrison, L.H. High frequency of rifampin resistance identified in an epidemic Clostridium difficile clone from a large teaching hospital. Clin. Infect. Dis. 2009, 48, 425–429. [Google Scholar] [CrossRef]
- Babakhani, F.; Gomez, A.; Robert, N.; Sears, P. Killing kinetics of fidaxomicin and its major metabolite, OP-1118, against Clostridium difficile. J. Med. Microbiol. 2011, 60, 1213–1217. [Google Scholar] [CrossRef]
- Gonzales-Luna, A.J.; Spinler, J.K.; Oezguen, N.; Khan, M.A.W.; Danhof, H.A.; Endres, B.T.; Alam, M.J.; Begum, K.; Lancaster, C.; Costa, G.P.; et al. Systems biology evaluation of refractory Clostridioides difficile infection including multiple failures of fecal microbiota transplantation. Anaerobe 2021, 70, 102387. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Cui, L.; Katayama, Y.; Kozue, K.; Hiramatsu, K. Impact of rpoB mutations on reduced vancomycin susceptibility in Staphylococcus aureus. J. Clin. Microbiol. 2011, 49, 2680–2684. [Google Scholar] [CrossRef] [PubMed]
- Meza-Torres, J.; Auria, E.; Dupuy, B.; Tremblay, Y.D.N. Wolf in Sheep’s Clothing: Clostridioides difficile Biofilm as a Reservoir for Recurrent Infections. Microorganisms 2021, 9, 1922. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef]
- Wu, X.; Cherian, P.T.; Lee, R.E.; Hurdle, J.G. The membrane as a target for controlling hypervirulent Clostridium difficile infections. J. Antimicrob. Chemother. 2013, 68, 806–815. [Google Scholar] [CrossRef]
- Donelli, G.; Vuotto, C.; Cardines, R.; Mastrantonio, P. Biofilm-growing intestinal anaerobic bacteria. FEMS Immunol. Med. Microbiol. 2012, 65, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Semenyuk, E.G.; Laning, M.L.; Foley, J.; Johnston, P.F.; Knight, K.L.; Gerding, D.N.; Driks, A. Spore formation and toxin production in Clostridium difficile biofilms. PLoS ONE 2014, 9, e87757. [Google Scholar] [CrossRef] [PubMed]
- Janoir, C.; Pechine, S.; Grosdidier, C.; Collignon, A. Cwp84, a surface-associated protein of Clostridium difficile, is a cysteine protease with degrading activity on extracellular matrix proteins. J. Bacteriol. 2007, 189, 7174–7180. [Google Scholar] [CrossRef] [PubMed]
- Kirby, J.M.; Ahern, H.; Roberts, A.K.; Kumar, V.; Freeman, Z.; Acharya, K.R.; Shone, C.C. Cwp84, a surface-associated cysteine protease, plays a role in the maturation of the surface layer of Clostridium difficile. J. Biol. Chem. 2009, 284, 34666–34673. [Google Scholar] [CrossRef]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Lazazzera, B.A. The sporulation transcription factor Spo0A is required for biofilm development in Bacillus subtilis. Mol. Microbiol. 2001, 42, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-Sensing Regulation of Antimicrobial Resistance in Bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef]
- Slater, R.T.; Frost, L.R.; Jossi, S.E.; Millard, A.D.; Unnikrishnan, M. Clostridioides difficile LuxS mediates inter-bacterial interactions within biofilms. Sci. Rep. 2019, 9, 9903. [Google Scholar] [CrossRef]
- Hardie, K.R.; Heurlier, K. Establishing bacterial communities by ‘word of mouth’: LuxS and autoinducer 2 in biofilm development. Nat. Rev. Microbiol. 2008, 6, 635–643. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Quadri, L.E.; Kuipers, O.P.; de Vos, W.M. Quorum sensing by peptide pheromones and two-component signal-transduction systems in Gram-positive bacteria. Mol. Microbiol. 1997, 24, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Buehrle, D.; Clancy, C.J. Medicare Prescription Plans Limit Access to Recommended Drugs for Clostridioides difficile Infection. Clin. Infect. Dis. 2021, ciab898. [Google Scholar] [CrossRef] [PubMed]

| Ref. | No. Isolates † | Strain Origin | Ribotype | ST Type | Other | Gene (Mutation, If Known) | VAN MIC (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| vanG/vanSR | vanA | vanB | vanW | vanZ | murG | rpoC | |||||||
| [22] | 4 | Clinical | 012 | 54 | NAPCR1 | vanG | 4 | ||||||
| [23] * | 1 | Laboratory | 027 | 1 | WS2/ R20291 | vanS (Arg314Leu) | 8 | ||||||
| 1 | Laboratory | 027 | 1 | WS4/ R20291 | vanS (Gly319Asp) | 16 | |||||||
| 1 | Clinical | 027 | 1 | MT1470 | vanS (Ser313Phe) | 8 | |||||||
| 1 | Clinical | 027 | 1 | MT5006 | vanS (Thr349Ile) | 8 | |||||||
| 9 | Clinical | 027 | 1 | see note ‡ | vanR (Thr115Ala) | 4–8 ‡ | |||||||
| [24] | 1 | Clinical | 35 | vanA | vanW | 4 | |||||||
| 1 | Clinical | 014-020 | 2 | vanB | 8 | ||||||||
| 1 | Clinical | 42 | vanB | 4 | |||||||||
| 1 | Clinical | 67 | vanB | vanW | >16 | ||||||||
| 1 | Clinical | 012 | 54 | vanW | vanZ | 4 | |||||||
| [25] * | 1 | Laboratory | 087 | NB95013/ATCC 43255 | murG/cd2725 (Pro108Leu) | 16 | |||||||
| 1 | Laboratory | NB95026 | rpoC/cd0067 (Gly733Thr) | 16 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eubank, T.A.; Gonzales-Luna, A.J.; Hurdle, J.G.; Garey, K.W. Genetic Mechanisms of Vancomycin Resistance in Clostridioides difficile: A Systematic Review. Antibiotics 2022, 11, 258. https://doi.org/10.3390/antibiotics11020258
Eubank TA, Gonzales-Luna AJ, Hurdle JG, Garey KW. Genetic Mechanisms of Vancomycin Resistance in Clostridioides difficile: A Systematic Review. Antibiotics. 2022; 11(2):258. https://doi.org/10.3390/antibiotics11020258
Chicago/Turabian StyleEubank, Taryn A., Anne J. Gonzales-Luna, Julian G. Hurdle, and Kevin W. Garey. 2022. "Genetic Mechanisms of Vancomycin Resistance in Clostridioides difficile: A Systematic Review" Antibiotics 11, no. 2: 258. https://doi.org/10.3390/antibiotics11020258
APA StyleEubank, T. A., Gonzales-Luna, A. J., Hurdle, J. G., & Garey, K. W. (2022). Genetic Mechanisms of Vancomycin Resistance in Clostridioides difficile: A Systematic Review. Antibiotics, 11(2), 258. https://doi.org/10.3390/antibiotics11020258