
High Activity of N-Acetylcysteine in Combination with Beta-Lactams against Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii

 , , ,
, , ,  ,
,  , , and
, , and
Abstract
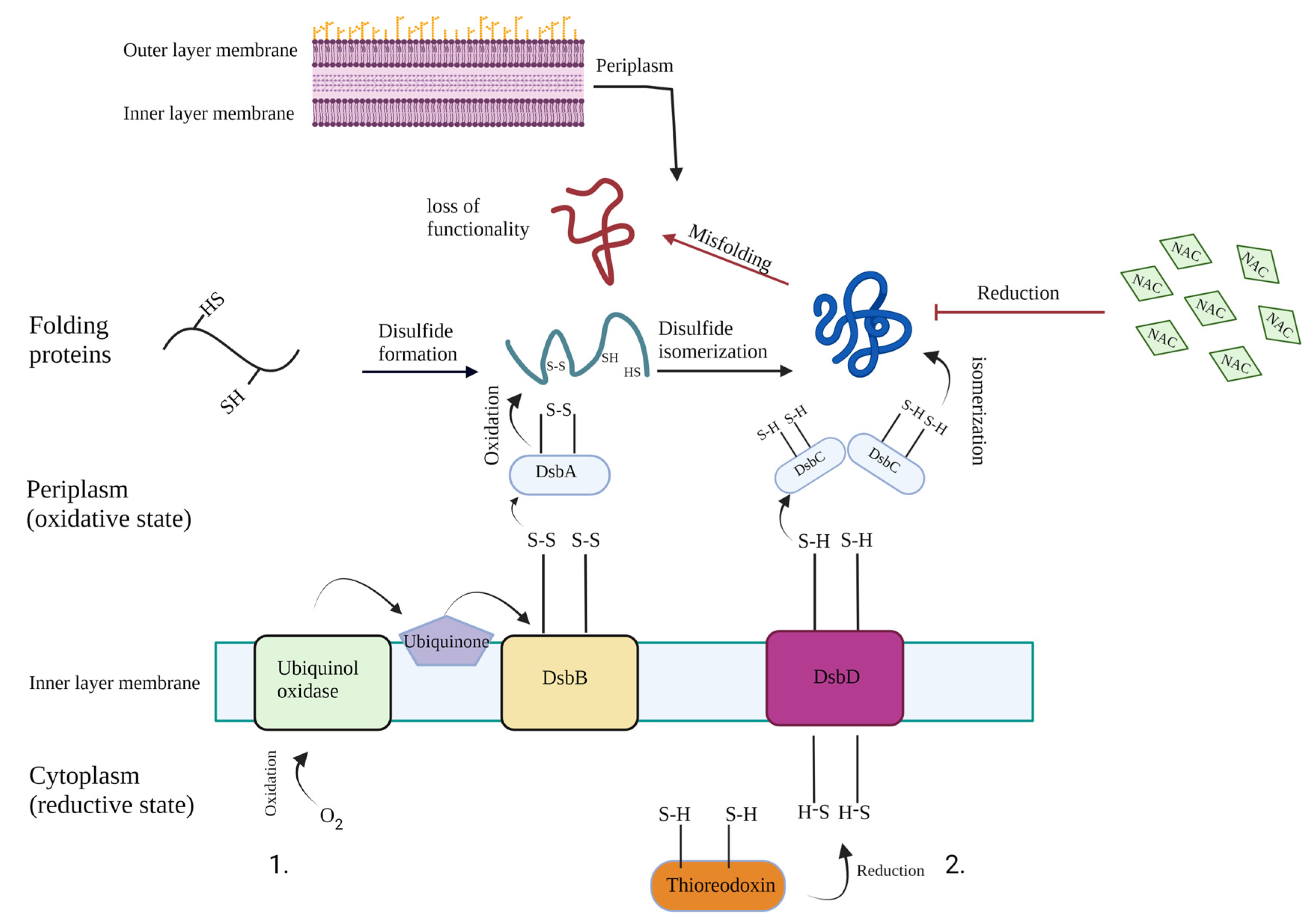
:1. Introduction
2. Results
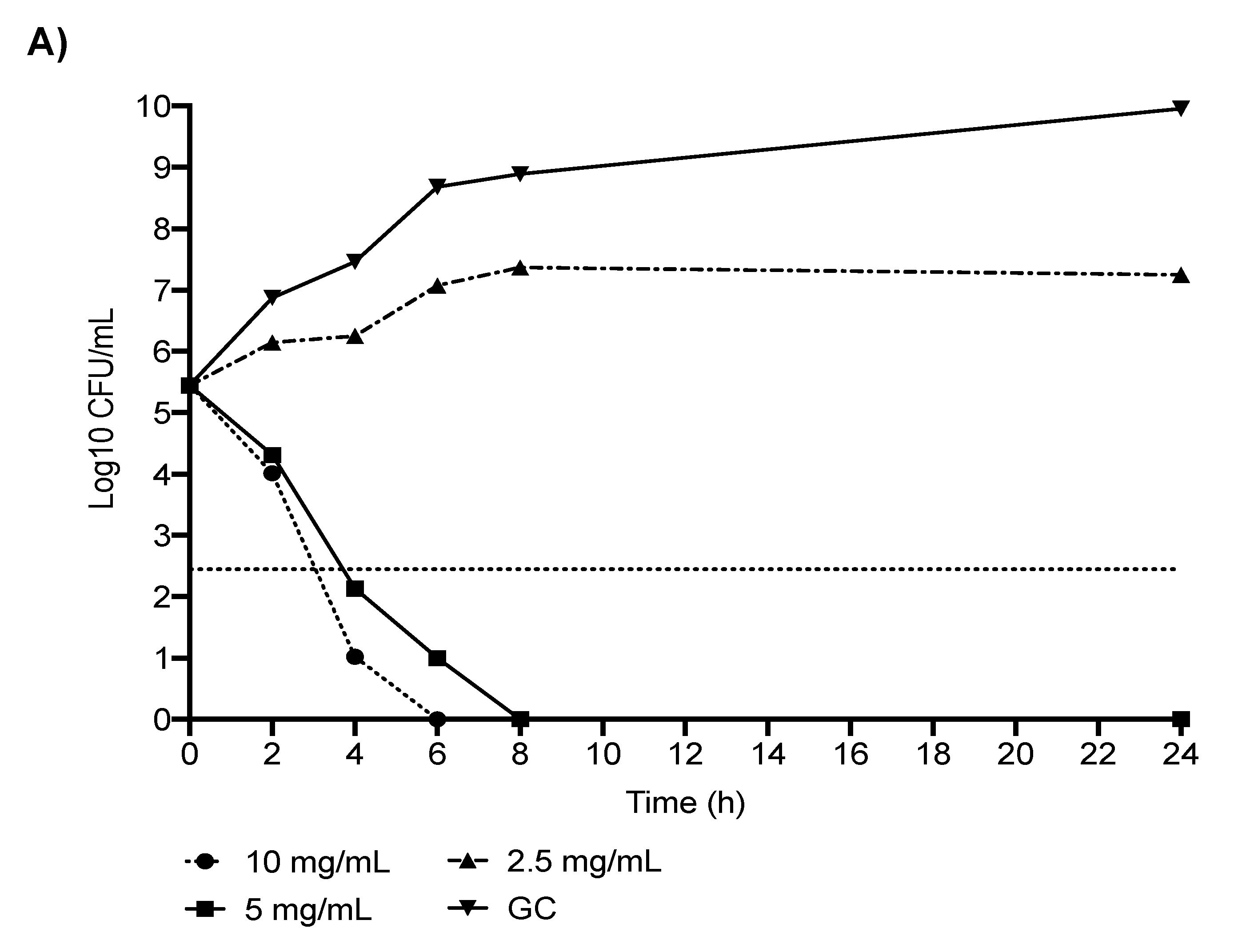
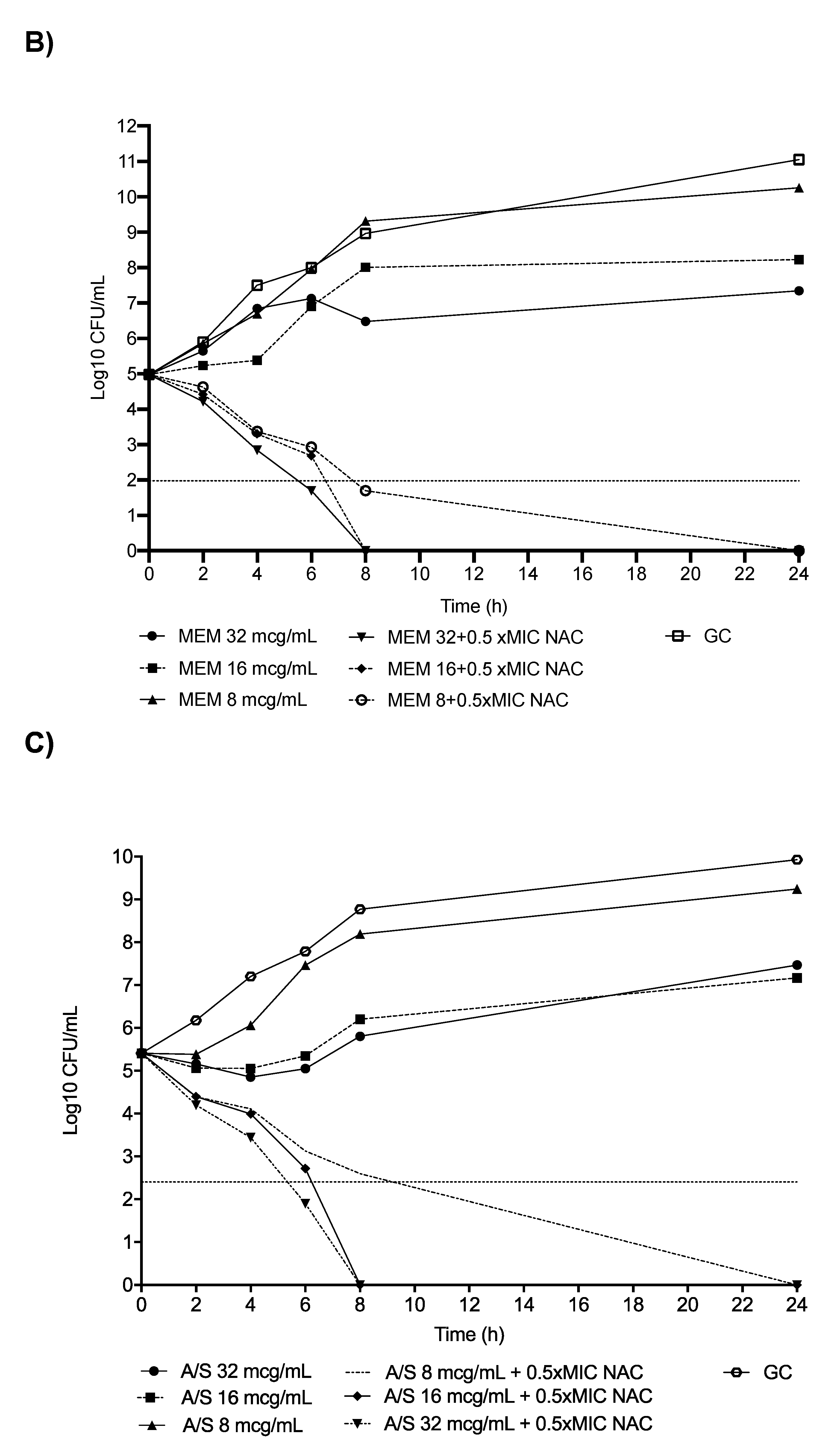
2.1. In Vitro Analyses: CR-Kp
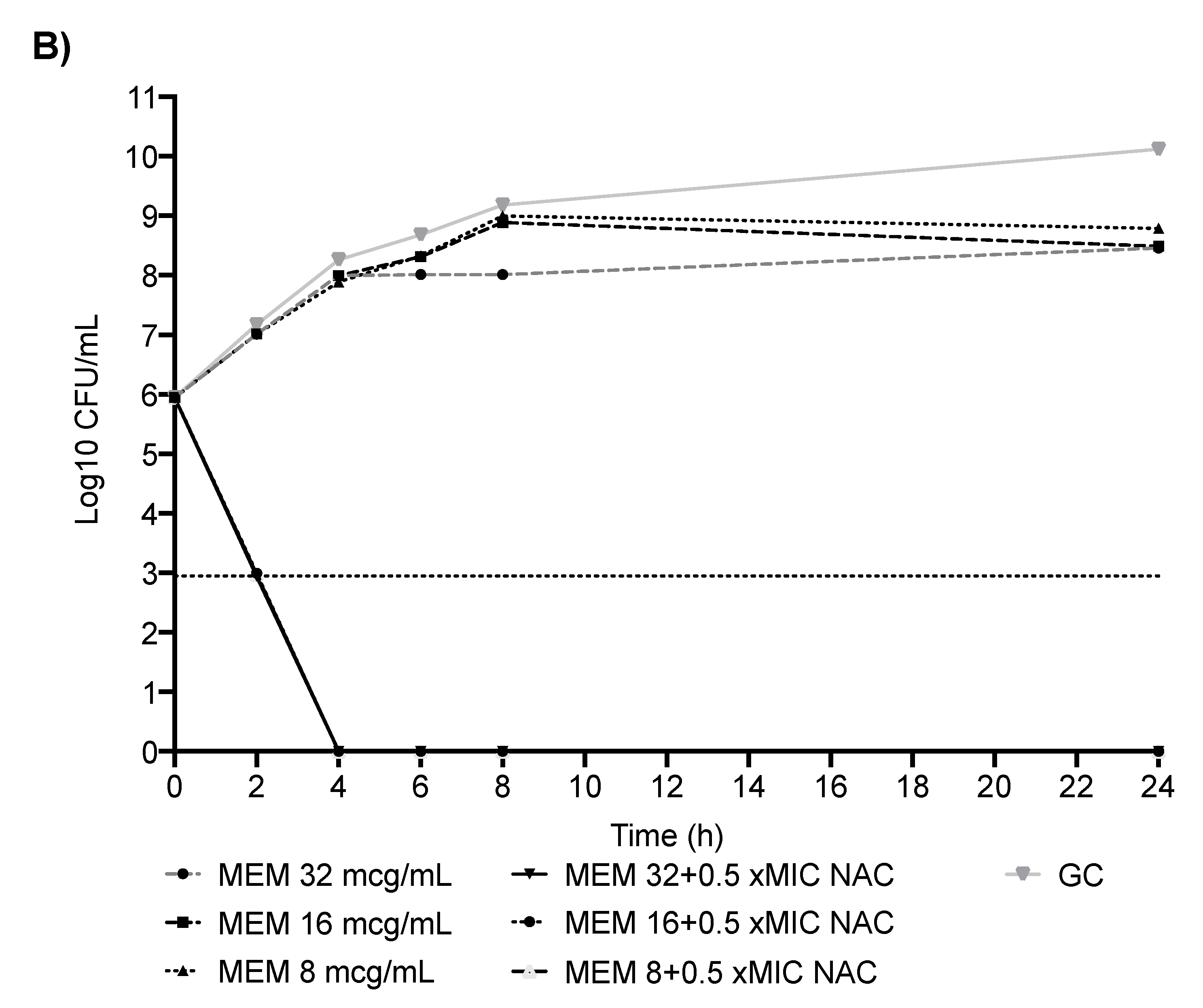
2.2. In Vitro Analyses: CR-Ab
2.3. Scanning Electron Microscopy Analyses
2.4. Serum Bactericidal Activity
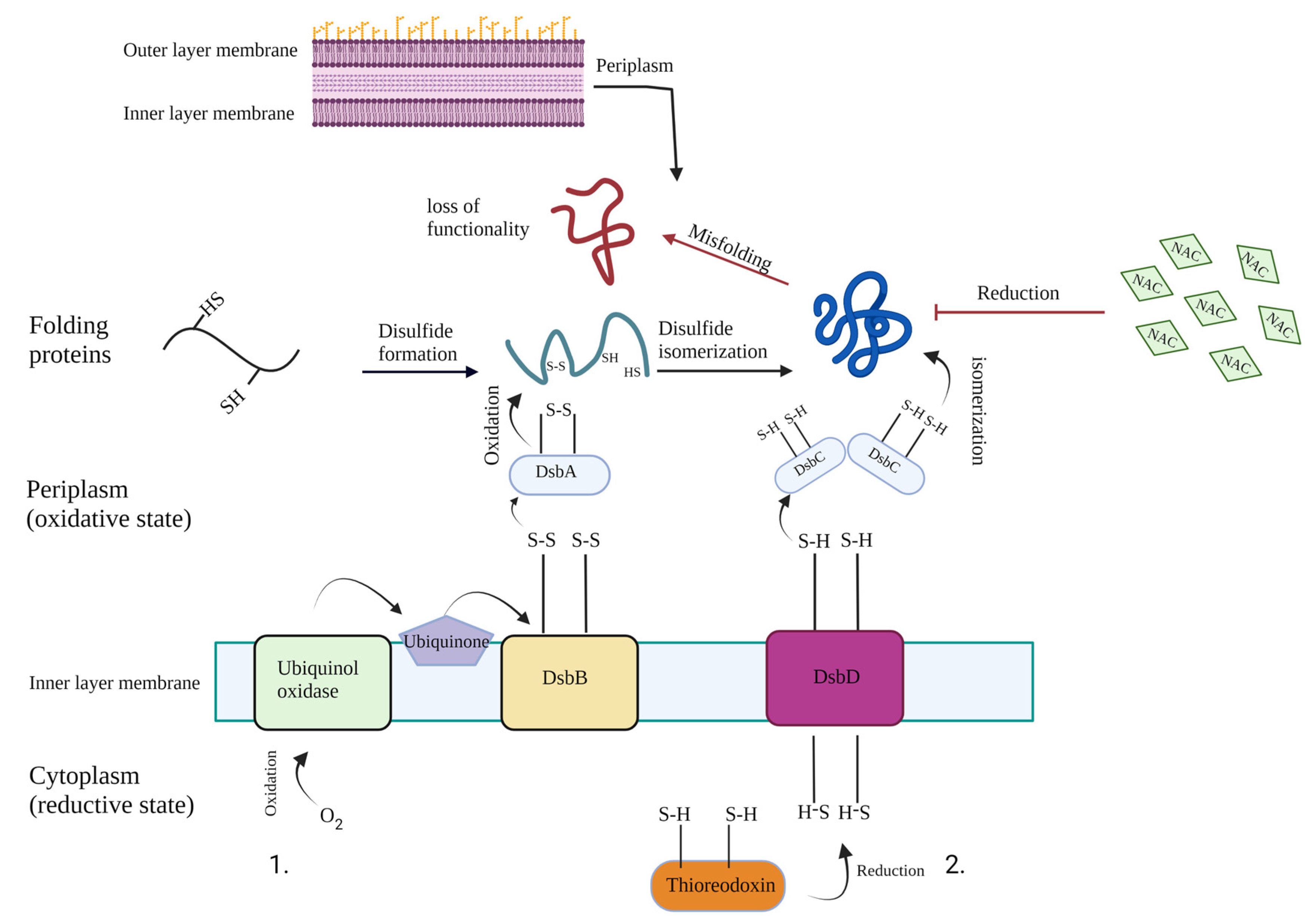
3. Discussion
4. Materials and Methods
4.1. In Vitro Analyses
4.2. Scanning Electron Microscopy Analyses
4.3. Serum Bactericidal Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; de Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of clinical microbiology and infectious diseases (ESCMID) guidelines for the treatment of infections caused by Multidrug-resistant Gram-negative bacilli (endorsed by ESICM-European Society of intensive care Medicine). Clin. Microbiol. Infect. 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Pezzani, M.D. Public health burden of antimicrobial resistance in Europe. Lancet Infect. Dis. 2019, 19, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Kadri, S.S.; Adjemian, J.; Lai, Y.L.; Spaulding, A.B.; Ricotta, E.; Prevots, D.R.; Palmore, T.N.; Rhee, C.; Klompas, M.; Dekker, J.P.; et al. National Institutes of Health Antimicrobial Resistance Outcomes Research Initiative (NIH–ARORI). Difficult-to-Treat Resistance in Gram-negative Bacteremia at 173 US Hospitals: Retrospective Cohort Analysis of Prevalence, Predictors, and Outcome of Resistance to All First-line Agents. Clin. Infect. Dis. 2018, 67, 1803–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busani, S.; Serafini, G.; Mantovani, E.; Venturelli, C.; Giannella, M.; Viale, P.; Mussini, C.; Cossarizza, A.; Girardis, M. Mortality in Patients with Septic Shock by Multidrug Resistant Bacteria: Risk Factors and Impact of Sepsis Treatments. J. Intensive Care Med. 2019, 34, 48–54. [Google Scholar] [CrossRef]
- Russo, A.; Giuliano, S.; Ceccarelli, G.; Alessandri, F.; Giordano, A.; Brunetti, G.; Venditti, M. Comparison of Septic Shock Due to Multidrug-Resistant Acinetobacter baumannii or Klebsiella pneumoniae Carbapenemase-Producing K. pneumoniae in Intensive Care Unit Patients. Antimicrob. Agents Chemother. 2018, 62, e02562-17. [Google Scholar] [CrossRef] [Green Version]
- Oliva, A.; Bianchi, A.; Russo, A.; Ceccarelli, G.; Cancelli, F.; Aloj, F.; Alunni Fegatelli, D.; Mastroianni, C.M.; Venditti, M. Effect of N-Acetylcysteine Administration on 30-Day Mortality in Critically Ill Patients with Septic Shock Caused by Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii: A Retrospective Case-Control Study. Antibiotics 2021, 10, 271. [Google Scholar] [CrossRef]
- Volpicelli, L.; Venditti, M.; Ceccarelli, G.; Oliva, A. Place in Therapy of the Newly Available Armamentarium for Multi-Drug-Resistant Gram-Negative Pathogens: Proposal of a Prescription Algorithm. Antibiotics 2021, 10, 1475. [Google Scholar] [CrossRef]
- Giannella, M.; Bussini, L.; Pascale, R.; Bartoletti, M.; Malagrinò, M.; Pancaldi, L.; Toschi, A.; Ferraro, G.; Marconi, L.; Ambretti, S.; et al. Prognostic Utility of the New Definition of Difficult-to-Treat Resistance among Patients with Gram-Negative Bloodstream Infections. Open Forum Infect. Dis. 2019, 6, ofz505. [Google Scholar] [CrossRef]
- Tenório, M.C.D.S.; Graciliano, N.G.; Moura, F.A.; Oliveira, A.C.M.; Goulart, M.O.F. N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants 2021, 10, 967. [Google Scholar] [CrossRef]
- Pollini, S.; Boncompagni, S.; Di Maggio, T.; Di Pilato, V.; Spanu, T.; Fiori, B.; Blasi, F.; Aliberti, S.; Sergio, F.; Rossolini, G.M.; et al. In vitro synergism of colistin in combination with N-acetylcysteine against Acinetobacter baumannii grown in planktonic phase and in biofilms. J. Antimicrob. Chemother. 2018, 73, 2388–2395. [Google Scholar] [CrossRef] [Green Version]
- Ciacci, N.; Boncompagni, S.; Valzano, F.; Cariani, L.; Aliberti, S.; Blasi, F.; Pollini, S.; Rossolini, G.M.; Pallecchi, L. In Vitro Synergism of Colistin and N-acetylcysteine against Stenotrophomonas maltophilia. Antibiotics 2019, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Blasi, F.; Page, C.; Rossolini, G.M.; Pallecchi, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. The effect of N-acetylcysteine on biofilms: Implications for the treatment of respiratory tract infections. Respir. Med. 2016, 117, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Zafarullah, M.; Li, W.Q.; Sylvester, J.; Ahmad, M. Molecular mechanisms of N-acetylcysteine actions. Cell Mol. Life Sci. 2003, 60, 6–20. [Google Scholar] [CrossRef]
- Parry, M.F.; Neu, H.C. Effect of N-acetylcysteine on antibiotic activity and bacterial growth in vitro. J. Clin. Microbiol. 1977, 5, 58–61. [Google Scholar] [CrossRef]
- Roberts, D.; Cole, P. N-Acetylcysteine potentiates the antipseudomonas activity of carbenicillin in vitro. J. Infect. 1981, 3, 353–359. [Google Scholar] [CrossRef]
- Alfredsson, H.; Malmborg, A.S.; Strandvik, B. N-Acetylcysteine and 2-mercaptoethane sulphonate inhibit anti-pseudomonas activity of antibiotics in vitro. Eur. J. Respir. Dis. 1987, 70, 213–217. [Google Scholar]
- Pollini, S.; Di Pilato, V.; Landini, G.; Di Maggio, T.; Cannatelli, A.; Sottotetti, S.; Cariani, L.; Aliberti, S.; Blasi, F.; Sergio, F.; et al. In vitro activity of N-acetylcysteine against Stenotrophomonas maltophilia and Burkholderia cepacia complex grown in planktonic phase and biofilm. PLoS ONE 2018, 13, e0203941. [Google Scholar] [CrossRef] [Green Version]
- Aiyer, A.; Manoharan, A.; Paino, D.; Farrell, J.; Whiteley, G.S.; Kriel, F.H.; Glasbey, T.O.; Manos, J.; Das, T. Disruption of biofilms and killing of Burkholderia cenocepacia from cystic fibrosis lung using an antioxidant-antibiotic combination therapy. Int. J. Antimicrob. Agents 2021, 58, 106372. [Google Scholar] [CrossRef]
- Alarfaj, R.E.; Alkhulaifi, M.M.; Al-Fahad, A.J.; Aljihani, S.; Yassin, A.E.B.; Alghoribi, M.F.; Halwani, M.A. Antibacterial Efficacy of Liposomal Formulations Containing Tobramycin and N-Acetylcysteine against Tobramycin-Resistant Escherichia coli, Klebsiella pneumoniae, and Acinetobacter baumannii. Pharmaceutics 2022, 14, 130. [Google Scholar] [CrossRef]
- Goswami, M.; Jawali, N. N-acetylcysteine-mediated modulation of bacterial antibiotic susceptibility. Antimicrob. Agents Chemother. 2010, 54, 3529–3530. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.H.; Jang, E.Y.; Shim, K.S.; Lee, J.Y. In vitro effects of N-acetyl cysteine alone and in combination with antibiotics on Prevotella intermedia. J. Microbiol. 2015, 53, 321–329. [Google Scholar] [CrossRef]
- Rodríguez-Beltrán, J.; Cabot, G.; Valencia, E.Y.; Costas, C.; Bou, G.; Oliver, A.; Blázquez, J. N-acetylcysteine selectively antagonizes the activity of imipenem in Pseudomonas aeruginosa by an OprD-mediated mechanism. Antimicrob. Agents Chemother. 2015, 59, 3246–3251. [Google Scholar] [CrossRef] [Green Version]
- Landini, G.; Di Maggio, T.; Sergio, F.; Docquier, J.D.; Rossolini, G.M.; Pallecchi, L. Effect of High N-Acetylcysteine Concentrations on Antibiotic Activity against a Large Collection of Respiratory Pathogens. Antimicrob. Agents Chemother. 2016, 60, 7513–7517. [Google Scholar] [CrossRef] [Green Version]
- Prescott, L.F.; Donovan, J.W.; Jarvie, D.R.; Proudfoot, A.T. The disposition and kinetics of intravenous N-acetylcysteine in patients with paracetamol overdosage. Eur. J. Clin. Pharmacol. 1989, 37, 501–506. [Google Scholar] [CrossRef]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Volgers, C.; Benedikter, B.J.; Grauls, G.E.; Hellebrand, P.H.M.; Savelkoul, P.H.M.; Stassen, F.R.M. Effects of N-acetyl-L-cysteine on the membrane vesicle release and growth of respiratory pathogens. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxman, J.J.; Borg, N.A.; Horne, J.; Thompson, P.E.; Chin, Y.; Sharma, P.; Simpson, J.S.; Wielens, J.; Piek, S.; Kahler, C.M.; et al. The structure of the bacterial oxidoreductase enzyme DsbA in complex with a peptide reveals a basis for substrate specificity in the catalytic cycle of DsbA enzymes. J. Biol. Chem. 2009, 284, 17835–17845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing. Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth microdilution. EUCAST Discussion Document E.Def 2003, 5.1. Clin. Microbiol. Infect. 2003, 9, 1–10. Available online: http://www.escmid.org/Seviware/Script/SvFiles.asp?Ref=359 (accessed on 18 January 2022).
- Cojutti, P.; Sartor, A.; Righi, E.; Scarparo, C.; Bassetti, M.; Pea, F. Population Pharmacokinetics of High-Dose Continuous-Infusion Meropenem and Considerations for Use in the Treatment of Infections Due to KPC-Producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2017, 61, e00794-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdiness, M.R. Clinical pharmacokinetics of N-Acetylcysteine. Clin. Pharmacokinet. 1991, 20, 123–134. [Google Scholar] [CrossRef]
- Stratton, C.W. Serum Bactericidal Test. Clin. Microbiol. Rev. 1988, 1, 19–26. [Google Scholar] [CrossRef]
- Oliva, A.; Ceccarelli, G.; De Angelis, M.; Sacco, F.; Miele, M.C.; Mastroianni, C.M.; Venditti, M. Cefiderocol for compassionate use in the treatment of complicated infections caused by extensively and pan-resistant Acinetobacter baumannii. J. Glob. Antimicrob. Resist. 2020, 23, 292–296. [Google Scholar] [CrossRef]
- Oliva, A.; Miele, M.C.; De Angelis, M.; Costantini, S.; Mascellino, M.T.; Mastroianni, C.M.; Vullo, V.; d’Ettorre, G. Antibacterial Effectiveness of Fecal Water and in Vitro Activity of a Multi-Strain Probiotic Formulation against Multi-Drug Resistant Microorganisms. Microorganisms 2020, 8, 332. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC NAC (mg/mL) | MIC MEM (μg/mL) | MIC COL (μg/mL) | MIC TIG (μg/mL) | MIC RIF (μg/mL) | NAC + MEM FICI | NAC + COL FICI | NAC + TIG FICI | NAC + RIF FICI |
|---|---|---|---|---|---|---|---|---|---|
| #1 | 5 | 256 | 256 | 1 | 16 | 0.25 | >0.5 | >0.5 | >0.5 |
| #2 * | 5 | 1024 | 2048 | 0.5 | 32 | 0.5 | >0.5 | >0.5 | 0.5 |
| #3 | 5 | 16 | 256 | 0.5 | 16 | 0.25 | >0.5 | >0.5 | >0.5 |
| #4 | 5 | 256 | 256 | 1 | 32 | 0.25 | >0.5 | >0.5 | >0.5 |
| #5 | 5 | 256 | 32 | 0.5 | 32 | 0.37 | >0.5 | >0.5 | >0.5 |
| #6 | 5 | 64 | 1024 | 0.5 | 128 | 0.18 | >0.5 | >0.5 | >0.5 |
| #7 | 2.5 | 128 | 1024 | 0.5 | 8 | 0.37 | >0.5 | >0.5 | 0.25 |
| #8 | 5 | 256 | 2 | 0.25 | 16 | 0.25 | >0.5 | >0.5 | >0.5 |
| #9 | 2.5 | 512 | 1 | 0.25 | 8 | 0.5 | >0.5 | >0.5 | >0.5 |
| #10 | 5 | 1024 | 2048 | 4 | 1024 | 0.37 | >0.5 | >0.5 | >0.5 |
| #11 | 5 | 1024 | 8 | 1 | 16 | 0.5 | >0.5 | >0.5 | 0.5 |
| #12 | 2.5 | 128 | 16 | 0.25 | 16 | 0.25 | >0.5 | >0.5 | >0.5 |
| #13 | 5 | 256 | 32 | 0.5 | 256 | 0.18 | >0.5 | >0.5 | >0.5 |
| #14 | 5 | 512 | 256 | 0.5 | 256 | 0.25 | >0.5 | >0.5 | >0.5 |
| #15 | 5 | 512 | 64 | 1 | 32 | 0.5 | >0.5 | >0.5 | >0.5 |
| MIC50/90 | 5/5 | 256/1024 | 256/1024 | 0.5/1 | 32/256 | NA | NA | NA | NA |
| Synergism, n (%) | NA | NA | NA | NA | NA | 15 (100) | 0 (0) | 0 (0) | 3 (20) |
| Strain | MIC NAC (mg/mL) | MIC MEM (μg/mL) | MIC A/S ° (μg/mL) | MIC COL (μg/mL) | MIC TIG (μg/mL) | MIC RIF (μg/mL) | NAC + MEM FICI | NAC + A/S FICI | NAC + COL FICI | NAC + TIG FICI | NAC + RIF FICI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| #1 | 5 | 8 | 16 | 1 | 0.25 | 4 | 0.5 | 0.25 | >0.5 | >0.5 | 0.5 |
| #2 | 2.5 | 64 | 128 | 0.25 | 0.25 | 4 | 0.37 | 0.5 | >0.5 | >0.5 | 0.5 |
| #3 | 2.5 | 128 | 16 | 4 | 0.5 | 2 | 0.25 | 0.18 | >0.5 | >0.5 | 0.25 |
| #4 | 5 | 32 | 16 | 0.5 | 0.5 | 4 | 0.5 | 0.25 | >0.5 | >0.5 | >0.5 |
| #5 | 5 | 512 | 32 | 0.016 | 1 | 32 | 0.25 | 0.18 | >0.5 | >0.5 | 0.5 |
| #6 | 2.5 | 256 | 32 | 0.25 | 0.5 | 4 | 0.37 | 0.37 | >0.5 | >0.5 | 0.37 |
| #7 | 2.5 | 32 | 16 | 8 | 0.5 | 2 | 0.18 | 0.25 | >0.5 | >0.5 | 0.25 |
| #8 * | 2.5 | 1024 | 32 | 4 | 1 | 8 | 0.5 | 0.5 | >0.5 | >0.5 | 0.5 |
| #9 | 2.5 | 128 | 64 | 0.25 | 0.5 | 2 | 0.25 | 0.25 | >0.5 | >0.5 | 0.25 |
| #10 | 5 | 64 | 128 | 0.25 | 0.125 | 2 | 0.37 | 0.25 | >0.5 | >0.5 | 0.18 |
| #11 | 2.5 | 128 | 64 | 1 | 0.5 | 4 | 0.25 | 0.5 | >0.5 | >0.5 | 0.5 |
| #12 | 2.5 | 128 | 64 | 2 | 0.25 | 2 | 0.5 | 0.5 | >0.5 | >0.5 | 0.37 |
| #13 | 2.5 | 256 | 256 | 0.25 | 1 | 8 | 0.37 | 0.25 | >0.5 | >0.5 | 0.5 |
| #14 | 2.5 | 64 | 64 | 0.25 | 0.25 | 2 | 0.25 | 0.5 | >0.5 | >0.5 | 0.25 |
| #15 | 2.5 | 256 | 256 | 0.25 | 0.5 | 2 | 0.18 | 0.37 | >0.5 | >0.5 | 0.25 |
| MIC50/90 | 2.5/5 | 128/512 | 64/256 | 0.25/4 | 0.5/1 | 2/8 | NA | NA | NA | NA | NA |
| Synergism, n (%) | NA | NA | NA | NA | NA | NA | 15 (100) | 15 (100) | 0 (0) | 0 (0) | 14 (93.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Angelis, M.; Mascellino, M.T.; Miele, M.C.; Al Ismail, D.; Colone, M.; Stringaro, A.; Vullo, V.; Venditti, M.; Mastroianni, C.M.; Oliva, A. High Activity of N-Acetylcysteine in Combination with Beta-Lactams against Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii. Antibiotics 2022, 11, 225. https://doi.org/10.3390/antibiotics11020225
De Angelis M, Mascellino MT, Miele MC, Al Ismail D, Colone M, Stringaro A, Vullo V, Venditti M, Mastroianni CM, Oliva A. High Activity of N-Acetylcysteine in Combination with Beta-Lactams against Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii. Antibiotics. 2022; 11(2):225. https://doi.org/10.3390/antibiotics11020225
Chicago/Turabian StyleDe Angelis, Massimiliano, Maria T. Mascellino, Maria C. Miele, Dania Al Ismail, Marisa Colone, Annarita Stringaro, Vincenzo Vullo, Mario Venditti, Claudio M. Mastroianni, and Alessandra Oliva. 2022. "High Activity of N-Acetylcysteine in Combination with Beta-Lactams against Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii" Antibiotics 11, no. 2: 225. https://doi.org/10.3390/antibiotics11020225
APA StyleDe Angelis, M., Mascellino, M. T., Miele, M. C., Al Ismail, D., Colone, M., Stringaro, A., Vullo, V., Venditti, M., Mastroianni, C. M., & Oliva, A. (2022). High Activity of N-Acetylcysteine in Combination with Beta-Lactams against Carbapenem-Resistant Klebsiella pneumoniae and Acinetobacter baumannii. Antibiotics, 11(2), 225. https://doi.org/10.3390/antibiotics11020225