
Variability in Adaptive Resistance of Salmonella Typhimurium to Sublethal Levels of Antibiotics



Abstract
1. Introduction
2. Results and Discussion
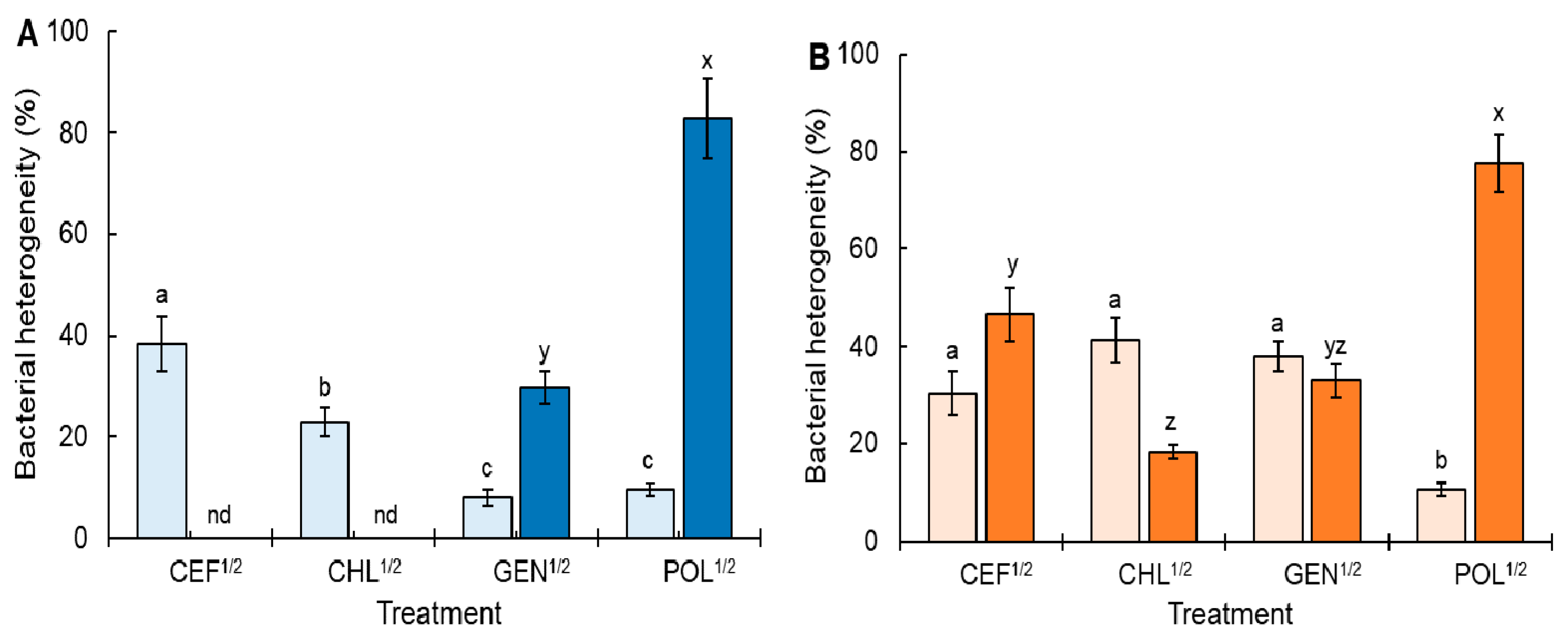
2.1. Antibiotic Susceptibility and Heterogeneous Subpopulation under Continuous Sublethal Selective Pressure
2.2. Cross-Resistance under Continuous Sublethal Selective Pressure
2.3. Relative Fitness under Continuous Sublethal Selective Pressure
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Antibiotic Susceptibility Assay
3.3. Analysis of Heterogeneous Cell Populations
3.4. Disk Diffusion Susceptibility Test
3.5. Relative Fitness
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Hossain, T.; Deter, H.S.; Peters, E.J.; Butzin, N.C. Antibiotic tolerance, persistence, and resistance of the evolved minimal cell, Mycoplasma mycoides JCVI-Syn3B. iScience 2021, 24, 102391. [Google Scholar] [CrossRef] [PubMed]
- Windels, E.M.; Michiels, J.E.; Van den Bergh, B.; Fauvart, M.; Michiels, J. Antibiotics: Combatting tolerance to stop resistance. mBio 2019, 10, e02095-19. [Google Scholar] [CrossRef]
- Windels, E.M.; Bergh, B.V.D.; Michiels, J. Bacteria under antibiotic attack: Different strategies for evolutionary adaptation. PLoS Pathog. 2020, 16, e1008431. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, L.; Fernandez-Cuenca, F.; Blasco, L.; López-Rojas, R.; Ambroa, A.; Lopez, M.; Pascual, Á.; Bou, G.; Tomás, M. Relationship between tolerance and persistence mechanisms in Acinetobacter baumannii strains with AbkAB toxin-antitoxin system. Antimicrob. Agents Chemother. 2018, 62, e00250-18. [Google Scholar] [CrossRef]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef] [PubMed]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.; Wright, G.D. Intrinsic antibiotic resistance: Mechanisms, origins, challenges and solutions. Int. J. Med. Microbiol. 2013, 303, 287–292. [Google Scholar] [CrossRef]
- Wilmaerts, D.; Windels, E.; Verstraeten, N.; Michiels, J. General Mechanisms Leading to Persister Formation and Awakening. Trends Genet. 2019, 35, 401–411. [Google Scholar] [CrossRef]
- Rocha-Granados, M.C.; Zenick, B.; Englander, H.E.; Mok, W.W. The social network: Impact of host and microbial interactions on bacterial antibiotic tolerance and persistence. Cell. Signal. 2020, 75, 109750. [Google Scholar] [CrossRef]
- Goormaghtigh, F.; Melderen, L.V. Single-Cell imaging and characterization of Escherichia coli persister cells to ofloxacin in exponential cultures. Sci Adv. 2019, 5, eaav9462. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jo, A.; Ding, T.; Lee, H.-Y.; Ahn, J. Assessment of altered binding specificity of bacteriophage for ciprofloxacin-induced antibiotic-resistant Salmonella Typhimurium. Arch. Microbiol. 2016, 198, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Veeraraghavan, B.; Elangovan, R.; Perumal, V. Antibiotic resistance and epigenetics: More to it than meets the eye. Antimicrob. Agents Chemother. 2020, 64, e02225-19. [Google Scholar] [CrossRef] [PubMed]
- Völzing, K.G.; Brynildsen, M.P. Stationary-Phase Persisters to Ofloxacin Sustain DNA Damage and Require Repair Systems Only during Recovery. mBio 2015, 6, e00731-15. [Google Scholar] [CrossRef]
- Long, Y.; Fu, W.; Li, S.; Ren, H.; Li, M.; Liu, C.; Zhang, B.; Xia, Y.; Fan, Z.; Xu, C.; et al. Identification of novel genes that promote persister formation by repressing transcription and cell division in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2019, 74, 2575–2587. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial Persistence as a Phenotypic Switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef]
- Zou, J.; Peng, B.; Qu, J.; Zheng, J. Are bacterial persisters dormant cells only? Front. Microbiol. 2021, 12, 708580. [Google Scholar] [CrossRef]
- Liu, S.; Wu, N.; Zhang, S.; Yuan, Y.; Zhang, W.; Zhang, Y. Variable persister gene interactions with (p)ppGpp for persister formation in Escherichia coli. Front. Microbiol. 2017, 8, 1795. [Google Scholar] [CrossRef]
- Claudi, B.; Spröte, P.; Chirkova, A.; Personnic, N.; Zankl, J.; Schürmann, N.; Schmidt, A.; Bumann, D. Phenotypic Variation of Salmonella in Host Tissues Delays Eradication by Antimicrobial Chemotherapy. Cell 2014, 158, 722–733. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Debbia, E.; Roveta, S.; Schito, A.; Gualco, L.; Marchese, A. Antibiotic Persistence: The Role of Spontaneous DNA Repair Response. Microb. Drug Resist. 2001, 7, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Trastoy, R.; Manso, T.; Fernández-García, L.; Blasco, L.; Ambroa, A.; del Molino, M.L.P.; Bou, G.; García-Contreras, R.; Wood, T.K.; Tomás, M. Mechanisms of bacterial tolerance and persistence in the gastrointestinal and respiratory environments. Clin. Microbiol. Rev. 2018, 31, e00023-18. [Google Scholar] [CrossRef]
- Goneau, L.W.; Yeoh, N.S.; MacDonald, K.W.; Cadieux, P.A.; Burton, J.P.; Razvi, H.; Reid, G. Selective Target Inactivation Rather than Global Metabolic Dormancy Causes Antibiotic Tolerance in Uropathogens. Antimicrob. Agents Chemother. 2014, 58, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Wood, T.K. Toxins Hha and CspD and small RNA regulator Hfq are involved in persister cell formation through MqsR in Escherichia coli. Biochem. Biophys. Res. Commun. 2010, 391, 209–213. [Google Scholar] [CrossRef]
- Helaine, S.; Kugelberg, E. Bacterial persisters: Formation, eradication, and experimental systems. Trends Microbiol. 2014, 22, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Kaldalu, N.; Hauryliuk, V.; Tenson, T. Persisters—As elusive as ever. Appl. Microbiol. Biotechnol. 2016, 100, 6545–6553. [Google Scholar] [CrossRef]
- Dewachter, L.; Verstraeten, N.; Fauvart, M.; Michiels, J. An integrative view of cell cycle control in Escherichia coli. FEMS Microbiol. Rev. 2018, 42, 116–136. [Google Scholar] [CrossRef]
- Page, R.; Peti, W. Toxin-Antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef]
- Podlesek, Z.; Bertok, D.Ž. The DNA Damage Inducible SOS Response Is a Key Player in the Generation of Bacterial Persister Cells and Population Wide Tolerance. Front. Microbiol. 2020, 11, 1785. [Google Scholar] [CrossRef]
- Chang, J.; Lee, R.-E.; Lee, W. A pursuit of Staphylococcus aureus continues: A role of persister cells. Arch. Pharmacal Res. 2020, 43, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, R.; Ryan, S.S.; McCoy, A.J.; Gunn, J.S. Identification and genetic characterization of PmrA-regulated genes and genes involved in polymyxin B resistance in Salmonella enterica serovar typhimurium. Infect. Immun. 2002, 70, 6770–6778. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.E.; Van den Bergh, B.; Verstraeten, N.; Michiels, J. Molecular mechanisms and clinical implications of bacterial persistence. Drug Resist. Updat. 2016, 29, 76–89. [Google Scholar] [CrossRef]
- Chellat, M.F.; Raguž, L.; Riedl, R. Targeting Antibiotic Resistance. Angew. Chem. Int. Ed. 2016, 55, 6600–6626. [Google Scholar] [CrossRef] [PubMed]
- Wistrand-Yuen, E.; Knopp, M.; Hjort, K.; Koskiniemi, S.; Berg, O.G.; Andersson, D.I. Evolution of high-level resistance during low-level antibiotic exposure. Nat. Commun. 2018, 9, 1599. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, J.; Swaminath, S.; Nair, R.R.; Jakkala, K.; Pradhan, A.; Ajitkumar, P. De Novo Emergence of Genetically Resistant Mutants of Mycobacterium tuberculosis from the Persistence Phase Cells Formed against Antituberculosis Drugs In Vitro. Antimicrob. Agents Chemother. 2017, 61, e01343-16. [Google Scholar] [CrossRef] [PubMed]
- Dörr, T.; Davis, B.M.; Waldor, M.K. Endopeptidase-Mediated Beta Lactam Tolerance. PLoS Pathog. 2015, 11, e1004850. [Google Scholar] [CrossRef]
- Pu, Y.; Zhao, Z.; Li, Y.; Zou, J.; Ma, Q.; Zhao, Y.; Ke, Y.; Zhu, Y.; Chen, H.; Baker, M.A.; et al. Enhanced Efflux Activity Facilitates Drug Tolerance in Dormant Bacterial Cells. Mol. Cell 2016, 62, 284–294. [Google Scholar] [CrossRef]
- Bergmiller, T.; Andersson, A.M.; Tomasek, K.; Balleza, E.; Kiviet, D.J.; Hauschild, R.; Tkačik, G.; Guet, C.C. Biased partitioning of the multidrug efflux pump AcrAB-TolC underlies long-lived phenotypic heterogeneity. Science 2017, 356, 311–315. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Aulin, L.B.S.; Buffoni, M.; Fragkiskou, E.; van Hasselt, J.G.C.; Rozen, D.E. Allele-specific collateral and fitness effects determine the dynamics of fluoroquinolone resistance evolution. Proc. Natl. Acad. Sci. USA 2022, 119, e2121768119. [Google Scholar] [CrossRef]
- Edelmann, D.; Berghoff, B.A. A Shift in Perspective: A Role for the Type I Toxin TisB as Persistence-Stabilizing Factor. Front. Microbiol. 2022, 13, 871699. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.; Trebosc, V.; Kemmer, C.; Rosenstiel, P.; Beardmore, R.; Schulenburg, H.; Jansen, G. Alternative Evolutionary Paths to Bacterial Antibiotic Resistance Cause Distinct Collateral Effects. Mol. Biol. Evol. 2017, 34, 2229–2244. [Google Scholar] [CrossRef] [PubMed]
- Baym, M.; Stone, L.K.; Kishony, R. Multidrug evolutionary strategies to reverse antibiotic resistance. Science 2016, 351, aad3292. [Google Scholar] [CrossRef] [PubMed]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Kussell, E.; Leibler, S. Phenotypic Diversity, Population Growth, and Information in Fluctuating Environments. Science 2005, 309, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, S.; van Leeuwe, T.M.; Qachach, O.; Zhang, Z.; van Wezel, G.P.; Rozen, D.E. The evolution of no-cost resistance at sub-MIC concentrations of streptomycin in Streptomyces coelicolor. ISME J. 2017, 11, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Prax, M.; Mechler, L.; Weidenmaier, C.; Bertram, R. Glucose Augments Killing Efficiency of Daptomycin Challenged Staphylococcus aureus Persisters. PLoS ONE 2016, 11, e0150907. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Huang, X.; Wang, X.; Yan, S.; Guo, S.; Abdalla, A.E.; Huang, C.; Xie, J. l-Serine potentiates fluoroquinolone activity against Escherichia coli by enhancing endogenous reactive oxygen species production. J. Antimicrob. Chemother. 2016, 71, 2192–2199. [Google Scholar] [CrossRef]
- Lebeaux, D.; Chauhan, A.; Létoffé, S.; Fischer, F.; de Reuse, H.; Beloin, C.; Ghigo, J.M. pH-Mediated potentiation of aminoglycosides kills bacterial persisters and eradicates In vivo biofilms. J. Infect. Dis. 2014, 210, 1357–1366. [Google Scholar] [CrossRef]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Khan, S.; Beattie, T.K.; Knapp, C.W. The use of minimum selectable concentrations (MSCs) for determining the selection of antimicrobial resistant bacteria. Ecotoxicology 2017, 26, 283–292. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Antibiotic | Spectrum | Activity | Affinity | Inhibitory Mechanisms |
|---|---|---|---|---|---|
| Cephems | Cefotaxime | Broad | Cidal | Hydrophilic | Cell wall synthesis |
| Aminoglycosides | Gentamicin | Narrow | Cidal | Hydrophilic | Protein synthesis (30S) |
| Glycopeptides | Polymyxin B | Narrow | Cidal | Hydrophilic | Membrane permeability |
| Phenicols | Chloramphenicol | Broad | Static | Hydrophobic | Protein synthesis (50S) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dawan, J.; Ahn, J. Variability in Adaptive Resistance of Salmonella Typhimurium to Sublethal Levels of Antibiotics. Antibiotics 2022, 11, 1725. https://doi.org/10.3390/antibiotics11121725
Dawan J, Ahn J. Variability in Adaptive Resistance of Salmonella Typhimurium to Sublethal Levels of Antibiotics. Antibiotics. 2022; 11(12):1725. https://doi.org/10.3390/antibiotics11121725
Chicago/Turabian StyleDawan, Jirapat, and Juhee Ahn. 2022. "Variability in Adaptive Resistance of Salmonella Typhimurium to Sublethal Levels of Antibiotics" Antibiotics 11, no. 12: 1725. https://doi.org/10.3390/antibiotics11121725
APA StyleDawan, J., & Ahn, J. (2022). Variability in Adaptive Resistance of Salmonella Typhimurium to Sublethal Levels of Antibiotics. Antibiotics, 11(12), 1725. https://doi.org/10.3390/antibiotics11121725