
A Water-Soluble Antibiotic in Rhubarb Stalk Shows an Unusual Pattern of Multiple Zones of Inhibition and Preferentially Kills Slow-Growing Bacteria

Abstract
1. Introduction
2. Results
2.1. Antibacterial Activity of Aqueous Extract of Rhubarb Stalk
2.2. No Antibacterial Activity Present in the Organic Extract of Rhubarb Stalk
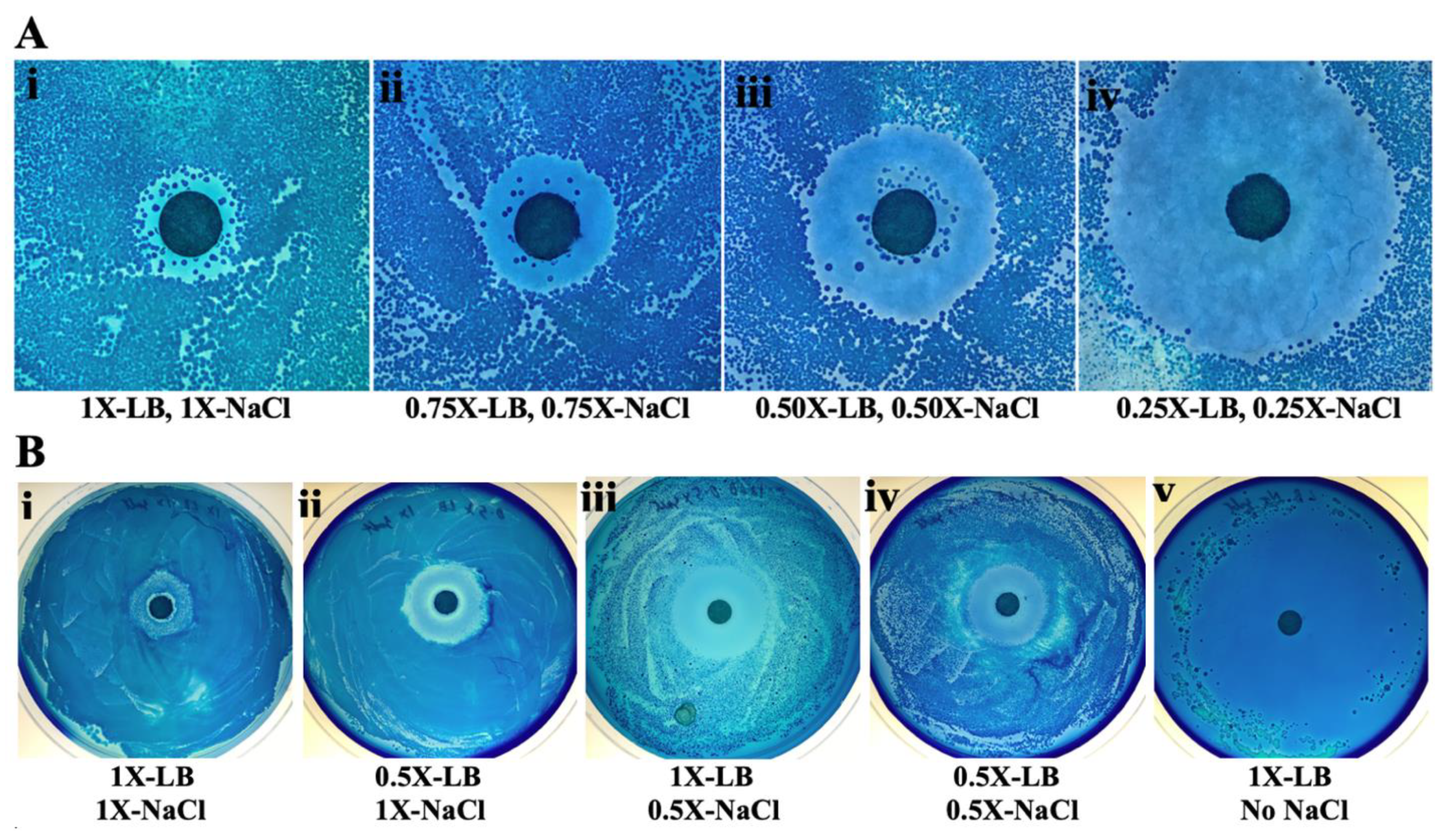
2.3. Increased Antibiotic Activity at Low Nutrient and Salt Concentrations
2.4. Unusual Multiple Zones of Inhibition
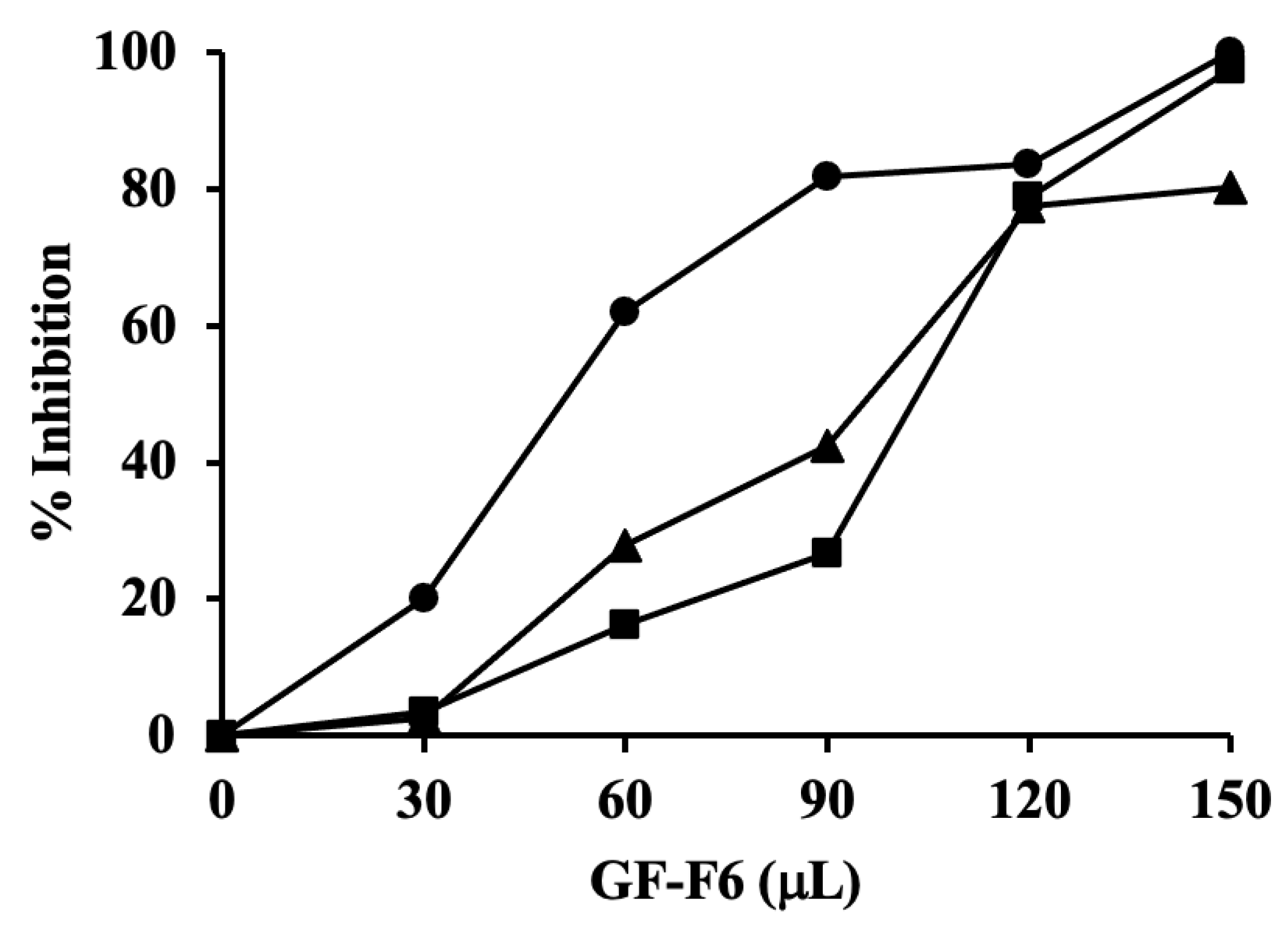
2.5. Chromatographic Purification of the Antibiotic in Rhubarb Stalk
2.6. Broad Spectrum Activity of the Antibiotic in Rhubarb Stalk
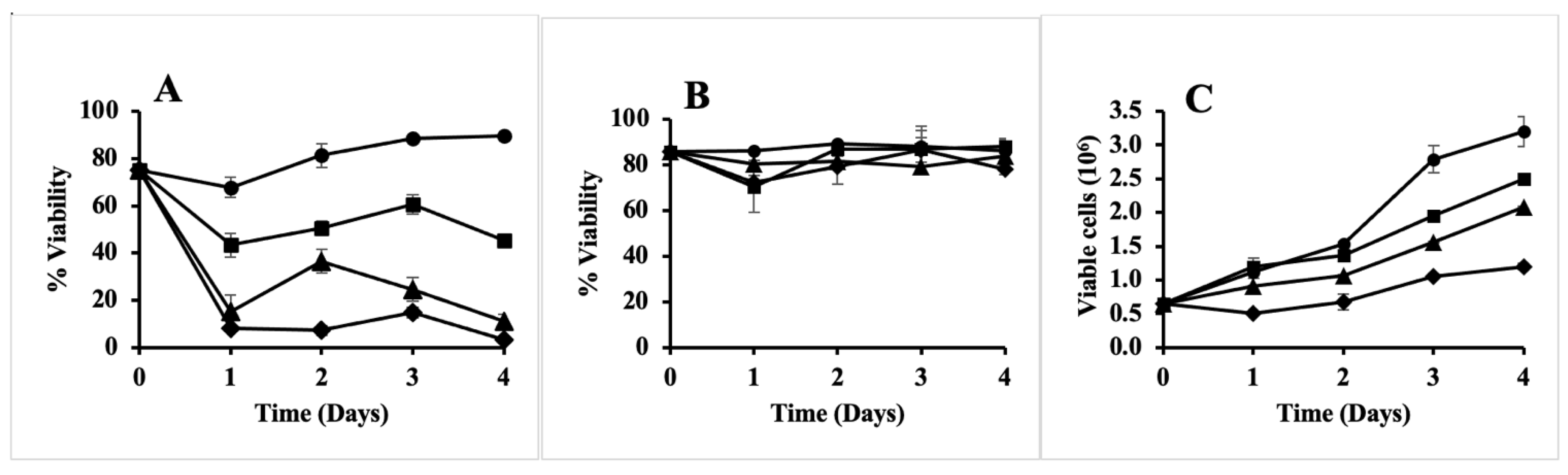
2.7. The Antibiotic in Rhubarb Stalk Is Bactericidal
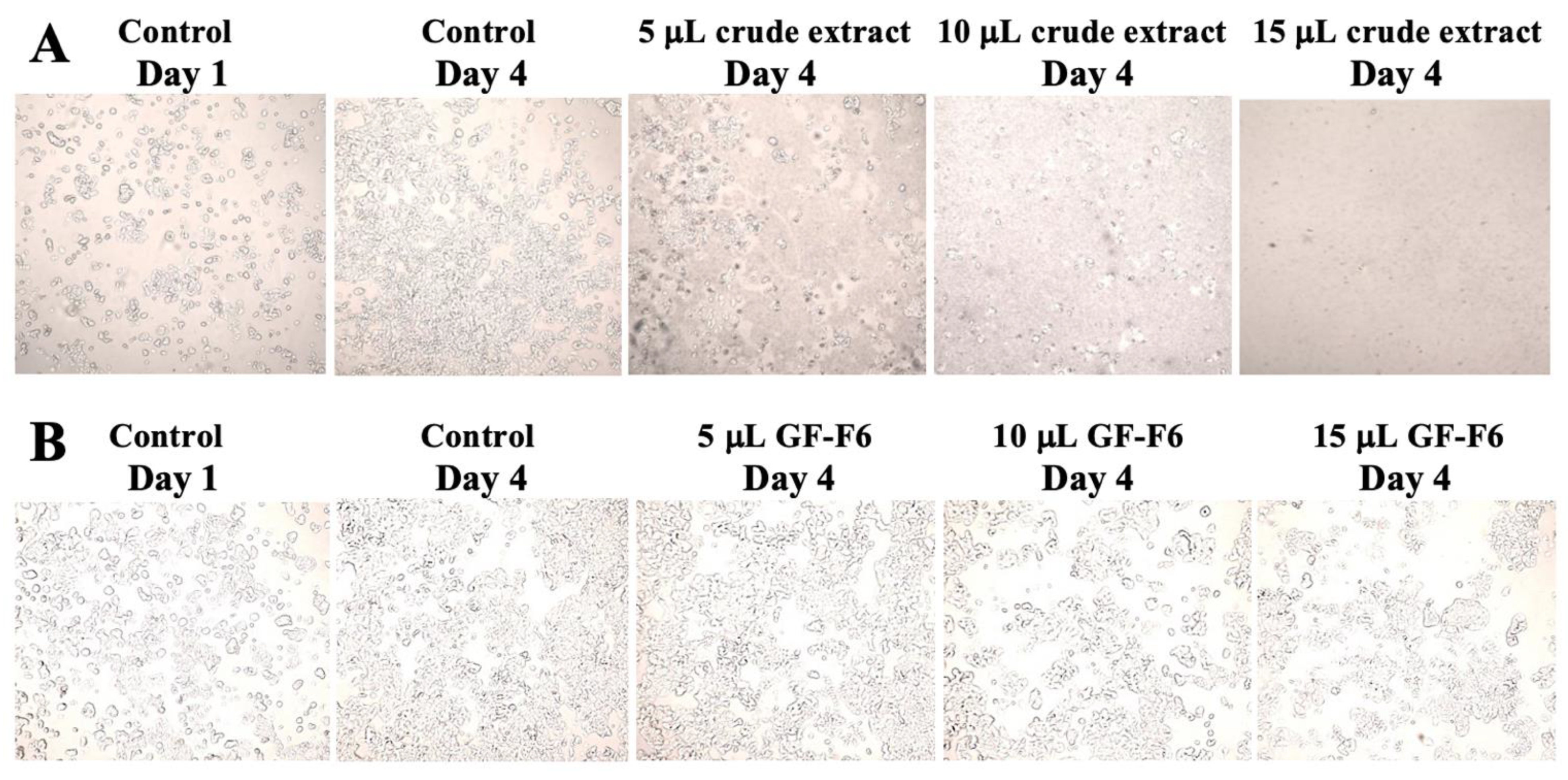
2.8. Anti-Proliferative Activity, but No Cytotoxicity of the Antibiotic in Rhubarb
3. Discussion
4. Materials and Methods
4.1. Preparation of Rhubarb Extract
4.2. Disk Diffusion Assays
4.3. Chromatographic Techniques
4.4. Bacterial Strains, Culture Conditions, and Inhibition Studies
4.5. Determination of the Rate of Killing of A. actinomycetemcomitans
4.6. MCF-7 Cell Culture
4.6.1. Standard Cell Culture Protocols
4.6.2. Determination of Anti-Proliferative and Cytotoxic Effects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Alibi, S.; Crespo, D.; Navas, J. Plant-derivatives small molecules with antibacterial activity. Antibiotics 2021, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Lai, P.K.; Roy, J. Antimicrobial and chemopreventive properties of herbs and spices. Curr. Med. Chem. 2004, 11, 1451–1460. [Google Scholar] [CrossRef]
- Mou, X.; Kesari, S.; Wen, P.Y.; Huang, X. Crude drugs as anticancer agents. Int. J. Clin. Exp. Med. 2011, 4, 17–25. [Google Scholar] [PubMed]
- Jain, R.; Jain, S. Traditional medicinal plants as anticancer agents from Chhattishgarh, India: An overview. Int. J. Phytomed. 2010, 2, 186–196. [Google Scholar] [CrossRef]
- Slaughter, R.J.; Beasley, D.M.; Lambie, B.S.; Wilkins, G.T.; Schep, L.J. Poisonous plants in New Zealand: A review of those that are most commonly enquired about to the National Poisons Centre. N. Z. Med. J. 2012, 125, 87–118. [Google Scholar] [PubMed]
- Kolodziejczyk-Czepas, J.; Liudvytska, O. Rheum rhaponticum and Rheum rhabarbarum: A review of phytochemistry, biological activities and therapeutic potential. Phytochem. Rev. 2021, 20, 589–607. [Google Scholar] [CrossRef]
- Zargar, B.A.; Masoodi, M.H.; Ahmed, B.; Ganie, S.A. Phytoconstituents and therapeutic uses of Rheum emodi wall. ex Meissn. Food Chem. 2011, 128, 585–589. [Google Scholar] [CrossRef]
- Babu, K.S.; Srinivas, P.V.; Praveen, B.; Kishore, K.S.; Murty, U.S.; Rao, J.M. Antimicrobial constituents from the rhizomes of Rheum emodi. Phytochemistry 2003, 62, 203–207. [Google Scholar] [CrossRef]
- Chinsembu, K.C. Plants and other natural products used in the management of oral infections and improvement of oral health. Acta Trop. 2016, 154, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Lu, G.; Shen, H.-M.; Chung, M.C.M.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, M.K. Antimetabolites: Antibiotics that inhibit nucleotide synthesis. In Chemistry of Antibiotics and Related Drugs; Springer Nature: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Traxler, P.; Gruner, J.; Auden, J.A. Papulacandins, a new family of antibiotics with antifungal activity: Fermentation, isolation, chemical and biological characterization of Papulacandins A, B, C, D and E. J. Antibiot. 1976, 30, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Matsuura, I.; Nawata, Y.; Endo, H.; Sasaki, H.; Okytomi, T.; Saehi, T.; Tamura, G. Funiculosin, a new antibiotic: II. Structure elucidation and antifungal activity. J. Antibiot. 1978, 31, 533–538. [Google Scholar] [CrossRef]
- Svetlov, M.S.; Koller, T.O.; Meydan, S.; Shankar, V.; Klepacki, D.; Polacek, N.; Guydosh, N.R.; Vazquez-Laslop, N.; Wilson, D.N.; Mankin, A.S. Context-specific action of macrolide antibiotics on the eukaryotic ribosome. Nat. Commun. 2021, 12, 2803–2816. [Google Scholar] [CrossRef]
- Colson, P.; Raoult, D. Fighting viruses with antibiotics: An overlooked path. Int. J. Antimicrob. Agents 2016, 48, 349–352. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, R.; Li, G.; Gao, Q.; Yuan, S.; Altmeyer, R.; Zou, G. Teicoplanin inhibits Ebola pseudovirus infection in cell culture. Antivir. Res. 2016, 125, 1–7. [Google Scholar] [CrossRef]
- Varghese, F.S.; Kaukinen, P.; Glasker, S.; Bespalov, M.; Hanski, L.; Wennerberg, K.; Kummerer, B.M.; Ahola, T. Discovery of berberine, abamectin and ivermectin as antivirals against chikungunya and other alphaviruses. Antivir. Res. 2016, 126, 117–124. [Google Scholar] [CrossRef]
- Poddighe, D.; Aljofan, M. Clinical evidences on the antiviral properties of macrolide antibiotics in the COVID-19 era and beyond. Antivir. Chem. Chemother. 2020, 28, 2040206620961712. [Google Scholar] [CrossRef]
- Galm, U.; Hager, M.H.; Van Lanen, S.G.; Ju, J.; Thorson, J.S.; Shen, B. Antitumor antibiotics: Bleomycin, enediynes, and mitomycin. Chem. Rev. 2005, 105, 739–758. [Google Scholar] [CrossRef]
- Bhattacharya, B.; Mukherjee, S. Cancer therapy using antibiotics. J. Cancer Ther. 2015, 6, 849–858. [Google Scholar] [CrossRef]
- Karpinski, T.M.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Shang, Q.; Li, W.; Guo, W.; Stojadinovic, A.; Mannion, C.; Man, Y.-G.; Chen, T. Antibiotics for cancer treatment: A double-edged sword. J. Cancer 2020, 11, 5135–5149. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant Products as Antimicrobial Agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Bakal, S.N.; Bereswill, S.; Heimesaat, M.M. Finding novel antibiotic substances from medicinal plants–antimicrobial properties of Nigella sativa directed against multidrug-resistant bacteria. Eur. J. Microbiol. Immunol. 2017, 7, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Prasad, M.A.; Zolnik, C.P.; Molina, J. Leveraging phytochemicals: The plant phylogeny predicts sources of novel antibacterial compounds. Future Sci. OA 2019, 5, FSO407. [Google Scholar] [CrossRef] [PubMed]
- Benzaid, C.; Belmadani, A.; Tichati, L.; Djeribi, R.; Rouabhia, M. Effect of Citrus aurantium L. Essential Oil on Streptococcus mutans Growth, Biofilm Formation and Virulent Genes Expression. Antibiotics 2021, 10, 54. [Google Scholar] [CrossRef]
- Puvaca, N.; Milenkovic, J.; Galonja Coghill, T.; Bursic, V.; Petrovic, A.; Tanaskovic, S.; Pelic, M.; Ljubojevic Pelic, D.; Miljkovic, T. Antimicrobial Activity of Selected Essential Oils against Selected Pathogenic Bacteria: In Vitro Study. Antibiotics 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Rogawansamy, S.; Gaskin, S.; Taylor, M.; Pisaniello, D. An Evaluation of Antifungal Agents for the Treatment of Fungal Contamination in Indoor Air Environments. Int. J. Environ. Res. Public Health 2015, 12, 6319–6332. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Ermolenko, E.; Savidov, N.; Gloriozova, T.A.; Poroikov, V.V. Antiprotozoal and antitumor activity of natural polycyclic endoperoxides: Origin, structures and biological activity. Molecules 2021, 26, 686. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-T.; Ke, C.-Y.; Wu, W.-T.; Tseng, Y.-H.; Lee, R.-P. Antimicrobial and anti-inflammatory potential of Angelica dahurica and Rheum officinale extract accelerates wound healing in Staphylococcus aureus-infected wounds. Sci. Rep. 2020, 10, 5596. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-Y.; Wy, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Knudsen, J.D.; Frimodt-Møller, N.; Espersen, F. Pharmacodynamics of penicillin are unaffected by bacterial growth phases of Streptococcus pneumoniae in the mouse peritonitis model. J. Antimicrob. Chemother. 1998, 41, 451–459. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Eng, R.H.K.; Padberg, F.T.; Smith, S.M.; Tan, E.N.; Cherubin, C.E. Bactericidal effects of antibiotics on slowly growing and nongrowing bacteria. Antimicrob. Agents Chemother. 1991, 35, 1824–1828. [Google Scholar] [CrossRef] [PubMed]
- McCall, I.C.; Shah, N.; Govindan, A.; Baquero, F.; Levin, B.R. Antibiotic killing of diversely generated populations of nonreplicating bacteria. Antimicrob. Agents Chemother. 2019, 63, e02360-18. [Google Scholar] [CrossRef]
- Bhattacharjee, M.K.; Alenezi, T. Antibiotic in myrrh from Commiphora molmol preferentially kills nongrowing bacteria. Future Sci. OA 2020, 6, FSO458. [Google Scholar] [CrossRef]
- Shrestha, P.; Ni, J.; Wong, T.-Y. Synergistic and antagonistic interactions of triclosan with various antibiotics in bacteria. J. Environ. Sci. Health Part C 2020, 38, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Schwalbe, R.S.; Ritz, W.J.; Verma, P.R.; Barranco, E.A.; Gilligan, P.H. Selection for vancomycin resistance in clinical isolates of Staphylococcus haemolyticus. J. Infect. Dis. 1990, 161, 45–51. [Google Scholar] [CrossRef]
- Baker, N.C.; Elkins, S.; Williams, A.J.; Tropsha, A. A bibliometric review of drug repurposing. Drug Discov. Today 2018, 23, 661–672. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Guo, D.W.; Lao, Q.C.; Xu, Y.Q.; Meng, Z.K.; Xia, B.; Yang, H.; Li, C.Q.; Li, P. Sensitization and synergistic anti-cancer effects of Furanodiene identified in zebrafish models. Sci. Rep. 2019, 9, 4541. [Google Scholar] [CrossRef]
- Marostica, L.L.; Silva, I.T.; Kratz, J.M.; Persich, L.; Geller, F.C.; Lang, K.L.; Caro, M.S.B.; Duran, F.J.; Schenkel, E.P.; Simoes, C.M.O. Synergistic Antiproliferative Effects of a New Cucurbitacin B Derivative and Chemotherapy Drugs on Lung Cancer Cell Line A549. Chem. Res. Toxicol. 2015, 28, 1949–1960. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, M.K. Better visualization and photodocumentation of zone of inhibition by staining cells and background agar differently. J. Antibiot. 2015, 68, 657–659. [Google Scholar] [CrossRef]
- Bhattacharjee, M.K.; Sugawara, K.; Ayandeji, O.T. Microwave sterilization of growth medium alleviates inhibition of Aggregatibacter actinomycetemcomitans by Maillard reaction products. J. Microbiol. Methods 2009, 78, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, M.K.; Delsol, J.K. Does microwave sterilization of growth media involve any non-thermal effect? J. Microbiol. Methods 2014, 96, 70–72. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GF-F6 (μL) | Viable Cells (108 CFU/mL) 1 | ||
|---|---|---|---|
| 0 h | 4 h | 12 h | |
| 0 | 1.39 ± 0.48 | 1.90 ± 0.15 | 1.92 ± 0.15 |
| 20 | 1.52 ± 0.42 | 0.16 ± 0.06 | 0.005 ± 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharjee, M.K.; Bommareddy, P.K.; DePass, A.L. A Water-Soluble Antibiotic in Rhubarb Stalk Shows an Unusual Pattern of Multiple Zones of Inhibition and Preferentially Kills Slow-Growing Bacteria. Antibiotics 2021, 10, 951. https://doi.org/10.3390/antibiotics10080951
Bhattacharjee MK, Bommareddy PK, DePass AL. A Water-Soluble Antibiotic in Rhubarb Stalk Shows an Unusual Pattern of Multiple Zones of Inhibition and Preferentially Kills Slow-Growing Bacteria. Antibiotics. 2021; 10(8):951. https://doi.org/10.3390/antibiotics10080951
Chicago/Turabian StyleBhattacharjee, Mrinal K., Praveen K. Bommareddy, and Anthony L. DePass. 2021. "A Water-Soluble Antibiotic in Rhubarb Stalk Shows an Unusual Pattern of Multiple Zones of Inhibition and Preferentially Kills Slow-Growing Bacteria" Antibiotics 10, no. 8: 951. https://doi.org/10.3390/antibiotics10080951
APA StyleBhattacharjee, M. K., Bommareddy, P. K., & DePass, A. L. (2021). A Water-Soluble Antibiotic in Rhubarb Stalk Shows an Unusual Pattern of Multiple Zones of Inhibition and Preferentially Kills Slow-Growing Bacteria. Antibiotics, 10(8), 951. https://doi.org/10.3390/antibiotics10080951