
Colistin Resistance and ESBL Production in Salmonella and Escherichia coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Salmonella Prevalence and Serovars in Pigs and Pork
2.2. E. coli Prevalence in Pork
2.3. ESBL Production and Colistin Resistance in Salmonella and E. coli
2.4. Prevalence of ESBL-Producing Salmonella and E. coli
2.5. Genotype of Colistin-Resistant and ESBL-Producing Salmonella and E. coli
3. Discussion
4. Materials and Methods
4.1. Sampling Location and Sample Collection
4.2. Isolation and Identification of Salmonella and E. coli
4.3. Determination of Minimum Inhibitory Concentration (MICs) of Colistin
4.4. Determination of ESBL Production
4.5. PCR and Nucleotide Sequencing
4.6. Detection of ESBL and mcr Genes
4.7. Test of Conjugation Transfer of ESBL Genes
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Humans as the world’s greatest evolutionary force. Science 2001, 293, 1786–1790. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, L.M.; Abatih, E.; Aarestrup, F.M.; Guardabassi, L. Selection and persistence of CTX-M-producing Escherichia coli in the intestinal flora of pigs treated with amoxicillin, ceftiofur, or cefquinome. Antimicrob. Agents Chemother. 2008, 52, 3612–3616. [Google Scholar] [CrossRef] [PubMed]
- WHO; WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR). Critically Important Antimicrobials for Human Medicine, 3rd ed.; WHO: Geneva, Switzerland, 2012; Available online: https://www.alimenti-salute.it/sites/default/files/CIA’s%20human_WHO_2011_eng.pdf (accessed on 24 March 2021).
- OIE. OIE List of Antimicrobials of Veterinary Importance 2007. Resolution No. XXVIII, General Session.: 1–7. WHO 2007. Available online: https://www.oie.int/fileadmin/Home/eng/Internationa_Standard_Setting/docs/pdf/OIE_list_antimicrobials.pdf (accessed on 24 December 2019).
- Panta, K.; Ghimire, P.; Rai, S.K.; Mukhiya, R.K.; Singh, R.N.; Rai, G. Antibiogram typing of gram negative isolates in different clinical samples of a tertiary hospital. Asian J. Pharm. Clin. Res. 2013, 6, 153–156. [Google Scholar]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef]
- Geser, N.; Stephan, R.; Hächler, H. Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 2012, 8, 21. [Google Scholar] [CrossRef]
- Ben Sallem, R.; Ben Slama, K.; Sáenz, Y.; Rojo-Bezares, B.; Estepa, V.; Jouini, A.; Gharsa, H.; Klibi, N.; Boudabous, A.; Torres, C. Prevalence and characterization of extended-spectrum beta-lactamase (ESBL)–and CMY-2–producing Escherichia coli isolates from healthy food-producing animals in Tunisia. Foodborne Pathog. Dis. 2012, 9, 1137–1142. [Google Scholar] [CrossRef]
- Sihem, M.; Salah, H.; Riadh, M.; Salah, A.M. Overview of ESBL-producing Escherichia coli of Animal Origin in Tunisia: In the Way of the Global Spread of CTX-M β-Lactamases. Arch. Clin. Microbiol. 2015, 6, 1–7. [Google Scholar]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Navas, A.L.; Mackay, D.; et al. Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Butaye, P.; Wang, C. Colistin resistance, beyond the current knowledge. EBioMedicine 2018, 34, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Feng, Y.; Ma, K.; Liu, L.; McNally, A.; Zong, Z. The co-transfer of plasmid-borne colistin-resistant genes mcr-1 and mcr-3.5, the carbapenemase gene blaNDM-5 and the 16S methylase gene rmtB from Escherichia coli. Sci. Rep. 2019, 9, 696. [Google Scholar] [CrossRef] [PubMed]
- Manarungsan, S. Thailand’s Trade Relations with GMS and CLMV. In Japan and Korea with Mekong River Basin Countries; Kagami, M., Ed.; BRV Research Report No. 3; Bangkok Research Center, IDE-JETRO: Bangkok, Thailand, 2010; Available online: https://www.ide.go.jp/library/English/Publish/Download/Brc/pdf/03_chapter6.pdf (accessed on 24 March 2021).
- Trongjit, S.; Angkittitrakul, S.; Chuanchuen, R. Occurrence and molecular characteristics of antimicrobial resistance of Escherichia coli from broilers, pigs and meat products in Thailand and Cambodia provinces. Microbiol. Immunol. 2016, 60, 575–585. [Google Scholar] [CrossRef] [PubMed]
- DLD. Control Salmonella in Poultry. Notification of the Department of Livestock Development. The Ministry of Agriculture and Cooperatives, Thailand. 2010. Available online: http://www.ratchakitcha.soc.go.th (accessed on 20 May 2021).
- BQSF. Microbiological Quality Criteria for Food and Food Contact Containers No. 3. Notification of the Bureau of Quality and Safety of Food, Department of Medical Sciences, The Ministry of Public Health, Thailand. 2017. Available online: http://bqsf.dmsc.moph.go.th (accessed on 20 May 2021).
- Pornsukarom, S.; Patchanee, P.; Erdman, M.; Cray, P.F.; Wittum, T.; Lee, J.; Gebreyes, W.A. Comparative phenotypic and genotypic analyses of Salmonella Rissen that originated from food animals in Thailand and United States. Zoonoses Public Health. 2015, 62, 151–158. [Google Scholar] [CrossRef]
- Boonyasiri, A.; Tangkoskul, T.; Seenama, C.; Saiyarin, J.; Tiengrim, S.; Thamlikitkul, V. Prevalence of antibiotic resistant bacteria in healthy adults, foods, food animals, and the environment in selected areas in Thailand. Pathog. Glob. Health 2014, 108, 235–245. [Google Scholar] [CrossRef]
- Nuangmek, A.; Rojanasthien, S.; Chotinun, S.; Yamsakul, P.; Tadee, P.; Thamlikitkul, V.; Tansakul, N.; Patchanee, P. Antimicrobial Resistance in ESBL-Producing Escherichia coli Isolated from Layer and Pig Farms in Thailand. Acta. Sci. Vet. 2018, 46, 8. Available online: https://seer.ufrgs.br/ActaScientiaeVeterinariae/article/view/81823 (accessed on 15 April 2021). [CrossRef]
- Hammerum, A.M.; Larsen, J.; Andersen, V.D.; Lester, C.H.; Skytte, T.S.S.; Hansen, F.; Olsen, S.S.; Mordhorst, H.; Skov, R.L.; Aarestrup, F.M.; et al. Characterization of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third-or fourth-generation cephalosporins. J. Antimicrob. Chemother. 2014, 69, 2650–2657. [Google Scholar] [CrossRef]
- Zelendova, M.; Dolejska, M.; Masarikova, M.; Jamborova, I.; Vasek, J.; Smola, J.; Manga, I.; Cizek, A. CTX-M-producing Escherichia coli in pigs from a Czech farm during production cycle. Lett. Appl. Microbiol. 2020, 71, 369–376. [Google Scholar] [CrossRef]
- Olowe, O.A.; Adewumi, O.; Odewale, G.; Ojurongbe, O.; Adefioye, O.J. Phenotypic and molecular characterisation of extended-spectrum beta-lactamase producing Escherichia coli obtained from animal fecal samples in Ado Ekiti, Nigeria. J. Environ. Public Health 2015, 2015, 497980. [Google Scholar] [CrossRef]
- Schill, F.; Abdulmawjood, A.; Klein, G.; Reich, F. Prevalence and characterization of extended-spectrum β-lactamase (ESBL) and AmpC β-lactamase producing Enterobacteriaceae in fresh pork meat at processing level in Germany. Int. J. Food Microbiol. 2017, 257, 58–66. [Google Scholar] [CrossRef]
- Trongjit, S.; Angkititrakul, S.; Tuttle, R.E.; Poungseree, J.; Padungtod, P.; Chuanchuen, R. Prevalence and antimicrobial resistance in Salmonella enterica isolated from broiler chickens, pigs and meat products in Thailand–Cambodia border provinces. Microbiol. Immunol. 2017, 61, 23–33. [Google Scholar] [CrossRef]
- Guiral, E.; Pons, M.J.; Vubil, D.; Marí-Almirall, M.; Sigaúque, B.; Soto, S.M.; Alonso, P.L.; Ruiz, J.; Vila, J.; Mandomando, I. Epidemiology and molecular characterization of multidrug-resistant Escherichia coli isolates harboring blaCTX-M group 1 extended-spectrum β-lactamases causing bacteremia and urinary tract infection in Manhiça, Mozambique. Infect. Drug Resist. 2018, 11, 927–936. [Google Scholar] [CrossRef]
- Coyne, L.; Arief, R.; Benigno, C.; Giang, V.N.; Huong, L.Q.; Jeamsripong, S.; Kalpravidh, W.; McGrane, J.; Padungtod, P.; Patrick, I.; et al. Characterizing Antimicrobial Use in the Livestock Sector in Three South East Asian Countries (Indonesia, Thailand, and Vietnam). Antibiotics 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Nam, H.M.; Kim, S.R.; Chae, M.H.; Jang, G.C.; Jung, S.C.; Lim, S.K. Prevalence and molecular characterization of CTX-M β-lactamase–producing Escherichia coli isolated from healthy swine and cattle. Foodborne Pathog. Dis. 2013, 10, 13–20. [Google Scholar] [CrossRef]
- Tong, P.; Sun, Y.; Ji, X.; Du, X.; Guo, X.; Liu, J.; Zhu, L.; Zhou, B.; Zhou, W.; Liu, G.; et al. Characterization of antimicrobial resistance and extended-spectrum β-lactamase genes in Escherichia coli isolated from chickens. Foodborne Pathog. Dis. 2015, 12, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.P.; Lemma, F.; Rogers, J.P.; Cheney, T.E.A.; Powell, L.F.; Teale, C.J. Prevalence of extended-spectrum-β-lactamase-producing Escherichia coli from pigs at slaughter in the UK in 2013. J. Antimicrob. Chemother. 2014, 69, 2947–2950. [Google Scholar] [CrossRef][Green Version]
- Valentin, L.; Sharp, H.; Hille, K.; Seibt, U.; Fischer, J.; Pfeifer, Y.; Michael, G.B.; Nickel, S.; Schmiedel, J.; Falgenhauer, L.; et al. Subgrouping of ESBL-producing Escherichia coli from animal and human sources: An approach to quantify the distribution of ESBL types between different reservoirs. Inter. J. Med. Microbiol. 2014, 304, 805–816. [Google Scholar] [CrossRef]
- Zheng, H.; Zeng, Z.; Chen, S.; Liu, Y.; Yao, Q.; Deng, Y.; Chen, X.; Lv, L.; Zhuo, C.; Chen, Z.; et al. Prevalence and characterisation of CTX-M β-lactamases amongst Escherichia coli isolates from healthy food animals in China. Int. J. Antimicrob. Agents. 2012, 39, 305–310. [Google Scholar] [CrossRef] [PubMed]
- DLD. A List of Drugs that Must not be Mixed in Animal Feed for Prophylactic Purposes. Notification of the Department of Livestock Development. The Ministry of Agriculture and Cooperatives, 2018. Available online: http://legal.dld.go.th (accessed on 20 May 2021).
- DLD. Characteristics and Conditions of Animal feed Containing Drugs Prohibited from Producing, Importing, Selling and using. Notification of the Department of Livestock Development. The Ministry of Agriculture and Cooperatives, 2019. Available online: https://legal.dld.go.th/web2012/news/P15/13136182190762.PDF (accessed on 20 May 2021).
- Poolperm, P.; Tangkoskul, T.; Seenama, C.; Maknakhon, N.; Thamlikitkul, V. Association between the use of colistin for short-term treatment of Gram-negative bacterial infections and the emergence of colistin-resistant Enterobacteriaceae in swine from selected swine farms in Thailand. PLoS ONE 2020, 15, e0238939. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Wang, J.; Yassin, A.K.; Butaye, P.; Kelly, P.; Gong, J.; Guo, W.; Li, J.; Li, M.; et al. Molecular detection of colistin resistance genes (mcr-1, mcr-2 and mcr-3) in nasal/oropharyngeal and anal/cloacal swabs from pigs and poultry. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Eurosurveillance 2016, 21, 30280. [Google Scholar] [CrossRef]
- Falgenhauer, L.; Waezsada, S.E.; Yao, Y.; Imirzalioglu, C.; Käsbohrer, A.; Roesler, U.; Michael, G.B.; Schwarz, S.; Werner, G.; Kreienbrock, L.; et al. Colistin resistance gene mcr-1 in extended-spectrum β-lactamase-producing and carbapenemase-producing Gram-negative bacteria in Germany. Lancet Infect. Dis. 2016, 16, 282–283. [Google Scholar] [CrossRef]
- Yao, X.; Doi, Y.; Zeng, L.; Luchao, L.V.; Liu, J.H. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef]
- International Organization for Standardization (ISO). Microbiology of Food and Animal Feeding Stuffs–Horizontal Method for the Detection of Salmonella spp.; ISO 6579:2002; ISO: Rome, Italy, 2002. [Google Scholar]
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; World Health Organization Collaborating Center for Reference and Research on Salmonella; Institut Pasteur: Paris, France, 2007; pp. 1–166. [Google Scholar]
- Quinn, P.J. Clinical Veterinary Microbiology; Wolfe: London, UK, 1994; p. 648. [Google Scholar]
- Carter, G.R.; Cole, J.R., Jr. Diagnostic Procedure in Veterinary Bacteriology and Mycology; Academic Press: Cambridge, MA, USA, 2012; p. 620. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-third Informational Supplement M100-S25; Clinical Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015. [Google Scholar]
- EUCAST. European Committee on Antimicrobial Susceptibility Testing, Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0. In European Committee on Antimicrobial Susceptibility Testing; 2020; Available online: http://www.eucast.org/clinical_breakpoints/ (accessed on 25 May 2021).
- Levesque, C.; Piche, L.; Larose, C.; Roy, P.H. PCR mapping of integrons reveals several novel combinations of resistance genes. J. Antimicrob. Chemother. 1995, 39, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio 2017, 8, e00543. [Google Scholar] [CrossRef]
- Hasman, H.; Mevius, D.; Veldman, K.; Olesen, I.; Aarestrup, F.M. β-Lactamases among extended-spectrum β-lactamase (ESBL)-resistant Salmonella from poultry, poultry products and human patients in The Netherlands. J. Antimicrob. Chemother. 2005, 56, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Batchelor, M.; Hopkins, K.; Threlfall, E.J.; Clifton-Hadley, F.A.; Stallwood, A.D.; Davies, R.H.; Liebana, E. blaCTX-M genes in clinical Salmonella isolates recovered from humans in England and Wales from 1992 to 2003. Antimicrob. Agents Chemother. 2005, 49, 1319–1322. [Google Scholar] [CrossRef]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Sabaté, M.; Tarragó, R.; Navarro, F.; Miró, E.; Vergés, C.; Barbé, J.; Prats, G. Cloning and Sequence of the Gene Encoding a Novel Cefotaxime-Hydrolyzing β-Lactamase (CTX-M-9) from Escherichia coli in Spain. Antimicrob. Agents Chemother. 2000, 44, 1970–1973. [Google Scholar] [CrossRef]
- Muzaheed, Y.D.; Adams-Haduch, J.M.; Endimiani, A.; Sidjabat, H.E.; Gaddad, S.M.; Paterson, D.L. High prevalence of CTX-M-15-producing Klebsiella pneumoniae among inpatients and outpatients with urinary tract infection in Southern India. J. Antimicrob. Chemother. 2008, 61, 1393–1394. [Google Scholar] [CrossRef] [PubMed]
- Woodall, C.A. DNA transfer by bacterial conjugation. In E. coli Plasmid Vectors; Nicola, C., Andrew, P., Eds.; Humana Press: Totowa, NJ, USA, 2003; pp. 61–65. [Google Scholar]
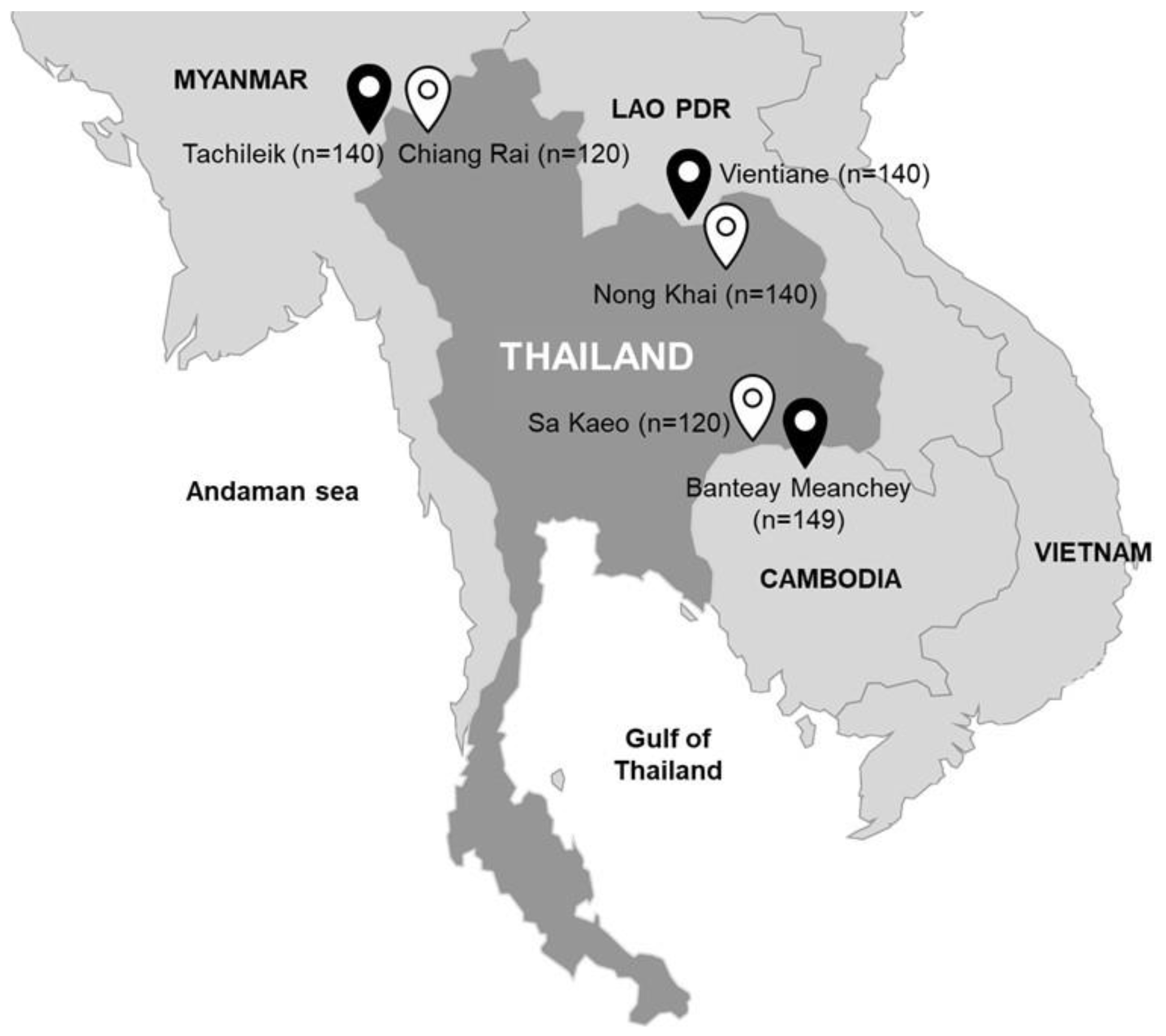
 ) and other neighboring counties (
) and other neighboring counties ( ) including Lao PDR, Cambodia and Myanmar. Number of samples is indicated in parenthesis.
) and other neighboring counties () including Lao PDR, Cambodia and Myanmar. Number of samples is indicated in parenthesis.
) including Lao PDR, Cambodia and Myanmar. Number of samples is indicated in parenthesis.
) and other neighboring counties () including Lao PDR, Cambodia and Myanmar. Number of samples is indicated in parenthesis.

{kind=link}
| Country | Location | Source | Type of Sample | No. of Sample | No. of Positive Sample (%) | |
|---|---|---|---|---|---|---|
| Salmonella | E. coli 1 | |||||
| Thailand | Nong Khai | Slaughterhouse | Rectal swab | 80 | 57 (71.3) | 79 (98.8) |
| Market | Carcass swab | 60 | 46 (76.7) | 57 (95.0) | ||
| Subtotal | 140 | 103 (73.6) | - | |||
| Sa Kaeo | Slaughterhouse | Rectal swab | 60 | 2 (3.3) | 58 (96.7) | |
| Market | Carcass swab | 60 | 34 (56.7) | 58 (96.7) | ||
| Subtotal | 120 | 36 (30.0) | - | |||
| Chiangrai | Slaughterhouse | Rectal swab | 60 | 30 (50.0) | 58 (96.7) | |
| Market | Carcass swab | 60 | 45 (75.0) | 58 (96.7) | ||
| Subtotal | 120 | 75 (62.5) | - | |||
| Cambodia | Banteay Meanchey | Slaughterhouse | Rectal swab | 84 | 39 (46.4) | 79 (94.0) |
| Market | Carcass swab | 65 | 59 (90.8) | 57 (87.7) | ||
| Subtotal | 149 | 98 (65.8) | - | |||
| Lao PDR | Vientiane | Slaughterhouse | Rectal swab | 82 | 28 (34.1) | 79 (96.3) |
| Market | Carcass swab | 58 | 44 (75.9) | 54 (93.1) | ||
| Subtotal | 140 | 72 (51.4) | - | |||
| Myanmar | Tarchileik | Slaughterhouse | Rectal swab | 75 | 1 (1.3) | 71 (94.7) |
| Market | Carcass swab | 65 | 18 (27.7) | 59 (90.8) | ||
| Subtotal | 140 | 19 (13.6) | - | |||
| Grand total | 809 | 403 (49.8) | 343(93) 2 | |||
| Salmonella Serotype | No. of Isolates (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Thailand (n = 237) | Laos PDR (n = 86) | Cambodia (n = 121) | Myanmar (n = 19) | |||||
| Pig | Pork | Pig | Pork | Pig | Pork | Pig | Pork | |
| Typhimurium | 12 (5.0) | - | 1 (1.2) | - | 2 (1.7) | - | - | - |
| Sao | 12 (5.0) | 8 (3.3) | - | - | - | - | - | - |
| Augustenborg | 1 (0.4) | 1 (0.4) | - | 2 (2.3) | - | - | - | - |
| Schwarzengrund | 2 (0.8) | - | - | 1 (1.2) | - | - | - | - |
| Derby | 2 (0.8) | - | - | - | 3 (2.5) | 1 (0.8) | - | - |
| Rissen | 37 (15.5) | 66 (27.6) | - | 11 (12.8) | 18 (14.9) | 30 (24.8) | 1 (5.3) | 2 (10.5) |
| Saintpaul | 2 (0.8) | 8 (3.3) | - | 1 (1.2) | - | - | - | 2 (10.5) |
| Eastbourne | 1 (0.4) | - | - | - | - | - | - | - |
| Anatum | 8 (3.3) | 30 (12.6) | 5 (5.8) | 5 (5.8) | - | 9 (7.4) | - | 14 (73.7) |
| Rideau | 1 (0.4) | 4 (1.7) | - | 1 (1.2) | - | - | - | - |
| Sanktmarx | 5 (2.1) | 10 (4.2) | 1 (1.2) | 2 (2.3) | - | - | - | - |
| Weltevreden | 4 (1.7) | 1 (0.4) | - | - | 1 (0.8) | 1 (0.8) | - | - |
| Braenderup | 1 (0.4) | - | - | - | - | 2 (1.7) | - | - |
| Fareham | 4 (1.7) | 2 (0.8) | - | - | - | - | - | - |
| Stanley | 2 (0.8) | 1 (0.4) | 8 (9.3) | 7 (8.1) | 3 (2.5) | 5 (4.1) | - | - |
| Vijle-1 | - | 1 (0.4) | - | - | - | - | - | - |
| Norwich | - | 2 (0.8) | - | 1 (1.2) | - | - | - | - |
| Yalding | - | 4 (1.7) | - | - | - | - | - | - |
| Calabar | - | 1 (0.4) | - | - | - | - | - | - |
| Hayindogo | - | 3 (1.3) | - | 6 (7.0) | - | 1 (0.8) | - | - |
| Muenster | - | 1 (0.4) | - | 3 (3.5) | 1 (0.8) | 5 (4.1) | - | - |
| Potto | - | - | 3 (3.5) | - | - | - | - | - |
| Tsevie | - | - | 1 (1.2) | - | 2 (1.7) | - | - | - |
| Brunei | - | - | 4 (4.7) | - | - | - | - | - |
| Kissi | - | - | 1 (1.2) | - | - | - | - | - |
| Eschberg | - | - | 1 (1.2) | - | - | - | - | - |
| Ayinde | - | - | 1 (1.2) | - | - | - | - | - |
| Kentucky | - | - | 1 (1.2) | 2 (2.3) | - | - | - | - |
| Rottnest | - | - | 1 (1.2) | - | - | - | - | - |
| Vilvoorde | - | - | 1 (1.2) | - | - | - | - | - |
| Kouka | - | - | 1 (1.2) | - | - | - | - | - |
| Portanigra | - | - | - | 1 (1.2) | - | - | - | - |
| Newlands | - | - | - | 2 (2.3) | - | - | - | - |
| Bristol | - | - | - | 1 (1.2) | - | - | - | - |
| Sandow | - | - | - | 1 (1.2) | - | 10 (8.3) | - | - |
| Haifa | - | - | - | 1 (1.2) | - | 1 (0.8) | - | - |
| Magumeri | - | - | - | 1 (1.2) | - | - | - | - |
| Lika | - | - | - | 1 (1.2) | - | - | - | - |
| V | - | - | - | 1 (1.2) | - | - | - | - |
| Koenigstuhl | - | - | - | 1 (1.2) | 1 (0.8) | - | - | - |
| II | - | - | - | 1 (1.2) | - | 1 (0.8) | - | - |
| Suberu | - | - | - | 1 (1.2) | - | - | - | - |
| Ikayi | - | - | - | 2 (2.3) | - | - | - | - |
| Dallgow | - | - | - | - | 1 (0.8) | - | - | - |
| Paratyphi-B | - | - | - | - | 1 (0.8) | - | - | - |
| Lekke | - | - | - | - | 1 (0.8) | - | - | - |
| Herston | - | - | - | - | 1 (0.8) | - | - | - |
| Hvittingfoss/II | - | - | - | - | 1 (0.8) | - | - | - |
| Stanley ville | - | - | - | - | 1 (0.8) | - | - | - |
| Bradford | - | - | - | - | 1 (0.8) | - | - | - |
| Yoruba | - | - | - | - | 1 (0.8) | - | - | - |
| Rechovot | - | - | - | - | 4 (3.3) | - | - | - |
| Bracknell | - | - | - | - | 1 (0.8) | - | - | - |
| Idikan | - | - | - | - | 1 (0.8) | - | - | - |
| Sinstorf | - | - | - | - | - | 5 (4.1) | - | - |
| Paris | - | - | - | - | - | 1 (0.8) | - | - |
| Newport | - | - | - | - | - | 1 (0.8) | - | - |
| Ituri | - | - | - | - | - | 1 (0.8) | - | - |
| Kedougou | - | - | - | - | - | 1 (0.8) | - | - |
| Havana | - | - | - | - | - | 1 (0.8) | - | - |
| Total | 94 (39.7) | 143 (60.3) | 30 (34.9) | 56 (65.1) | 45 (37.2) | 76 (62.8) | 1 (5.3) | 18 (94.7) |
| Country | Source | Total No. | Salmonella | Source | Total No. | E. coli | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ESBL Production, No. (%) 1 | Colistin Resistance, No. (%) 2 | ESBL Production, No. (%) 1 | Colistin Resistance, No. (%) 2 | ||||||||||||||
| Total | blaCTX-M-55 | blaTEM-1 | Total | mcr-1 | mcr-3 | Total | blaCTX-M-55 | blaCTX-M-14 | blaTEM-1 | Total | mcr-1 | mcr-3 | |||||
| Thailand | Pig | 94 | 2 (2.1) | 1 (1.1) | 1 (1.1) | - | - | 1 (1.1) | Pig | 195 | 12 (6.2) | 8 (4.1) | 4 (2.1) | 7 (3.6) | 8 (4.1) | 11 (5.6) | 5 (2.6) |
| Pork | 143 | 6 (4.2) | 6 (4.2) | 6 (4.2) | 6 (4.2) | 6 (4.2) | - | Pork | 173 | 9 (5.2) | 6 (3.5) | 4 (2.3) | 6 (3.5) | 14 (8.1) | 6 (3.5) | 4 (2.3) | |
| Total | 237 | 8 (3.4) | 7 (3.0) | 7 (3.0) | 6 (2.5) | 6 (2.5) | 1 (0.4) | Total | 368 | 21 (5.7) | 14 (3.8) | 8 (2.2) | 13 (3.5) | 22 (6.0) | 17 (4.6) | 9 (2.4) | |
| Cambodia | Pig | 45 | - | - | - | - | - | - | Pig | 79 | 2 (2.5) | 2 (2.5) | - | 2 (2.5) | 11 (13.9) | 9 (11.4) | 5 (6.3) |
| Pork | 76 | - | - | - | 4 (5.3) | 4 (5.3) | Pork | 57 | 1 (1.8) | 1 (1.8) | - | - | 7 (12.3) | 4 (7.0) | 5 (8.8) | ||
| Total | 121 | - | - | - | 4 (3.3) | 4 (3.3) | Total | 136 | 3 (2.2) | 3 (2.2) | - | 2 (1.5) | 18 (13.2) | 13 (9.6) | 10 (7.4) | ||
| Lao PDR | Pig | 30 | - | - | - | - | - | - | Pig | 79 | 6 (7.6) | 2 (2.5) | 4 (5.1) | 5 (6.3) | 18 (22.8) | 16 (20.3) | 6 (7.6) |
| Pork | 56 | 1 (1.8) | 1 (1.8) | - | 1 (1.8) | 1 (1.8) | - | Pork | 54 | - | 1 (1.9) | - | - | 20 (37.0) | 17 (31.5) | 6 (11.1) | |
| Total | 86 | 1 (1.2) | 1 (1.2) | - | 1 (1.2) | 1 (1.2) | - | Total | 133 | 6 (4.5) | 3 (2.6) | 4 (3.0) | 5 (3.8) | 38 (28.6) | 33 (24.8) | 12 (9.0) | |
| Myanmar | Pig | 1 | - | - | - | - | - | - | Pig | 71 | 13 (18.3) | 10 (14.1) | 3 (4.2) | 5 (7.0) | - | 2 (2.8) | - |
| Pork | 18 | - | - | - | 1 (5.6) | 1 (5.6) | - | Pork | 59 | 5 (8.5) | 4 (6.8) | 1 (1.7) | 1 (1.7) | 2 (3.4) | 3 (5.1) | - | |
| Total | 19 | - | - | - | 1 (5.6) | 1 (5.3) | - | Total | 130 | 18 (13.8) | 14 (10.8) | 4 (3.1) | 6 (4.6) | 2 (1.5) | 5 (3.8) | - | |
| Grand total | 463 | 9 (1.9) | 8 (1.7) | 7 (1.5) | 13 (2.8) | 12 (2.6) | 1 (0.2) | Grand total | 767 | 48 (6.3) | 34 (4.4) | 16 (2.1) | 26 (3.4) | 80 (10.4) | 68 (8.9) | 31 (4.0) | |
| Country | Source | Salmonella | E. coli | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total No. | No. of Positive (%) | Total No. | No. of Positive (%) | ||||||
| Ceftazidime | Cefotaxime | Cefpodoxime | Ceftazidime | Cefotaxime | Cefpodoxime | ||||
| Thailand | Pigs | 94 | 3 (3.2) | 2 (2.1) | 3 (3.2) | 195 | 12 (6.2) | 19 (9.7) | 19 (9.7) |
| Pork | 143 | 5 (3.5) | 6 (4.2) | 6 (4.2) | 173 | 14 (8.1) | 15 (8.7) | 16 (9.2) | |
| Total | 237 | 8 (3.4) | 8 (3.4) | 9 (3.8) | 368 | 26 (7.1) | 34 (9.2) | 35 (9.5) | |
| Cambodia | Pigs | 45 | 1 (2.2) | 1 (2.2) | 1 (2.2) | 79 | 3 (3.8) | 4 (5.1) | 2 (2.5) |
| Pork | 76 | - | - | - | 57 | 4 (7.0) | 1 (1.8) | 1 (1.8) | |
| Total | 121 | 1 (0.8) | 1 (0.8) | 1 (0.8) | 136 | 7 (5.1) | 5 (3.7) | 3 (2.2) | |
| Lao PDR | Pigs | 30 | 1 (3.3) | 1 (3.3) | 1 (3.3) | 79 | 3 (3.8) | 8 (10.1) | 7 (8.9) |
| Pork | 56 | 1 (1.8) | 1 (1.8) | 1 (1.8) | 54 | 1 (1.9) | 1 (1.9) | 1 (1.9) | |
| Total | 86 | 2 (2.3) | 2 (2.3) | 2 (2.3) | 133 | 4 (3.0) | 9 (6.8) | 8 (6.0) | |
| Myanmar | Pigs | 1 | - | - | - | 71 | 7 (9.9) | 13 (18.3) | 13 (18.3) |
| Pork | 18 | 1 (5.6) | - | - | 59 | 5 (8.5) | 8 (13.6) | 8 (13.6) | |
| Total | 19 | 1 (5.6) | - | - | 130 | 12 (9.2) | 21 (16.2) | 21 (16.2) | |
| Grand total | 463 | 12 (2.6) | 11 (2.4) | 12 (2.6) | 767 | 49 (6.4) | 69 (9.0) | 67 (8.7) | |
| Gene | Primer | Primer Sequence (5′-3′) | Tm (°C) | Reference |
|---|---|---|---|---|
| MCR | ||||
| mcr-1 | CLR-5F CLR-5R | CGGTCAGTCCGTTTGTTC CTTGGTCGGTCTGTA | 58 | [13] |
| mcr-2 | MCR2-IF MCR2-IR | TGTTGCTTGTGCCGATTGGA AGATGGTATTGTTGGTTGCTG | 58 | [39] |
| mcr-3 | MCR3-IF MCR3-IR | AAATAAAAATTGTTCCGCTTAT GAATGGAGATCCCCGTTTTT | 58 | [49] |
| ESBL | ||||
| blaCTX-M | CTX-M up CTX-M down | ATGTGCAGYACCAGTAARGTKATGGC TGGGTRAARTARGTSACCAGAAYCAGCGG | 60 | [50] |
| blaTEM | TEM up TEM down | GCGGAACCCCTATTTG TCTAAAGTATATATGAGTAAACTTGGTCTGAC | 50 | [51] |
| blaSHV | SHV up SHV down | TTCGCCTGTGTATTATCTCCCTG TTAGCGTTGCCAGTGYTCG | 50 | [51] |
| blaCMY-1 | CMY1 up CMY1 down | GTGGTGGATGCCAGCATCC GGTCGAGCCGGTCTTGTTGAA | 58 | [51] |
| blaCMY-2 | CMY2 up CMY2 down | GCACTTAGCCACCTATACGGCAG GCTTTTCAAGAATGCGCCAGG | 58 | [51] |
| blaPSE | PSE up PSE down | GCTCGTATAGGTGTTTCCGTTT CGATCCGCCGATGTTCCATCC | 55 | [50] |
| blaCTX-M1 | MultiCTXMGp1-F MultiCTXMGp1-R | TTAGGAARTGTGCCGCTGYA CGATATCGTTGGTGGTRCCAT | 688 | [52] |
| blaCTX-M2 | MultiCTXMGp2-F MultiCTXMGp2-R | CGTTAACGGCACGATGAC CGATATCGTTGGTGGTRCCAT | 404 | [52] |
| blaCTX-M8/25 | MultiCTXMGp8/25-F MultiCTXMGp8/25-R | AACRCRCAGACGCTCTAC TCGAGCCGGAASGTGTYAT | 326 | [52] |
| blaCTX-M9 | MultiCTXMGp9-F MultiCTXMGp9-R | GTGACAAAGAGAGTGCAACGG ATGATTCTCGCCGCTGAAGCC | 850 | [53] |
| blaCTX-M15 | CTX-M-15-SF CTX-M-15-SR | CACACGTGGAATTTAGGGACT GCCGTCTAAGGCGATAAACA | 996 | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lay, K.K.; Jeamsripong, S.; Sunn, K.P.; Angkititrakul, S.; Prathan, R.; Srisanga, S.; Chuanchuen, R. Colistin Resistance and ESBL Production in Salmonella and Escherichia coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area. Antibiotics 2021, 10, 657. https://doi.org/10.3390/antibiotics10060657
Lay KK, Jeamsripong S, Sunn KP, Angkititrakul S, Prathan R, Srisanga S, Chuanchuen R. Colistin Resistance and ESBL Production in Salmonella and Escherichia coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area. Antibiotics. 2021; 10(6):657. https://doi.org/10.3390/antibiotics10060657
Chicago/Turabian StyleLay, Khin Khin, Saharuetai Jeamsripong, Kyaw Phyoe Sunn, Sunpetch Angkititrakul, Ransiya Prathan, Songsak Srisanga, and Rungtip Chuanchuen. 2021. "Colistin Resistance and ESBL Production in Salmonella and Escherichia coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area" Antibiotics 10, no. 6: 657. https://doi.org/10.3390/antibiotics10060657
APA StyleLay, K. K., Jeamsripong, S., Sunn, K. P., Angkititrakul, S., Prathan, R., Srisanga, S., & Chuanchuen, R. (2021). Colistin Resistance and ESBL Production in Salmonella and Escherichia coli from Pigs and Pork in the Thailand, Cambodia, Lao PDR, and Myanmar Border Area. Antibiotics, 10(6), 657. https://doi.org/10.3390/antibiotics10060657