
Intrauterine Blood Plasma Platelet-Therapy Mitigates Persistent Breeding-Induced Endometritis, Reduces Uterine Infections, and Improves Embryo Recovery in Mares

 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Screening Mares for Susceptibility to PBIE
2.2. Platelet-Rich or -Poor Plasma
2.3. Semen Parameters
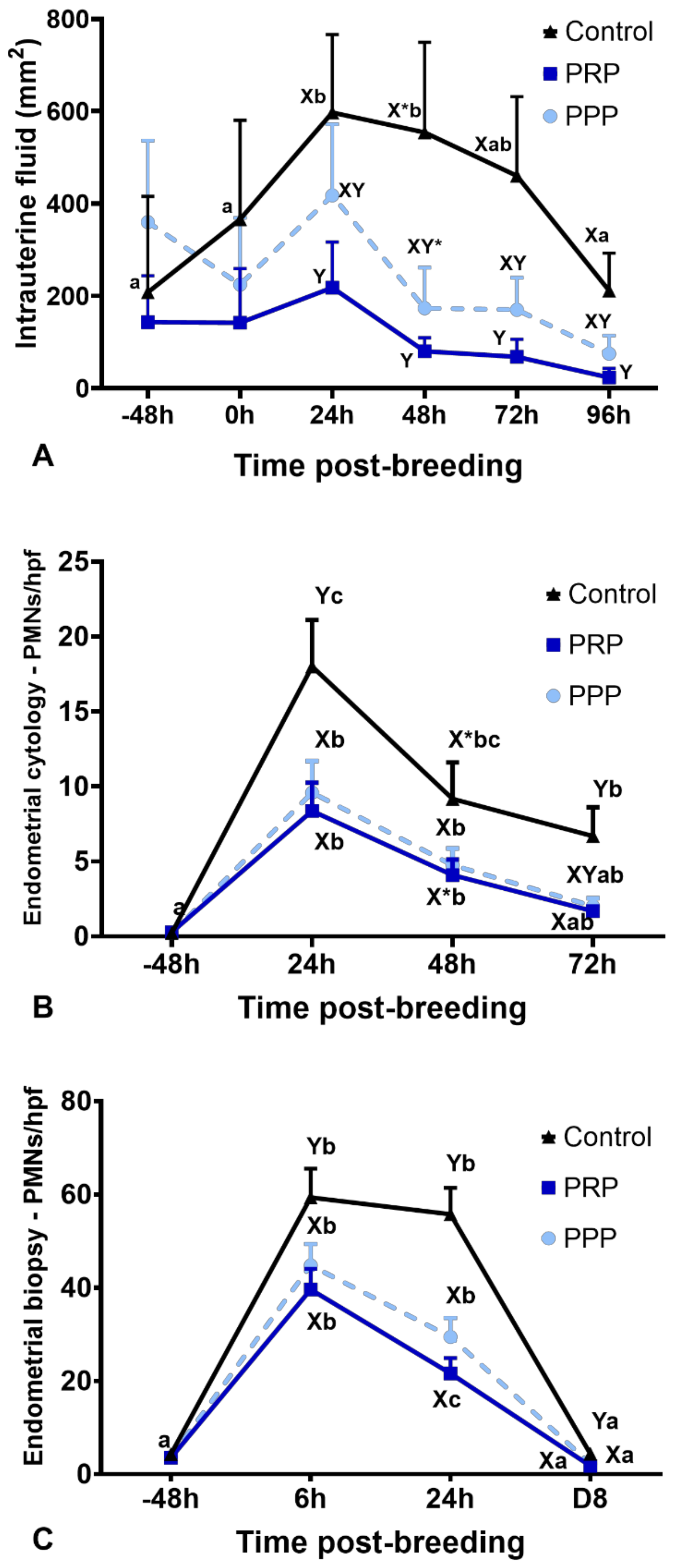
2.4. Intrauterine Fluid Accumulation and Endometrial Edema
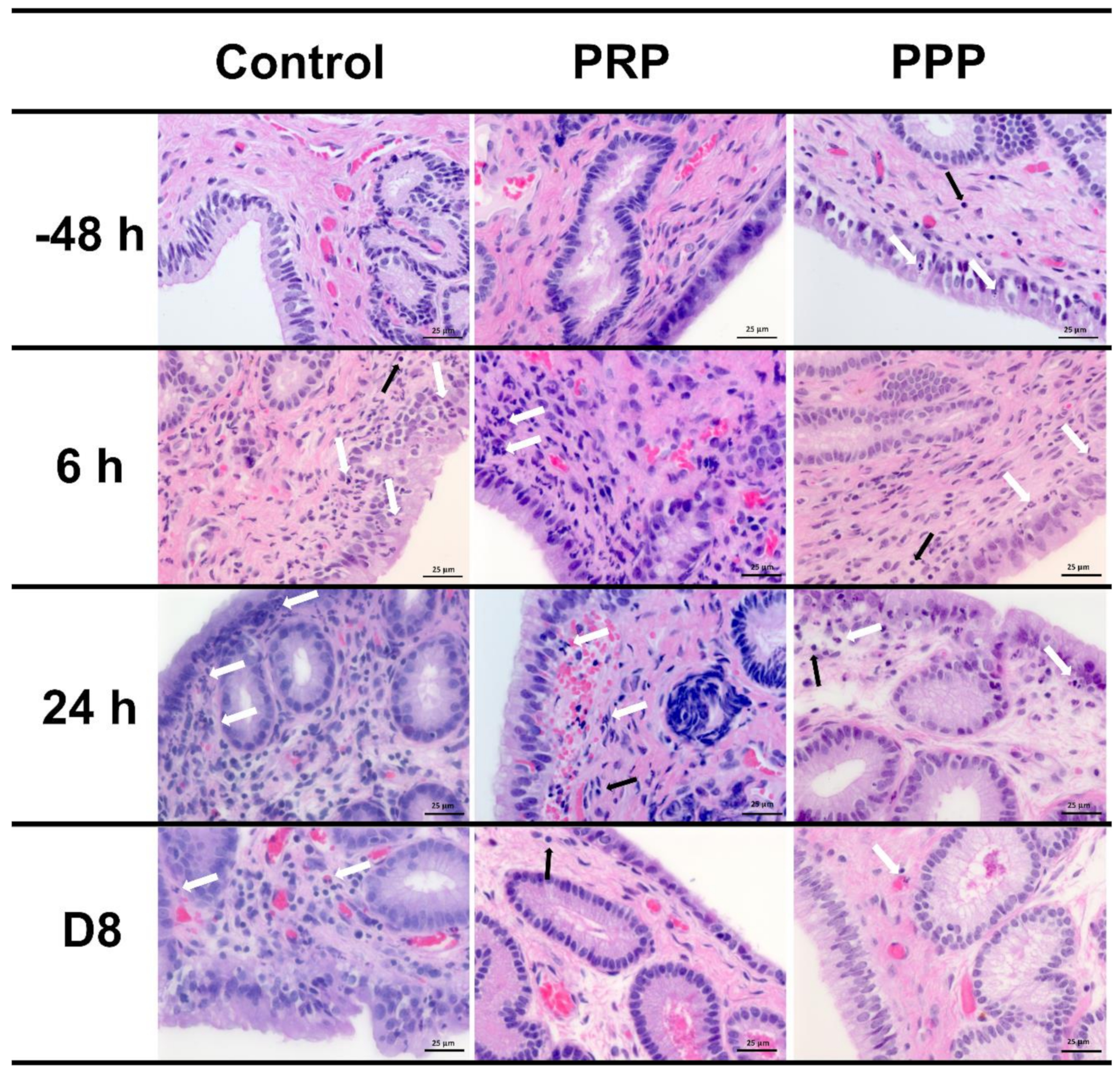
2.5. Inflammatory Cell Counts on Endometrial Biopsy and Cytology
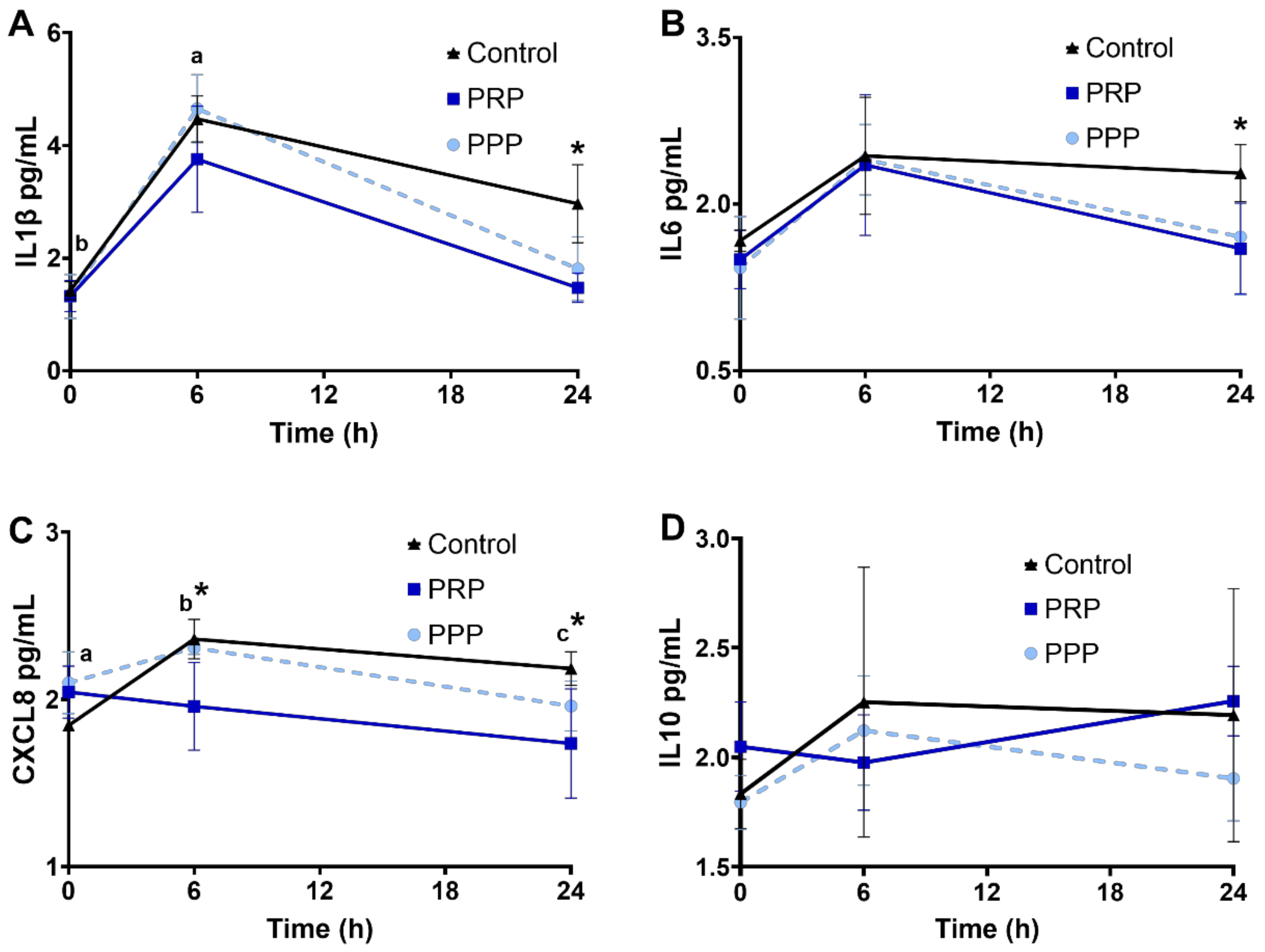
2.6. Cytokine Concentrations in Uterine Fluid
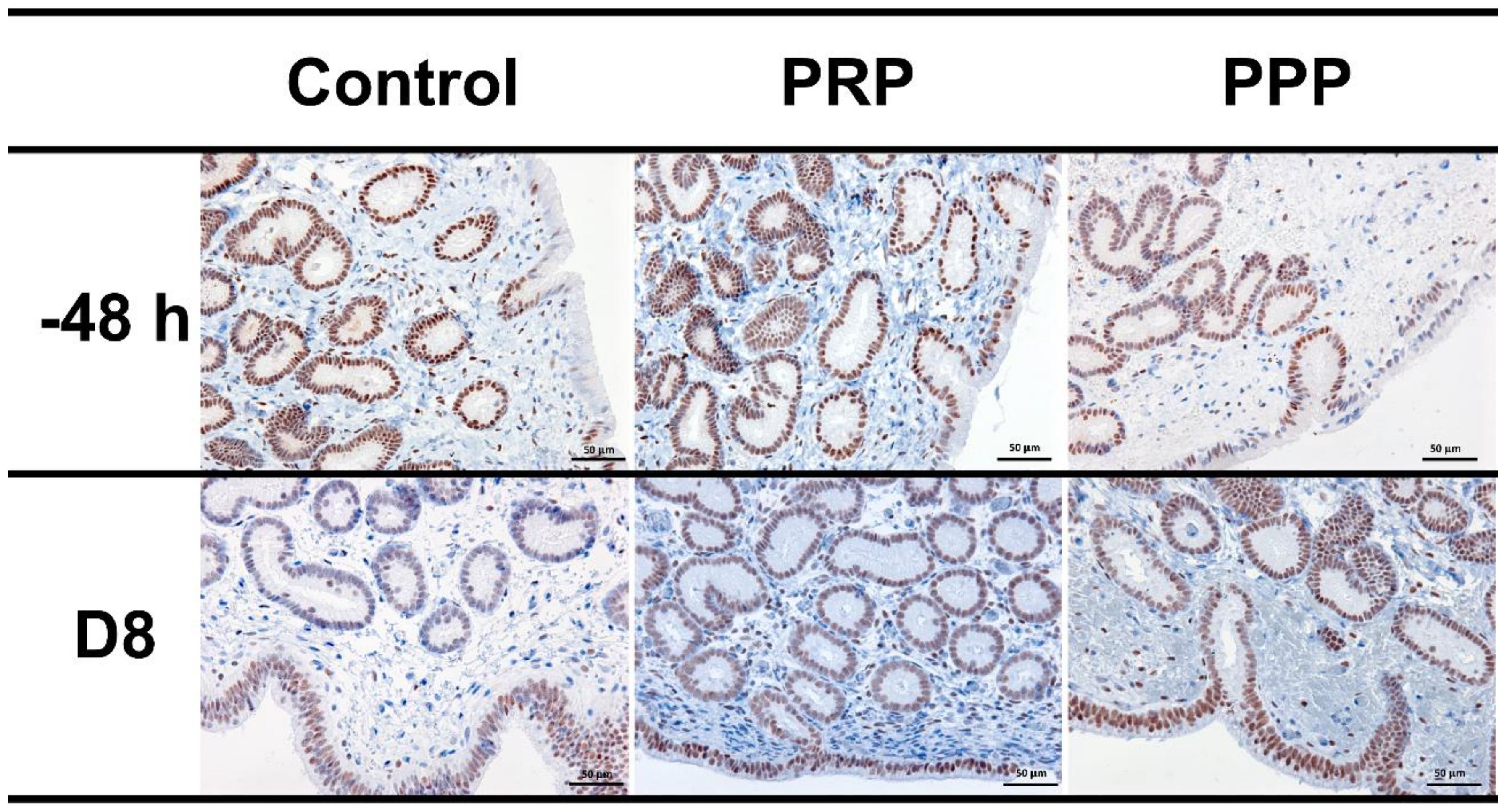
2.7. Immunohistochemical Evaluation
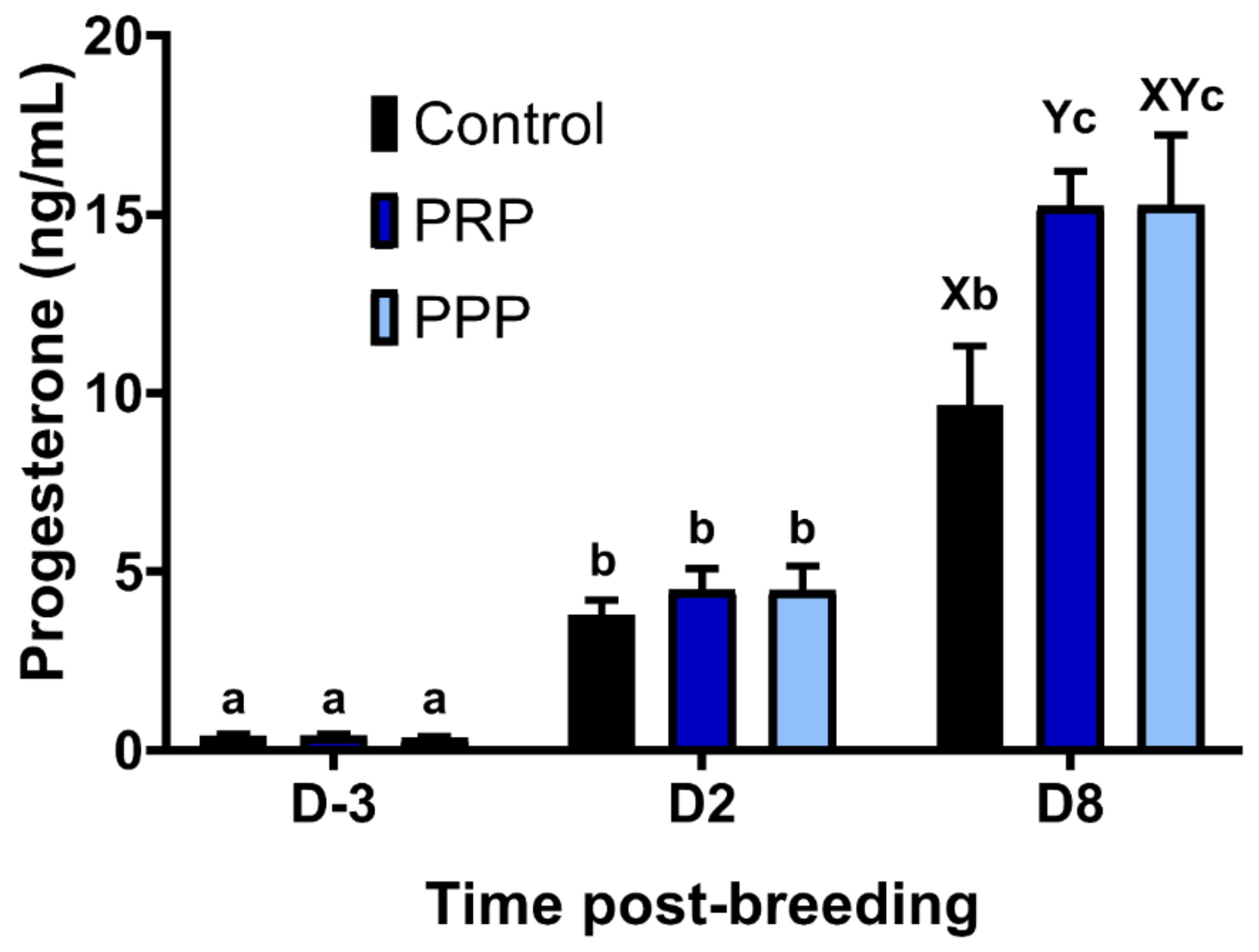
2.8. Endometrial Culture and Progesterone Concentrations
2.9. Embryo Recovery Rates
3. Discussion
4. Materials and Methods
4.1. Screening Mares for Susceptibility to PBIE
4.2. Experimental Design
4.3. Preparation of PRP and PPP
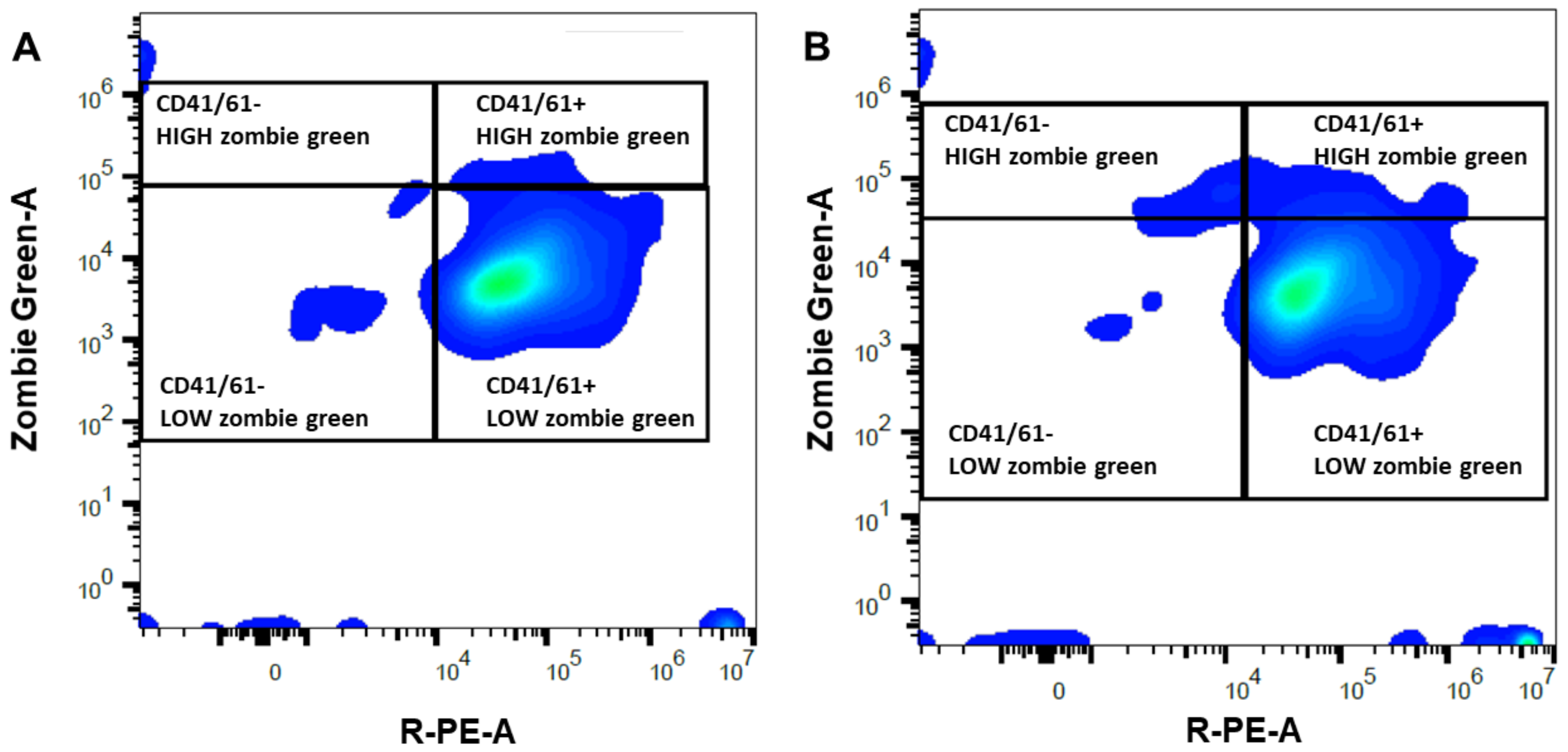
4.4. Assessment of Platelet Viability
4.5. Semen Collection and Processing
4.6. Aerobic Bacterial Culture
4.7. Endometrial Cytology
4.8. Low-Volume Lavage and Multiplex Immunoassay
4.9. Endometrial Biopsy: Inflammatory Cell Count and Immunolabelling for Progesterone Receptors
4.10. Progesterone Assay
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Traub-Dargatz, J.L.; Salman, M.D.; Voss, J.L. Medical problems of adult horses, as ranked by equine practitioners. J. Am. Vet. Med. Assoc. 1991, 198, 1745–1747. [Google Scholar] [PubMed]
- Troedsson, M.H.T. Uterine clearance and resistance to persistent endometritis in the mare. Theriogenology 1999, 52, 461–471. [Google Scholar] [CrossRef]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Betancourt, A.; Horohov, D.; Scoggin, K.E.; Squires, E.L.; Troedsson, M.H.T. Endometrial inflammatory markers of the early immune response in mares susceptible or resistant to persistent breeding-induced endometritis. Reproduction 2013, 145, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, M.H.; Liu, I.K.; Crabo, B. Sperm transport and survival in the mare. Theriogenology 1998, 49, 905–915. [Google Scholar] [CrossRef]
- Carnevale, E.M.; Ramirez, R.J.; Squires, E.L.; Alvarenga, M.A.; Vanderwall, D.K.; McCue, P.M. Factors affecting pregnancy rates and early embryonic death after equine embryo transfer. Theriogenology 2000, 54, 965–979. [Google Scholar] [CrossRef]
- Troedsson, M.H.T. Therapeutic considerations for mating-induced endometritis. Pferdeheilkd. Equine Med. 1997, 13, 516–520. [Google Scholar] [CrossRef]
- Troedsson, M.H.; Liu, I.K. Uterine clearance of non-antigenic markers (51Cr) in response to a bacterial challenge in mares potentially susceptible and resistant to chronic uterine infections. J. Reprod. Fertil. Suppl. 1991, 44, 283–288. [Google Scholar]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in mares—A multifaceted challenge: From clinical aspects to immunopathogenesis and pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef]
- Riddle, W.T.; LeBlanc, M.M.; Stromberg, A.J. Relationships between uterine culture, cytology and pregnancy rates in a Thoroughbred practice. Theriogenology 2007, 68, 395–402. [Google Scholar] [CrossRef]
- Collins, S. A study of the incidence of cervical and uterine infection in Thoroughbred mares in Ireland. Vet. Rec. 1964, 66, 673–676. [Google Scholar]
- Bain, A.M. The role of infection in infertility in the thoroughbred mare. Vet. Rec. 1966, 78, 168–173. [Google Scholar] [CrossRef]
- Leblanc, M.; Causey, R. Clinical and subclinical endometritis in the mare: Both threats to fertility. Reprod. Domest. Anim. 2009, 44, 10–22. [Google Scholar] [CrossRef]
- Scoggin, C.F. Endometritis: Nontraditional Therapies. Vet. Clin. North Am. Equine Pract. 2016, 32, 499–511. [Google Scholar] [CrossRef]
- Soares, C.S.; Babo, P.S.; Reis, R.L.; Carvalho, P.P.; Gomes, M.E. Platelet-Derived Products in Veterinary Medicine: A New Trend or an Effective Therapy? Trends Biotechnol. 2020, 20, 30206–30207. [Google Scholar] [CrossRef]
- Boswell, S.G.; Cole, B.J.; Sundman, E.A.; Karas, V.; Fortier, L.A. Platelet-rich plasma: A milieu of bioactive factors. Arthrosc. J. Arthrosc. Relat. Surg. 2012, 28, 429–439. [Google Scholar] [CrossRef]
- Farghali, H.A.; AbdElKader, N.A.; AbuBakr, H.O.; Aljuaydi, S.H.; Khattab, M.S.; Elhelw, R.; Elhariri, M. Antimicrobial action of autologous platelet-rich plasma on MRSA-infected skin wounds in dogs. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Gilbertie, J.M.; Schaer, T.P.; Schubert, A.G.; Jacob, M.E.; Menegatti, S.; Lavoie, R.A.; Schnabel, L. V Platelet-rich plasma lysate displays antibiofilm properties and restores antimicrobial activity against synovial fluid biofilms in vitro. J. Orthop. Res. 2020, 38, 1365–1374. [Google Scholar] [CrossRef]
- Álvarez, M.; López, C.; Giraldo, C.; Samudio, I.; Carmona, J. In vitro bactericidal activity of equine platelet concentrates, platelet poor plasma, and plasma against methicillin-resistant Staphylococcus aureus. Arch. Med. Vet. 2011, 43, 155–161. [Google Scholar] [CrossRef][Green Version]
- Hartwig, J.; Italiano, J. The birth of the platelet. J. Thromb. Haemost. 2003, 1, 1580–1586. [Google Scholar] [CrossRef]
- Bos-Mikich, A.; Ferreira, M.O.; de Oliveira, R.; Frantz, N. Platelet-rich plasma or blood-derived products to improve endometrial receptivity? J. Assist. Reprod. Genet. 2019, 36, 613–620. [Google Scholar] [CrossRef]
- Pavlovic, V.; Ciric, M.; Jovanovic, V.; Stojanovic, P. Platelet Rich Plasma: A short overview of certain bioactive components. Open Med. 2016, 11, 242–247. [Google Scholar] [CrossRef]
- Reghini, M.F.S.; Neto, C.R.; Segabinazzi, L.G.; Chaves, M.M.B.C.; Camila de Paula, F.; Bussiere, M.C.C.; Dell’Aqua, J.A., Jr.; Papa, F.O.; Alvarenga, M.A. Inflammatory response in chronic degenerative endometritis mares treated with platelet-rich plasma. Theriogenology 2016, 86, 516–522. [Google Scholar] [CrossRef]
- Metcalf, E.S.; Scoggin, K.; Troedsson, M.H.T. The effect of platelet-rich plasma on endometrial pro-inflammatory cytokines in susceptible mares following semen deposition. J. Equine Vet. Sci. 2012, 32, 498. [Google Scholar] [CrossRef]
- Metcalf, E.S. The effect of Platelet-Rich Plasma (PRP) on intraluminal fluid and pregnancy rates in mares susceptible to persistent mating-induced endometritis (PMIE). J. Equine Vet. Sci. 2014, 34, 128. [Google Scholar] [CrossRef]
- Segabinazzi, L.G.; Friso, A.M.; Correal, S.B.; Crespilho, A.M.; Dell’Aqua, J.A.; Miró, J.; Papa, F.O.; Alvarenga, M.A. Uterine clinical findings, fertility rate, leucocyte migration, and COX-2 protein levels in the endometrial tissue of susceptible mares treated with platelet-rich plasma before and after AI. Theriogenology 2017, 104, 120–126. [Google Scholar] [CrossRef]
- Pasch, L.; Schmidt, A.; King, W. Clinical observations after pre breeding intrauterine plasma infusion in 18 mares inseminated with thawed frozen semen. J. Equine Vet. Sci. 2021, 99, 103389. [Google Scholar] [CrossRef]
- Gilbertie, J.M.; Schaer, T.P.; Schubert, A.G.; Seiler, G.S.; Deddens, B.L.; Engiles, J.B.; Stowe, D.M.; Jacob, M.E. Platelet-rich plasma lysate improves bacterial load and outcomes of staphylococcus aureus infectious arthritis in horses. AAEP Proc. 2020, 66, 230–231. [Google Scholar]
- McCue, P.M.; DeLuca, C.A.; Ferris, R.A.; Wall, J.J. How to Evaluate Equine Embryos. In Proceedings of the 55th American Association of Equine Practitioners, Las Vegas, NV, USA, 5–9 December 2009; pp. 252–256. [Google Scholar]
- Aghajanova, L.; Houshdaran, S.; Balayan, S.; Manvelyan, E.; Irwin, J.C.; Huddleston, H.G.; Giudice, L.C. In vitro evidence that platelet-rich plasma stimulates cellular processes involved in endometrial regeneration. J. Assist. Reprod. Genet. 2018, 35, 757–770. [Google Scholar] [CrossRef]
- Tandulwadkar, S.; Naralkar, M.; Surana, A.; Selvakarthick, M.; Kharat, A. Autologous intrauterine platelet-rich plasma instillation for suboptimal endometrium in frozen embryo transfer cycles: A pilot study. J. Hum. Reprod. Sci. 2017, 10, 208–212. [Google Scholar] [CrossRef]
- Marini, M.G.; Perrini, C.; Esposti, P.; Corradetti, B.; Bizzaro, D.; Riccaboni, P.; Fantinato, E.; Urbani, G.; Gelati, G.; Cremonesi, F.; et al. Effects of platelet-rich plasma in a model of bovine endometrial inflammation in vitro. Reprod. Biol. Endocrinol. 2016, 14, 1–17. [Google Scholar] [CrossRef]
- Da Fontoura Pereira, R.C.; De La Côrte, F.D.; Brass, K.E.; da Silva Azevedo, M.; Gallio, M.; Cantarelli, C.; Dau, S.L.; Cezar, A.S.; Inkelmann, M.A. Evaluation of three methods of platelet-rich plasma for treatment of equine distal limb skin wounds. J. Equine Vet. Sci. 2019, 72, 1–7. [Google Scholar] [CrossRef]
- Carmona, J.U.; Argüelles, D.; Climent, F.; Prades, M. Autologous platelet concentrates as a treatment of horses with osteoarthritis: A preliminary pilot clinical study. J. Equine Vet. Sci. 2007, 27, 167–170. [Google Scholar] [CrossRef]
- Georg, R.; Maria, C.; Gisela, A.; Bianca, C. Autologous conditioned plasma as therapy of tendon and ligament lesions in seven horses. J. Vet. Sci. 2010, 11, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Fumuso, E.; Giguère, S.; Wade, J.; Rogan, D.; Videla-Dorna, I.; Bowden, R.A. Endometrial IL-1β, IL-6 and TNF-α, mRNA expression in mares resistant or susceptible to post-breeding endometritis. Effects of estrous cycle, artificial insemination and immunomodulation. Vet. Immunol. Immunopathol. 2003, 96, 31–41. [Google Scholar] [CrossRef]
- Christoffersen, M.; Woodward, E.M.; Bojesen, A.M.; Petersen, M.R.; Squires, E.L.; Lehn-Jensen, H.; Troedsson, M.H.T. Effect of immunomodulatory therapy on the endometrial inflammatory response to induced infectious endometritis in susceptible mares. Theriogenology 2012, 78, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, M.; Camilla Dooleweerdt, B.; Stine, J.; Anders Miki, B.; Morten Roenn, P.; Henrik, L.-J.; Bojesen, A.M.; Anders Miki, B.; Petersen, M.R.; Morten Roenn, P.; et al. Evaluation of the systemic acute phase response and endometrial gene expression of serum amyloid A and pro- and anti-inflammatory cytokines in mares with experimentally induced endometritis. Vet. Immunol. Immunopathol. 2010, 138, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, M.; Woodward, E.; Bojesen, A.M.; Jacobsen, S.; Petersen, M.R.; Troedsson, M.H.; Lehn-Jensen, H. Inflammatory responses to induced infectious endometritis in mares resistant or susceptible to persistent endometritis. BMC Vet. Res. 2012, 8, 41. [Google Scholar] [CrossRef]
- Marth, C.D.; Young, N.D.; Glenton, L.Y.; Noden, D.M.; Browning, G.F.; Krekeler, N. Deep sequencing of the uterine immune response to bacteria during the equine oestrous cycle. BMC Genomics 2015, 16, 1–19. [Google Scholar] [CrossRef]
- Bendinelli, P.; Matteucci, E.; Dogliotti, G.; Corsi, M.M.; Banfi, G.; Maroni, P.; Desiderio, M.A. Molecular basis of anti-inflammatory action of platelet-rich plasma on human chondrocytes: Mechanisms of NF-κB inhibition via HGF. J. Cell. Physiol. 2010, 225, 757–766. [Google Scholar] [CrossRef]
- Van Buul, G.M.; Koevoet, W.L.M.; Kops, N.; Bos, P.K.; Verhaar, J.A.N.; Weinans, H.; Bernsen, M.R.; van Osch, G.J.V.M. Platelet-rich plasma releasate inhibits inflammatory processes in osteoarthritic chondrocytes. Am. J. Sports Med. 2011, 39, 2362–2370. [Google Scholar] [CrossRef]
- Girling, J.E.; Hedger, M.P. Toll-like receptors in the gonads and reproductive tract: Emerging roles in reproductive physiology and pathology. Immunol. Cell Biol. 2007, 85, 481–489. [Google Scholar] [CrossRef]
- Chandrasekharan, N.V.; Simmons, D.L. The cyclooxygenases. Genome Biol. 2004, 5. [Google Scholar] [CrossRef]
- Fumuso, E.; Aguilar, J.; Giguère, S.; David, O.; Wade, J.; Rogan, D. Interleukin-8 (IL-8) and 10 (IL-10) mRNA transcriptions in the endometrium of normal mares and mares susceptible to persistent post-breeding endometritis. Anim. Reprod. Sci. 2006, 94, 282–285. [Google Scholar] [CrossRef]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar]
- Zerbe, H.; Schuberth, H.J.; Engelke, F.; Frank, J.; Klug, E.; Leibold, W. Development and comparison of in vivo and in vitro models for endometritis in cows and mares. Theriogenology 2003, 60, 209–223. [Google Scholar] [CrossRef]
- Drago, L.; Bortolin, M.; Vassena, C.; Romanò, C.L.; Taschieri, S.; Del Fabbro, M. Plasma components and platelet activation are essential for the antimicrobial properties of autologous platelet-rich plasma: An in vitro study. PLoS ONE 2014, 9, e107813. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Yeaman, M.R.; Selsted, M.E. Antimicrobial peptides from human platelets. Infect. Immun. 2002, 70, 6524–6533. [Google Scholar] [CrossRef]
- Anitua, E.; Alonso, R.; Girbau, C.; Aguirre, J.J.; Muruzabal, F.; Orive, G. Antibacterial effect of plasma rich in growth factors (PRGF®-Endoret®) against Staphylococcus aureus and Staphylococcus epidermidis strains. Clin. Exp. Dermatol. 2012, 37, 652–657. [Google Scholar] [CrossRef]
- Burnouf, T.; Chou, M.-L.; Wu, Y.-W.; Su, C.-Y.; Lee, L.-W. Antimicrobial activity of platelet (PLT)-poor plasma, PLT-rich plasma, PLT gel, and solvent/detergent-treated PLT lysate biomaterials against wound bacteria. Transfusion 2013, 53, 138–146. [Google Scholar] [CrossRef]
- Pascoe, D.R. Effect of Adding Autologous Plasma to an Intrauterine Antibiotic Therapy after Breeding on Pregnancy Rates in Mares1. Biol. Reprod. 1995, 52, 539–543. [Google Scholar] [CrossRef]
- Chen, J.; Losos, M.; Yang, S.; Li, J.; Wu, H.; Cataland, S. Increased complement activation during platelet storage. Transfusion 2017, 57, 2182–2188. [Google Scholar] [CrossRef]
- Troedsson, M.H.T.; Ababneh, M.M.; Ohlgren, A.F.; Madill, S.; Vetscher, N.; Gregas, M. Effect of periovulatory prostaglandin F2α on pregnancy rates and luteal function in the mare. Theriogenology 2001, 55, 1891–1899. [Google Scholar] [CrossRef]
- Canisso, I.F.; Gallacher, K.; Gilbert, M.A.; Korn, A.; Schweizer, C.M.; Bedford-Guaus, S.J.; Gilbert, R.O. Preovulatory progestagen treatment in mares fails to delay ovulation. Vet. J. 2013, 197, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Frois, A.R.C.; Lof, H.; Mattos, R.C. Effect of uterine inflammation by Streptococcus zooepidemicus on progesterone levels in mares. In Proceedings of the Reproducción Asistida en Equinos I, Bogotá, Colombia, 14–17 November 2017; p. 153. [Google Scholar]
- Lange-Consiglio, A.; Cazzaniga, N.; Garlappi, R.; Spelta, C.; Pollera, C.; Perrini, C.; Cremonesi, F. Platelet concentrate in bovine reproduction: Effects on in vitro embryo production and after intrauterine administration in repeat breeder cows. Reprod. Biol. Endocrinol. 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Contran, R.S.; Kumar, V. Robbins Pathologic Basic of Disease, 6th ed.; W. B. Saunders: Philadelphia, PA, USA, 1999. [Google Scholar]
- Brinsko, S.P.; Rigby, S.L.S.L.; Varner, D.D.; Blanchard, T.L. A practical method for recognizing mares susceptible to post-breeding endometritis. In Proceedings of the 49th Annual Conference of the American Association of Equine Practitioners, New Orleans, LA, USA, 21–25 November 2003; Volume 49, pp. 363–365. [Google Scholar]
- Tunón, A.M.; Ekwall, H.; Nummijärvi, A.; Rodríguez-Martínez, H. X-ray microanalysis of the secretory epithelium of the endometrial glands and intraluminal uterine fluid in oestrus mares. Reprod. Domest. Anim. 2000, 35, 221–227. [Google Scholar] [CrossRef]
- Henry, M.; Vandeplassche, G.; Coryn, M.; Spincemaille, J.; Vandeplassche, M. Excessive oedema of the genital tract in the mare. Zentralbl. Veterinarmed. A 1981, 28, 390–409. [Google Scholar] [CrossRef]
- Kenney, R.M.; Doig, P.A. Equine endometrial biopsy. In Current Therapy in Theriogenology; Morrow, D.A., Ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Pennell, E.N.; Wagner, K.H.; Mosawy, S.; Bulmer, A.C. Acute bilirubin ditaurate exposure attenuates ex vivo platelet reactive oxygen species production, granule exocytosis and activation. Redox Biol. 2019, 26, 101250. [Google Scholar] [CrossRef]
- O’Shea, C.M.; Werre, S.R.; Dahlgren, L.A. Comparison of Platelet Counting Technologies in Equine Platelet Concentrates. Vet. Surg. 2015, 44, 304–313. [Google Scholar] [CrossRef]
- Wagner, B.; Freer, H. Development of a bead-based multiplex assay for simultaneous quantification of cytokines in horses. Vet. Immunol. Immunopathol. 2009, 127, 242–248. [Google Scholar] [CrossRef]
- Skogstrand, K. Multiplex assays of inflammatory markers, a description of methods and discussion of precautions—Our experience through the last ten years. Methods 2012, 56, 204–212. [Google Scholar] [CrossRef]
- Fedorka, C.E.; Ball, B.A.; Scoggin, K.E.; Loux, S.C.; Troedsson, M.H.T.; Adams, A.A. The feto-maternal immune response to equine placentitis. Am. J. Reprod. Immunol. 2019, 82, 13179. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WB | PRP | PPP | |
|---|---|---|---|
| Platelets (103/µL) | 119.9 ± 30 b | 622.9 ± 144 a | 36.0 ± 25 c |
| RBC (106/µL) | 6.4 ± 0.9 a | 0.02 ± 0.022 b | 0.01 ± 0.011 b |
| WBC (103/µL) | 5.6 ± 1.2 a | <0.001 b | <0.001 b |
| D2 | D8 | |||||
|---|---|---|---|---|---|---|
| Mare ID | Control | PRP | PPP | Control | PRP | PPP |
| 1 | - | - | - | - | - | - |
| 2 | - | - | Enterococcus spp. | - | - | - |
| 3 | Streptococcus β-hemolytic | - | - | Streptococcus β-hemolytic | - | - |
| 4 | - | - | - | - | - | Klebsiella pneumoniae |
| 5 | Klebsiella pneumoniae | - | - | Klebsiella pneumoniae | - | - |
| 6 | - | - | - | - | - | - |
| 7 | Streptococcus β-hemolytic | - | Streptococcus β-hemolytic | - | - | - |
| 8 | - | - | - | Escherichia coli | - | - |
| 9 | - | - | Streptococcus β-hemolytic | Escherichia coli | - | Streptococcus β-hemolytic |
| 10 | - | - | NP | - | - | NP |
| 11 | - | - | NP | Escherichia coli | - | NP |
| 12 | - | - | - | - | - | - |
| Control | PRP | PPP | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mare ID | Embryo STAGE | Quality Grade | Diameter (µm) | Embryo Stage | Quality | Diameter (µm) | Embryo Stage | Quality Grade | Diameter (µm) |
| 1 | - | - | Blastocyst | 1 | 340 | Blastocyst | 1 | 260 | |
| Expanded blastocyst | 1 | 1100 | |||||||
| 2 | - | - | - | - | - | - | |||
| 3 | - | - | Expanded blastocyst | 1 | 1230 | Early blastocyst | 1 | 175 | |
| 4 | Expanded blastocyst | 1 | 540 | Expanded blastocyst | 1 | 860 | Expanded blastocyst | 1 | 840 |
| 5 | - | - | - | - | - | - | |||
| 6 | Early blastocyst | 2 | 195 | Expanded blastocyst | 1 | 1360 | - | - | |
| 7 | Blastocyst | 1 | 960 | Early blastocyst | 2 | 190 | Expanded blastocyst | 1 | 940 |
| 8 | - | - | Blastocyst | 1 | 340 | Expanded blastocyst | 1 | 650 | |
| 9 | - | - | Blastocyst | 1 | 300 | - | - | ||
| 10 | - | - | Blastocyst | 1 | 360 | NP | NP | ||
| 11 | - | - | Early blastocyst | 1 | 165 | NP | NP | ||
| 12 | Blastocyst | 1 | 360 | Blastocyst | 1 | 280 | Blastocyst | 1 | 320 |
| Blastocyst | 1 | 360 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segabinazzi, L.G.T.M.; Canisso, I.F.; Podico, G.; Cunha, L.L.; Novello, G.; Rosser, M.F.; Loux, S.C.; Lima, F.S.; Alvarenga, M.A. Intrauterine Blood Plasma Platelet-Therapy Mitigates Persistent Breeding-Induced Endometritis, Reduces Uterine Infections, and Improves Embryo Recovery in Mares. Antibiotics 2021, 10, 490. https://doi.org/10.3390/antibiotics10050490
Segabinazzi LGTM, Canisso IF, Podico G, Cunha LL, Novello G, Rosser MF, Loux SC, Lima FS, Alvarenga MA. Intrauterine Blood Plasma Platelet-Therapy Mitigates Persistent Breeding-Induced Endometritis, Reduces Uterine Infections, and Improves Embryo Recovery in Mares. Antibiotics. 2021; 10(5):490. https://doi.org/10.3390/antibiotics10050490
Chicago/Turabian StyleSegabinazzi, Lorenzo G. T. M., Igor F. Canisso, Giorgia Podico, Lais L. Cunha, Guilherme Novello, Michael F. Rosser, Shavahn C. Loux, Fabio S. Lima, and Marco A. Alvarenga. 2021. "Intrauterine Blood Plasma Platelet-Therapy Mitigates Persistent Breeding-Induced Endometritis, Reduces Uterine Infections, and Improves Embryo Recovery in Mares" Antibiotics 10, no. 5: 490. https://doi.org/10.3390/antibiotics10050490
APA StyleSegabinazzi, L. G. T. M., Canisso, I. F., Podico, G., Cunha, L. L., Novello, G., Rosser, M. F., Loux, S. C., Lima, F. S., & Alvarenga, M. A. (2021). Intrauterine Blood Plasma Platelet-Therapy Mitigates Persistent Breeding-Induced Endometritis, Reduces Uterine Infections, and Improves Embryo Recovery in Mares. Antibiotics, 10(5), 490. https://doi.org/10.3390/antibiotics10050490