
Efficacy of Novel Bacteriophages against Escherichia coli Biofilms on Stainless Steel

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Biofilm Formation Ability of E. coli Strains
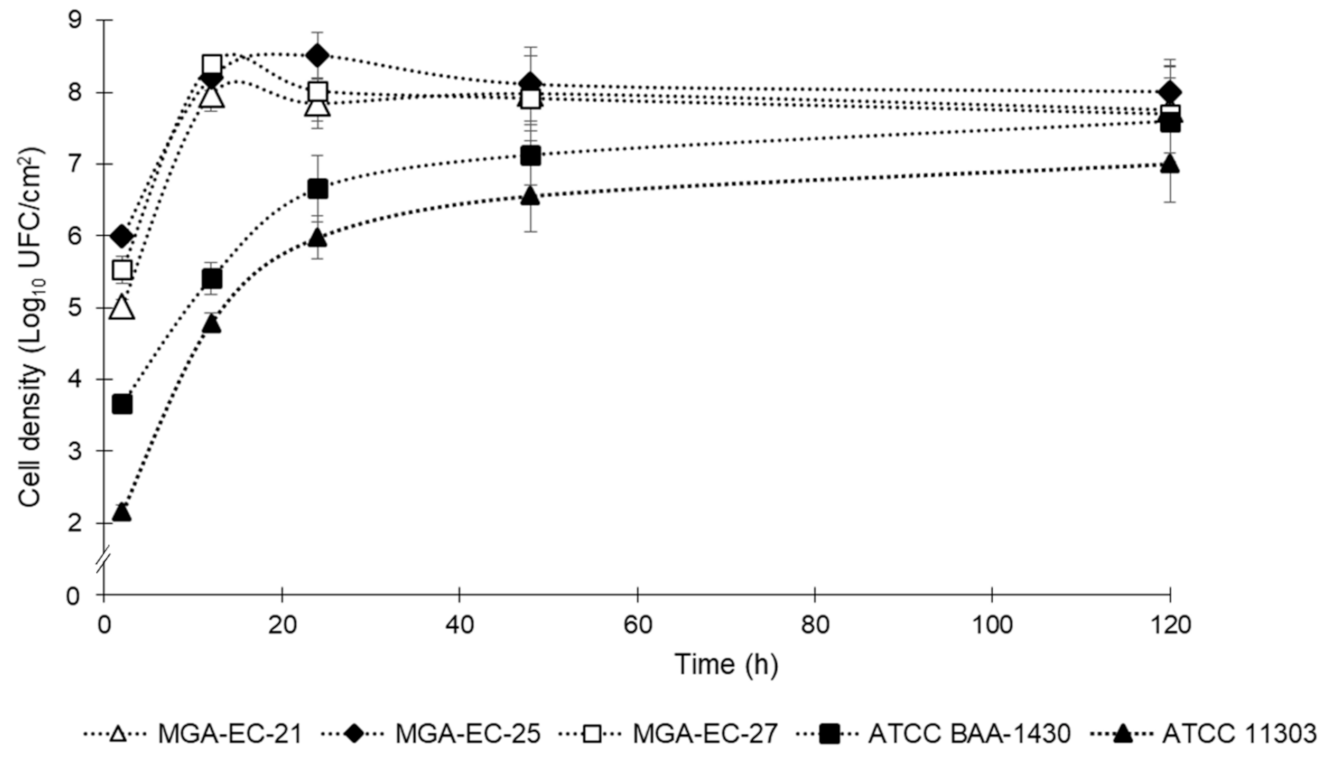
2.2. Biofilm Development Curve
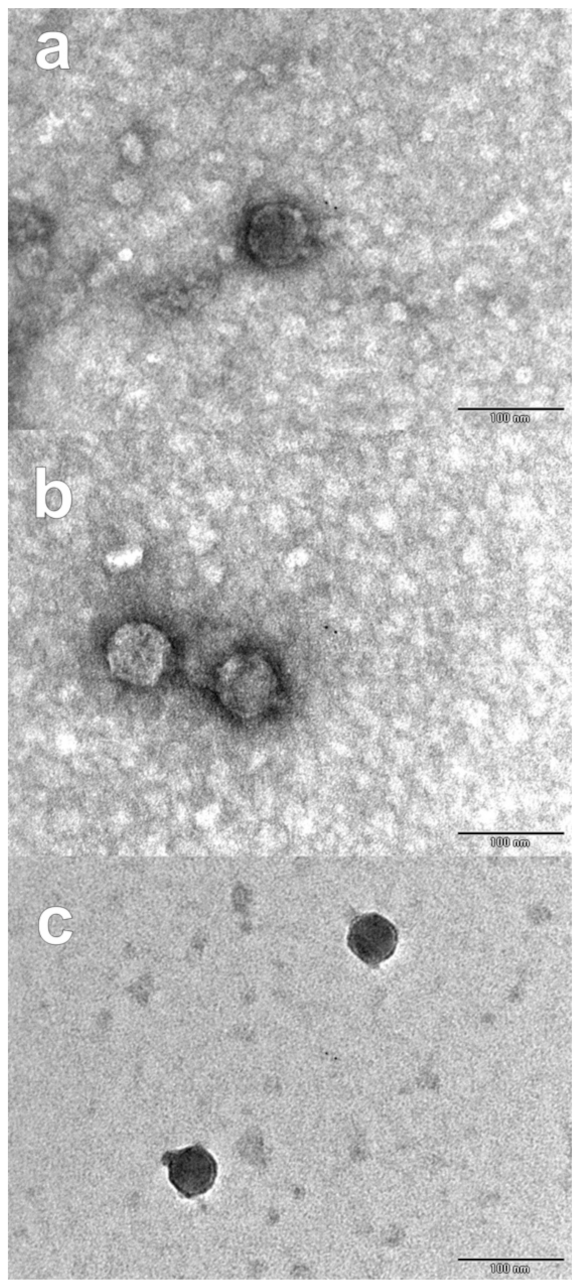
2.3. Bacteriophages Isolation and Characterization
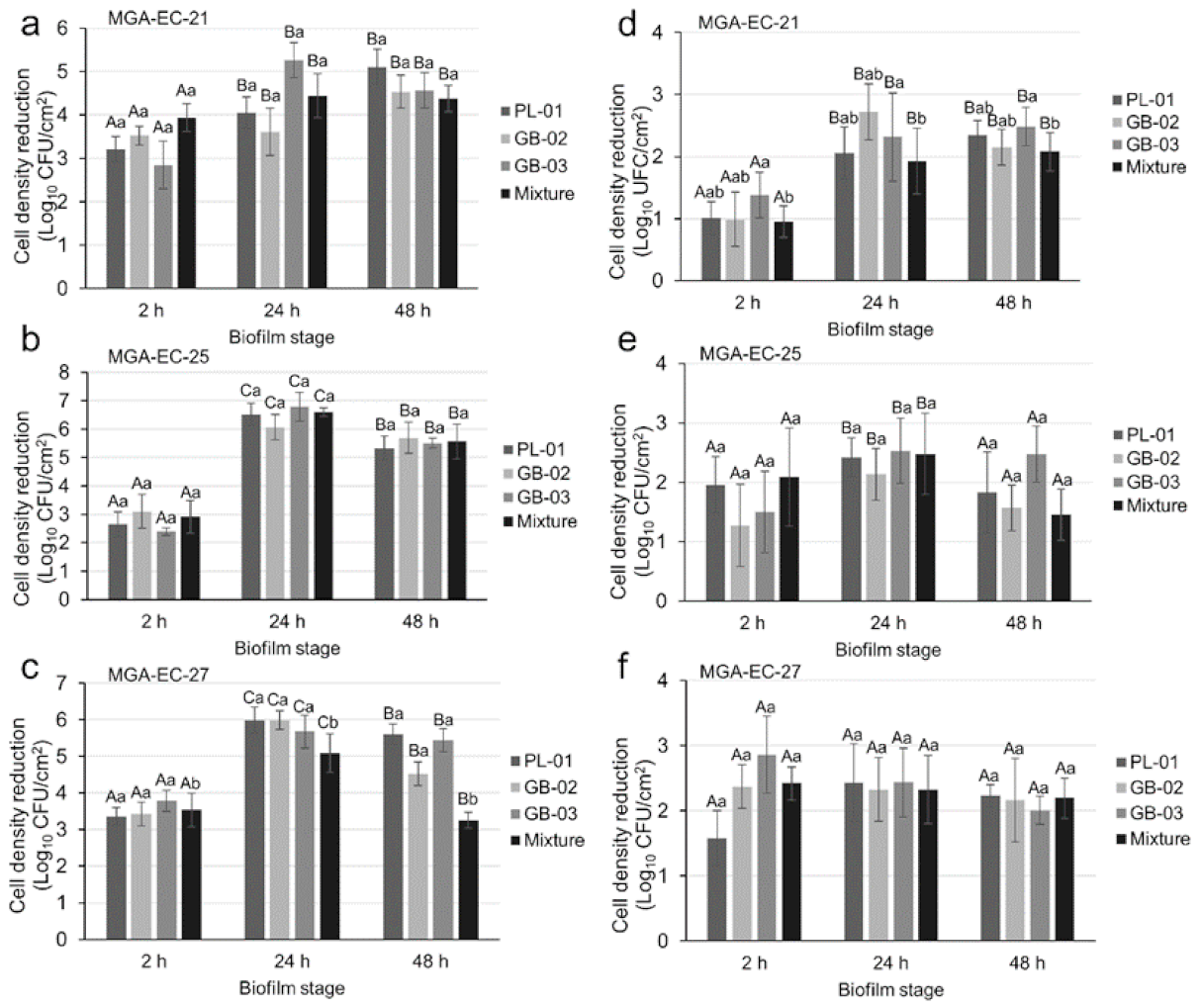
2.4. Biofilm Reduction Efficacy of Bacteriophages
2.5. Scanning Electron Microscope Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Characterization of the Strains’ Biofilm-Forming Ability
4.2.1. Production of Extracellular Polymeric Substances (EPS)
4.2.2. Semiquantitative Adherence Assay
4.3. Biofilm Formation on Stainless Steel
4.3.1. Biofilm Quantification
4.3.2. Scanning Electron Microscopy
4.4. Bacteriophage Isolation
4.4.1. Phage Host Range
4.4.2. Phage Morphology Determined by Transmission Electron Microscopy
4.5. Biofilm Exposure to Bacteriophages
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as weapons against bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef]
- Myszka, K.; Czaczyk, K. Bacterial biofilms on food contact surfaces—A review. Pol. J. Food Nutr. Sci. 2011, 61, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Anand, S.; Singh, A. Resistance of the constitutive microflora of biofilms formed on whey reverse-osmosis membranes to individual cleaning steps of a typical clean-in-place protocol. J. Dairy Sci. 2013, 96, 6213–6222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, M.Q.; Quinones, B.; He, X.; Zhong, W.; Louie, J.W.; Lee, B.G.; Yambao, J.C.; Mandrell, R.E.; Cooley, M.B. An environmental Shiga toxin-producing Escherichia coli O145 clonal population exhibits high-level phenotypic variation that includes virulence traits. Appl. Environ. Microbiol. 2016, 82, 1090–1101. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2009, 8, 26–38. [Google Scholar] [CrossRef]
- Orhan-Yanıkan, E.; da Silva-Janeiro, S.; Ruiz-Rico, M.; Jiménez-Belenguer, A.I.; Ayhan, K.; Barat, J.M. Essential oils compounds as antimicrobial and antibiofilm agents against strains present in the meat industry. Food Control. 2019, 101, 29–38. [Google Scholar] [CrossRef]
- Patel, J.; Sharma, M.; Millner, P.; Calaway, T.; Singh, M. Inactivation of Escherichia coli O157:H7 attached to spinach harvester blade using bacteriophage. Foodborne Pathog. Dis. 2011, 8, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Ryu, J.-H.; Beuchat, L.R. Inactivation of Escherichia coli O157:H7 in biofilm on stainless steel by treatment with an alkaline cleaner and a bacteriophage. J. Appl. Microbiol. 2005, 99, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Polaska, M.; Sokolowska, B. Bacteriophages-a new hope or a huge problem in the food industry. AIMS Microbiol. 2019, 5, 324–346. [Google Scholar] [CrossRef]
- Lang, L.H. FDA approves use of bacteriophages to be added to meat and poultry products. Gastroenterology 2006, 131, 1370. [Google Scholar] [CrossRef]
- García, P.; Madera, C.; Martínez, B.; Rodríguez, A. Biocontrol of Staphylococcus aureus in curd manufacturing processes using bacteriophages. Int. Dairy J. 2007, 17, 1232–1239. [Google Scholar] [CrossRef]
- Greer, G.G.; Dilts, B.D.; Ackermann, H.-W. Characterization of a Leuconostoc gelidum bacteriophage from pork. Int. J. food Microbiol. 2007, 114, 370–375. [Google Scholar] [CrossRef]
- Heringa, S.D.; Kim, J.; Jiang, X.; Doyle, M.P.; Erickson, M.C. Use of a mixture of bacteriophages for biological control of Salmonella enterica strains in compost. Appl. Environ. Microbiol. 2010, 76, 5327–5332. [Google Scholar] [CrossRef] [Green Version]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Diez-Gonzalez, F. Reduction of Escherichia coli O157:H7 viability on hard surfaces by treatment with a bacteriophage mixture. Int. J. Food Microbiol. 2011, 145, 37–42. [Google Scholar] [CrossRef]
- Ryan, E.M.; Alkawareek, M.Y.; Donnelly, R.F.; Gilmore, B.F. Synergistic phage-antibiotic combinations for the control of Escherichia coli biofilms in vitro. FEMS Immunol. Med. Microbiol. 2012, 65, 395–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, M.; Simões, L.E.; Vieria, M.J. A review of current and emergent biofilm control strategies. LWT-Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, I.W.; Hughes, K.A.; Skillman, L.C.; Tait, K. The interaction of phage and biofilms. FEMS Microbiol. Lett. 2004, 232, 1–6. [Google Scholar] [CrossRef]
- Mah, T.-F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Onmaz, N.E.; Yildirim, Y.; Karadal, F.; Hizlisoy, H.; Al, S.; Gungor, C.; Disli, H.B.; Barel, M.; Dishan, A.; Akai Tegin, R.A.; et al. Escherichia coli O157 in fish: Prevalence, antimicrobial resistance, biofilm formation capacity, and molecular characterization. LWT 2020, 133, 109940. [Google Scholar] [CrossRef]
- Bhardwaj, D.K.; Taneja, N.K.; Dp, S.; Chakotiya, A.; Patel, P.; Taneja, P.; Sachdev, D.; Gupta, S.; Sanal, M.G. Phenotypic and genotypic characterization of biofilm forming, antimicrobial resistant, pathogenic Escherichia coli isolated from Indian dairy and meat products. Int. J. Food Microbiol. 2021, 336, 108899. [Google Scholar] [CrossRef]
- Montso, P.K.; Mlambo, V.; Ateba, C.N. Efficacy of novel phages for control of multi-drug resistant Escherichia coli O177 on artificially contaminated beef and their potential to disrupt biofilm formation. Food Microbiol. 2021, 94, 103647. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, K.V.G.; Ribeiro, C.; Dias, R.S.; Cardoso, S.A.; de Paula, S.O.; Zanuncio, J.C.; de Oliveira, L.L. Bacteriophage Isolated from Sewage Eliminates and Prevents the Establishment of Escherichia coli Biofilm. Adv. Pharm. Bull. 2018, 8, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Beloin, C.; Roux, A.; Ghigo, J.M. Escherichia coli Biofilms. In Bacterial Biofilms; Romeo, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 249–289. [Google Scholar]
- Bridier, A.; Sanchez-Vizuete, P.; Guilbaud, M.; Piard, J.C.; Naitali, M.; Briandet, R. Biofilm-associated persistence of food-borne pathogens. Food Microbiol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Iniguez-Moreno, M.; Gutierrez-Lomeli, M.; Avila-Novoa, M.G. Kinetics of biofilm formation by pathogenic and spoilage microorganisms under conditions that mimic the poultry, meat, and egg processing industries. Int. J. Food Microbiol. 2019, 303, 32–41. [Google Scholar] [CrossRef]
- Carson, L.; Gorman, S.P.; Gilmore, B.F. The use of lytic bacteriophages in the prevention and eradication of biofilms of Proteus mirabilis and Escherichia coli. FEMS Immunol. Med. Microbiol. 2010, 59, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Marshall, K.M.; Niebuhr, S.E.; Acuff, G.R.; Lucia, L.M.; Dickson, J.S. Identification of Escherichia coli O157:H7 meat processing indicators for fresh meat through comparison of the effects of selected antimicrobial interventions. J. Food Prot. 2005, 68, 2580–2586. [Google Scholar] [CrossRef]
- Wang, C.; Hang, H.; Zhou, S.; Niu, Y.D.; Du, H.; Stanford, K.; McAllister, T.A. Bacteriophage biocontrol of Shiga toxigenic Escherichia coli (STEC) O145 biofilms on stainless steel reduces the contamination of beef. Food Microbiol. 2020, 92, 103572. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-W.; Lee, H.-Y.; Kang, D.-H. Synergistic bactericidal effect of hot water with citric acid against Escherichia coli O157:H7 biofilm formed on stainless steel. Food Microbiol. 2021, 95, 103676. [Google Scholar] [CrossRef] [PubMed]
- Iniguez-Moreno, M.; Avila-Novoa, M.G.; Gutierrez-Lomeli, M. Resistance of pathogenic and spoilage microorganisms to disinfectants in the presence of organic matter and their residual effect on stainless steel and polypropylene. J. Glob. Antimicrob. Resist. 2018, 14, 197–201. [Google Scholar] [CrossRef]
- Ackermann, H.W.; DuBow, M.S. Practical applications of bacteriophages. In Viruses of Prokaryotes: General Properties of Bacteriophages; Ackermann, H.W., DuBow, M.S., Eds.; CRC Press: Boca Raton, FL, USA, 1987; pp. 143–158. [Google Scholar]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; van Zyl, L.J.; Sarkar, B.L.; Dutilh, B.E.; Alfenas-Zerbini, P.; Lobocka, M.; Tong, Y.; Brister, J.R.; et al. Taxonomy of prokaryotic viruses: 2018–2019 update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Arch. Virol. 2020, 165, 1253–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Tsonos, J.; Vandenheuvel, D.; Briers, Y.; De Greve, H.; Hernalsteens, J.-P.; Lavigne, R. Hurdles in bacteriophage therapy: Deconstructing the parameters. Vet. Microbiol. 2014, 171, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Sadekuzzaman, M.; Yang, S.; Mizan, M.F.R.; Ha, S.-D. Reduction of Escherichia coli O157:H7 in Biofilms Using Bacteriophage BPECO 19. J. Food Sci. 2017, 82, 1433–1442. [Google Scholar] [CrossRef]
- Barbirz, S.; Becker, M.; Freiberg, A.; Seckler, R. Phage tailspike proteins with beta-solenoid fold as thermostable carbohydrate binding materials. Macromol. Biosci. 2009, 9, 169–173. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariana, N.S.; Salman, S.A.; Neela, V.; Zamberi, S. Evaluation of modified Congo red agar for detection of biofilm produced by clinical isolates of methicillin–resistance Staphylococcus aureus. Afr. J. Microbiol. Res. 2009, 3, 330–338. [Google Scholar]
- Milanov, D.; Lazić, S.; Vidić, B.; Petrović, J.; Bugarski, D.; Šegulev, Z. Slime production and biofilm forming ability by Staphylococcus aureus bovine mastitis isolates. Acta Vet. 2010, 60, 217–226. [Google Scholar] [CrossRef]
- Marques, S.C.; Rezende, J.d.G.O.S.; de Freitas Alves, L.A.; Silva, B.C.; Alves, E.; de Abreu, L.R.; Piccoli, R.H. Formation of biofilms by Staphylococcus aureus on stainless steel and glass surfaces and its resistance to some selected chemical sanitizers. Braz. J. Microbiol. 2007, 38, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Michu, E.; Cervinkova, D.; Babak, V.; Kyrova, K.; Jaglic, Z. Biofilm formation on stainless steel by Staphylococcus epidermidis in milk and influence of glucose and sodium chloride on the development of ica-mediated biofilm. Int. Dairy J. 2011, 21, 179–184. [Google Scholar] [CrossRef]
- Alhede, M.; Qvortrup, K.; Liebrechts, R.; Hoiby, N.; Givskov, M.; Bjarnsholt, T. Combination of microscopic techniques reveals a comprehensive visual impression of biofilm structure and composition. FEMS Immunol. Med. Microbiol. 2012, 65, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fratesi, S.E.; Lynch, F.L.; Kirkland, B.L.; Brown, L.R. Effects of SEM Preparation Techniques on the Appearance of Bacteria and Biofilms in the Carter Sandstone. J. Sediment. Res. 2004, 74, 858–867. [Google Scholar] [CrossRef]
- Carey-Smith, G.V.; Billington, C.; Cornelius, A.J.; Hudson, J.A.; Heinemann, J.A. Isolation and characterization of bacteriophages infecting Salmonella spp. FEMS Microbiol. Lett. 2006, 258, 182–186. [Google Scholar] [CrossRef] [Green Version]
- López-Cuevas, O.; Castro-del Campo, N.; León-Félix, J.; Chaidez, C. Characterization of bacteriophages with a lytic effect on various Salmonella serotypes and Escherichia coli O157:H7. Can. J. Microbiol. 2011, 57, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Betancourt, M.; Shackelford, S.D.; Arthur, T.M.; Westmoreland, K.E.; Bellinger, G.; Rossman, M.; Reagan, J.O.; Koohmaraie, M. Prevalence of Escherichia coli O157:H7, Listeria monocytogenes, and Salmonella in two geographically distant commercial beef processing plants in the United States. J. Food Prot. 2004, 67, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossoni, E.M.M.; Gaylarde, C.C. Comparison of sodium hypochlorite and peracetic acid as sanitising agents for stainless steel food processing surfaces using epifluorescence microscopy. Int. J. Food Microbiol. 2000, 61, 81–85. [Google Scholar] [CrossRef]
- Jamalludeen, N.; Johnson, R.P.; Friendship, R.; Kropinski, A.M.; Lingohr, E.J.; Gyles, C.L. Isolation and characterization of nine bacteriophages that lyse O149 enterotoxigenic Escherichia coli. Vet. Microbiol. 2007, 124, 47–57. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Adherence Assay | Phenotype CRA b | |
|---|---|---|---|
| OD (λ = 570) | Adhesion Ability a | ||
| MGA-EC-01 | 0.076 ± 0.008 | Null | EPS producer |
| MGA-EC-02 | 0.073 ± 0.008 | Null | EPS producer |
| MGA-EC-08 | 0.066 ± 0.004 | Null | EPS producer |
| MGA-EC-21 | 0.095 ± 0.021 | Weak | EPS producer |
| MGA-EC-23 | 0.067 ± 0.005 | Null | EPS producer |
| MGA-EC-25 | 0.898 ± 0.113 | Strong | EPS producer |
| MGA-EC-26 | 0.093 ± 0.026 | Null | EPS producer |
| MGA-EC-27 | 0.968 ± 0.042 | Strong | EPS producer |
| MGA-EC-28 | 0.071 ± 0.007 | Null | EPS producer |
| MGA-EC-30 | 0.062 ± 0.001 | Null | EPS producer |
| ATCC BAA-1430 | 0.074 ± 0.003 | Null | EPS producer |
| ATCC 11303 | 0.174 ± 0.018 | Weak | EPS producer |
| Phage Strain | Morphology a | Source | Host Range (MGA-EC Strains) b | Plaque Diameter c | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 01 | 02 | 08 | 21 | 23 | 25 | 26 | 27 | 28 | 30 | ||||
| PL-01 | Podovirus | Poultry liver | - | ± | ± | ++ | - | ++ | - | ++ | - | - | 3 mm |
| GB-02 | Podovirus | Ground beef | ± | - | - | + | - | + | - | + | - | + | 4 mm |
| GB-03 | Podovirus | Ground beef | ± | - | - | ++ | - | ++ | - | ++ | - | ± | 3.5 mm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Gómez, J.P.; González-Torres, B.; Guerrero-Medina, P.J.; López-Cuevas, O.; Chaidez, C.; Avila-Novoa, M.G.; Gutiérrez-Lomelí, M. Efficacy of Novel Bacteriophages against Escherichia coli Biofilms on Stainless Steel. Antibiotics 2021, 10, 1150. https://doi.org/10.3390/antibiotics10101150
González-Gómez JP, González-Torres B, Guerrero-Medina PJ, López-Cuevas O, Chaidez C, Avila-Novoa MG, Gutiérrez-Lomelí M. Efficacy of Novel Bacteriophages against Escherichia coli Biofilms on Stainless Steel. Antibiotics. 2021; 10(10):1150. https://doi.org/10.3390/antibiotics10101150
Chicago/Turabian StyleGonzález-Gómez, Jean Pierre, Berenice González-Torres, Pedro Javier Guerrero-Medina, Osvaldo López-Cuevas, Cristóbal Chaidez, María Guadalupe Avila-Novoa, and Melesio Gutiérrez-Lomelí. 2021. "Efficacy of Novel Bacteriophages against Escherichia coli Biofilms on Stainless Steel" Antibiotics 10, no. 10: 1150. https://doi.org/10.3390/antibiotics10101150
APA StyleGonzález-Gómez, J. P., González-Torres, B., Guerrero-Medina, P. J., López-Cuevas, O., Chaidez, C., Avila-Novoa, M. G., & Gutiérrez-Lomelí, M. (2021). Efficacy of Novel Bacteriophages against Escherichia coli Biofilms on Stainless Steel. Antibiotics, 10(10), 1150. https://doi.org/10.3390/antibiotics10101150