Development of a Modular miRNA-Responsive Biosensor for Organ-Specific Evaluation of Liver Injury

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Instruments
2.3. Polyacrylamide Gel Electrophoresis (PAGE)
2.4. Fluorescence Measurement
2.5. Cell Culture
2.6. Cell Imaging
2.7. Animal Studies
2.8. Drug-Induced Acute Liver Injury Model
2.9. Statistics
3. Results
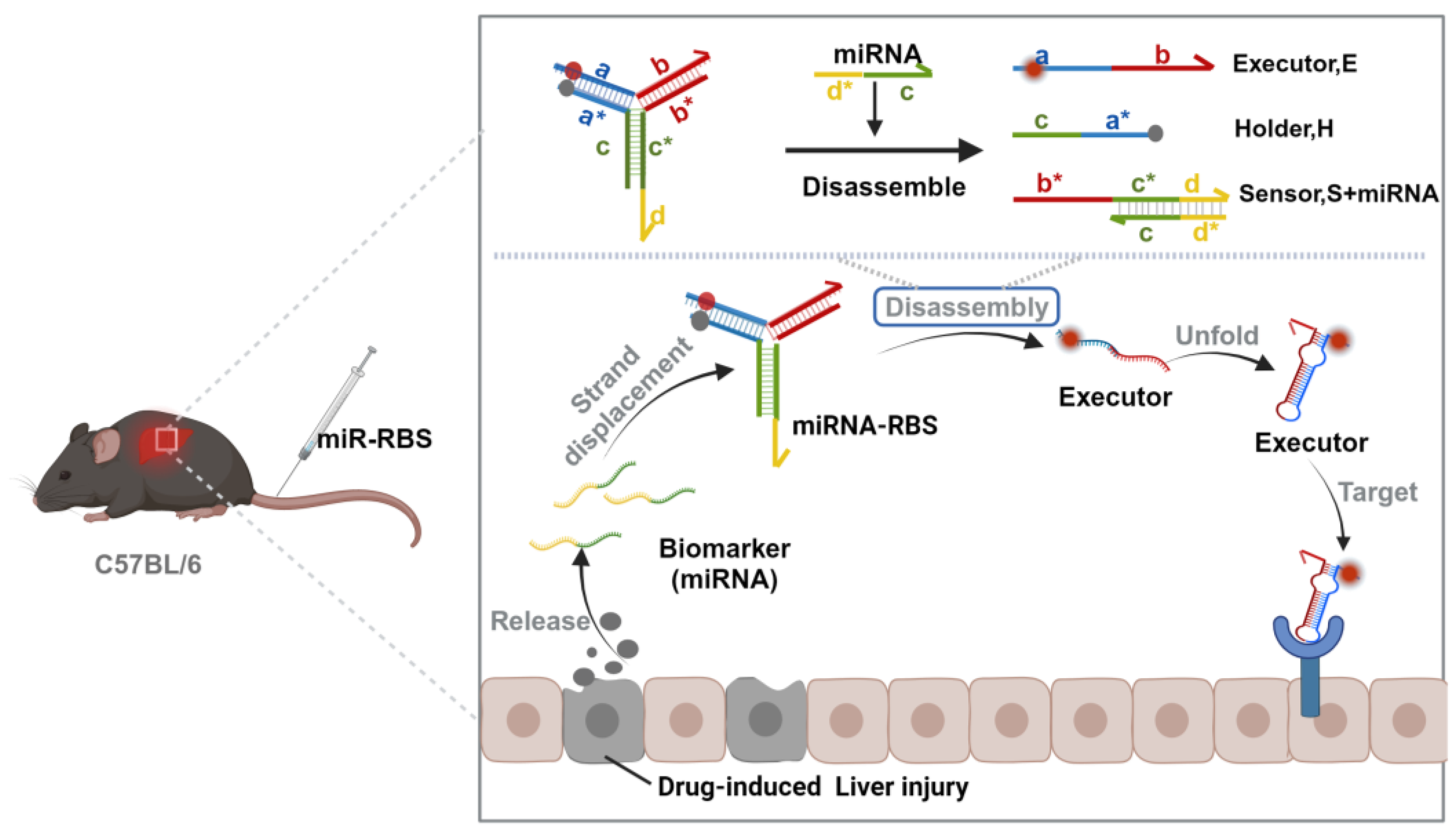
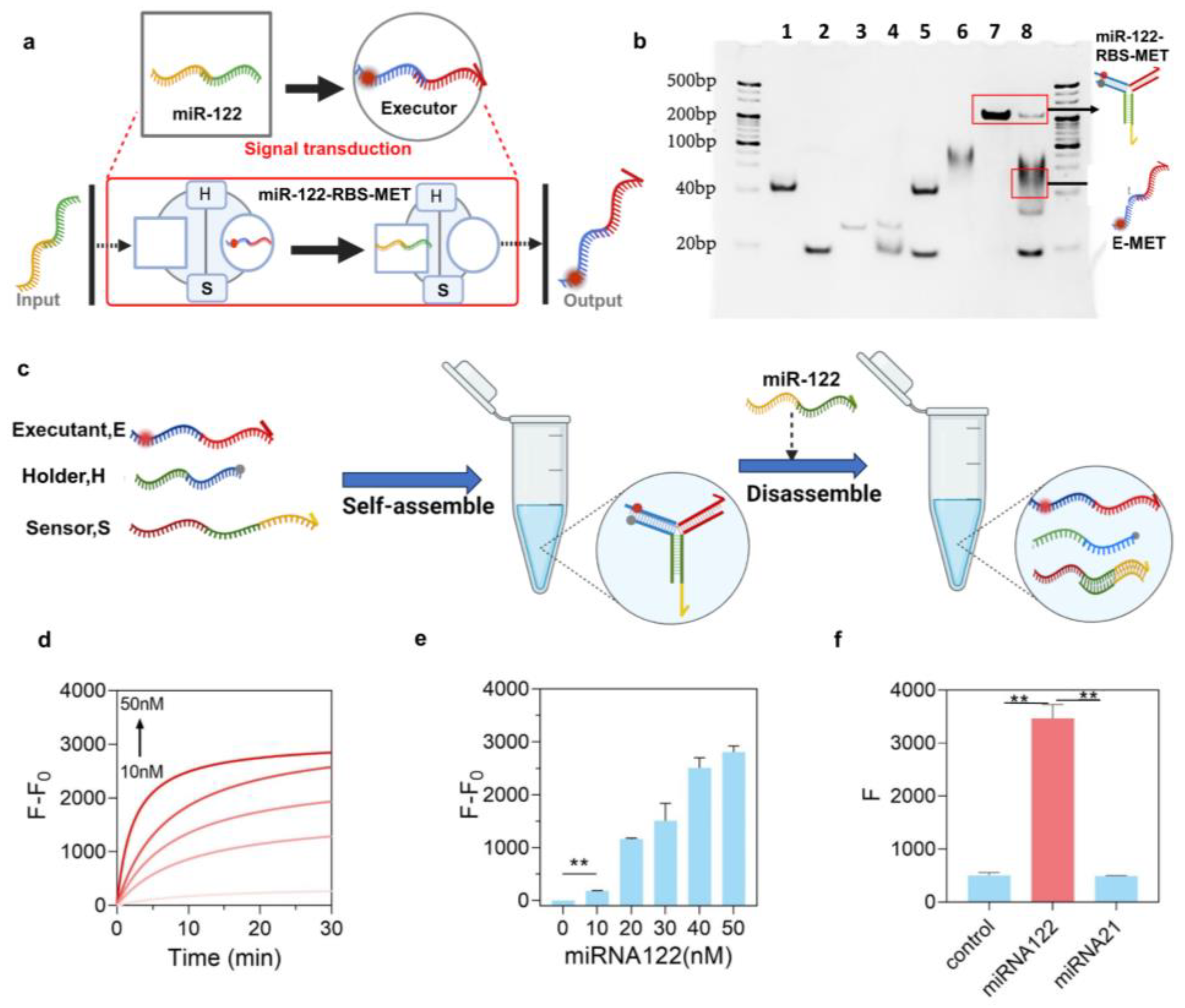
3.1. Design and Working Model of miR-RBS
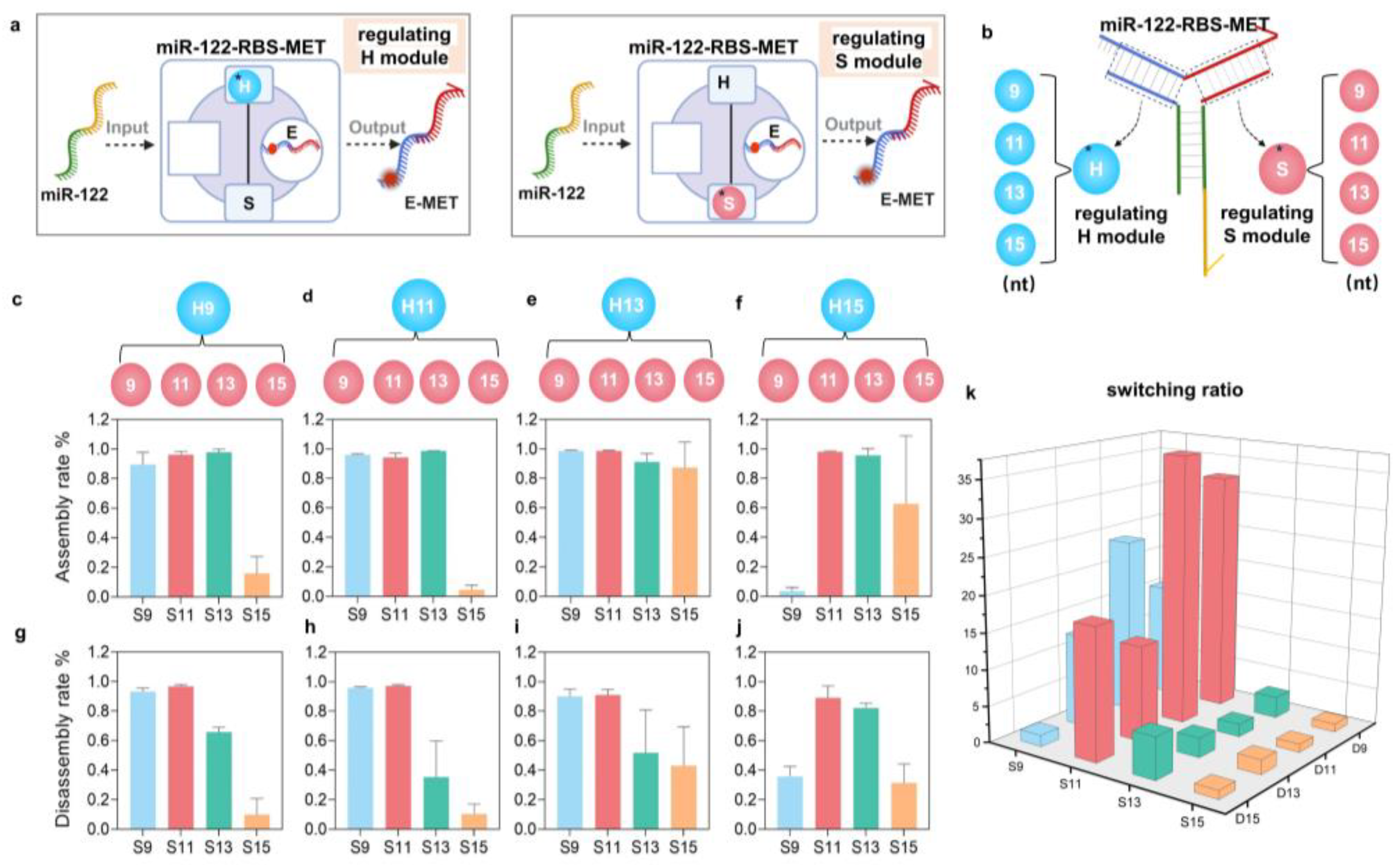
3.2. The Tunable Performance of miR-RBS
3.3. Modularity of miR-RBS for Cell-Specific miRNA Visualization
3.4. Organ-Specific Visualization of Disease-Associated miRNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biomarkers and Surrogate Endpoints: Preferred Definitions and Conceptual Framework. Available online: https://pubmed.ncbi.nlm.nih.gov/11240971/ (accessed on 29 June 2024).
- Bénichou, C. Criteria of Drug-Induced Liver Disorders. Report of an International Consensus Meeting. J. Hepatol. 1990, 11, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Nik Mohamed Kamal, N.N.S.B.; Shahidan, W.N.S. Non-Exosomal and Exosomal Circulatory MicroRNAs: Which Are More Valid as Biomarkers? Front. Pharmacol. 2019, 10, 1500. [Google Scholar] [CrossRef] [PubMed]
- Valihrach, L.; Androvic, P.; Kubista, M. Circulating miRNA Analysis for Cancer Diagnostics and Therapy. Mol. Asp. Med. 2020, 72, 100825. [Google Scholar] [CrossRef] [PubMed]
- Usuba, W.; Urabe, F.; Yamamoto, Y.; Matsuzaki, J.; Sasaki, H.; Ichikawa, M.; Takizawa, S.; Aoki, Y.; Niida, S.; Kato, K.; et al. Circulating miRNA Panels for Specific and Early Detection in Bladder Cancer. Cancer Sci. 2019, 110, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef]
- Hou, W.; Tian, Q.; Zheng, J.; Bonkovsky, H.L. MicroRNA-196 Represses Bach1 Protein and Hepatitis C Virus Gene Expression in Human Hepatoma Cells Expressing Hepatitis C Viral Proteins. Hepatology 2010, 51, 1494–1504. [Google Scholar] [CrossRef]
- Girard, M.; Jacquemin, E.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. miR-122, a Paradigm for the Role of microRNAs in the Liver. J. Hepatol. 2008, 48, 648–656. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of Tissue-Specific microRNAs from Mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S.; Marzolf, B.; Troisch, P.; Brightman, A.; Hu, Z.; Hood, L.E.; Galas, D.J. Circulating microRNAs, Potential Biomarkers for Drug-Induced Liver Injury. Proc. Natl. Acad. Sci. USA 2009, 106, 4402–4407. [Google Scholar] [CrossRef]
- Starkey Lewis, P.J.; Dear, J.; Platt, V.; Simpson, K.J.; Craig, D.G.N.; Antoine, D.J.; French, N.S.; Dhaun, N.; Webb, D.J.; Costello, E.M.; et al. Circulating microRNAs as Potential Markers of Human Drug-Induced Liver Injury. Hepatology 2011, 54, 1767–1776. [Google Scholar] [CrossRef]
- Dear, J.W.; Antoine, D.J.; Starkey-Lewis, P.; Goldring, C.E.; Park, B.K. Early Detection of Paracetamol Toxicity Using Circulating Liver microRNA and Markers of Cell Necrosis. Br. J. Clin. Pharmacol. 2014, 77, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Antoine, D.J.; Dear, J.W.; Lewis, P.S.; Platt, V.; Coyle, J.; Masson, M.; Thanacoody, R.H.; Gray, A.J.; Webb, D.J.; Moggs, J.G.; et al. Mechanistic Biomarkers Provide Early and Sensitive Detection of Acetaminophen-Induced Acute Liver Injury at First Presentation to Hospital. Hepatology 2013, 58, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Akmal, M.; Baig, M.S.; Khan, J.A. Suppression of Cotton Leaf Curl Disease Symptoms in Gossypium hirsutum through over Expression of Host-Encoded miRNAs. J. Biotechnol. 2017, 263, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Li, Z.; Moore, P.S.; Monaghan, A.P.; Chang, Y.; Nichols, M.; John, B. A Sensitive Non-Radioactive Northern Blot Method to Detect Small RNAs. Nucleic Acids Res. 2010, 38, e98. [Google Scholar] [CrossRef] [PubMed]
- Várallyay, E.; Burgyán, J.; Havelda, Z. MicroRNA Detection by Northern Blotting Using Locked Nucleic Acid Probes. Nat. Protoc. 2008, 3, 190–196. [Google Scholar] [CrossRef]
- Li, W.; Ruan, K. MicroRNA Detection by Microarray. Anal. Bioanal. Chem. 2009, 394, 1117–1124. [Google Scholar] [CrossRef]
- Cissell, K.A.; Deo, S.K. Trends in microRNA Detection. Anal. Bioanal. Chem. 2009, 394, 1109–1116. [Google Scholar] [CrossRef]
- Mohammadi-Yeganeh, S.; Paryan, M.; Mirab Samiee, S.; Soleimani, M.; Arefian, E.; Azadmanesh, K.; Mostafavi, E.; Mahdian, R.; Karimipoor, M. Development of a Robust, Low Cost Stem-Loop Real-Time Quantification PCR Technique for miRNA Expression Analysis. Mol. Biol. Rep. 2013, 40, 3665–3674. [Google Scholar] [CrossRef]
- Niu, Y.; Zhang, L.; Qiu, H.; Wu, Y.; Wang, Z.; Zai, Y.; Liu, L.; Qu, J.; Kang, K.; Gou, D. An Improved Method for Detecting Circulating microRNAs with S-Poly(T) Plus Real-Time PCR. Sci. Rep. 2015, 5, 15100. [Google Scholar] [CrossRef]
- Ouyang, T.; Liu, Z.; Han, Z.; Ge, Q. MicroRNA Detection Specificity: Recent Advances and Future Perspective. Anal. Chem. 2019, 91, 3179–3186. [Google Scholar] [CrossRef]
- Persano, S.; Guevara, M.L.; Wolfram, J.; Blanco, E.; Shen, H.; Ferrari, M.; Pompa, P.P. Label-Free Isothermal Amplification Assay for Specific and Highly Sensitive Colorimetric miRNA Detection. ACS Omega 2016, 1, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Batule, B.S.; Kang, K.S.; Park, K.S.; Park, H.G. Rapid and Ultrasensitive Detection of microRNA by Target-Assisted Isothermal Exponential Amplification Coupled with Poly (Thymine)-Templated Fluorescent Copper Nanoparticles. Nanotechnology 2016, 27, 425502. [Google Scholar] [CrossRef]
- Yang, H.; Jin, Y.; Qian, H.; Wang, Y.; Bao, T.; Wu, Z.; Wen, W.; Zhang, X.; Wang, S. Target-Driven Cascade Amplified Assembly of Covalent Organic Frameworks on Tetrahedral DNA Nanostructure with Multiplex Recognition Domains for Ultrasensitive Detection of microRNA. Anal. Chim. Acta 2024, 1311, 342743. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Li, P.; He, W.; Liu, C.; Li, Z. Rolling Circle Extension-Actuated Loop-Mediated Isothermal Amplification (RCA-LAMP) for Ultrasensitive Detection of microRNAs. Biosens. Bioelectron. 2019, 128, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, Y.; Zhang, S.; Sun, M.; Li, W.; Jiang, Y.; Wu, Z.-S. Ultrasensitive Assay Based on a Combined Cascade Amplification by Nicking-Mediated Rolling Circle Amplification and Symmetric Strand-Displacement Amplification. Anal. Chim. Acta 2019, 1047, 172–178. [Google Scholar] [CrossRef]
- Xu, H.; Wu, D.; Zhang, Y.; Shi, H.; Ouyang, C.; Li, F.; Jia, L.; Yu, S.; Wu, Z.-S. RCA-Enhanced Multifunctional Molecule Beacon-Based Strand-Displacement Amplification for Sensitive microRNA Detection. Sens. Actuators B Chem. 2018, 258, 470–477. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, H.; Yu, H.; Jiang, T.; Luo, Y. Duplex-Specific Nuclease-Mediated Bioanalysis. Trends Biotechnol. 2015, 33, 180–188. [Google Scholar] [CrossRef]
- Shi, C.; Liu, Q.; Ma, C.; Zhong, W. Exponential Strand-Displacement Amplification for Detection of MicroRNAs. Anal. Chem. 2014, 86, 336–339. [Google Scholar] [CrossRef]
- Lin, X.; Jiang, J.; Wang, J.; Xia, J.; Wang, R.; Diao, G. Competitive Host-Guest Recognition Initiated by DNAzyme-Cleavage Cycling for Novel Ratiometric Electrochemical Assay of miRNA-21. Sens. Actuators B Chem. 2021, 333, 129556. [Google Scholar] [CrossRef]
- Li, X.; Chen, L. Fluorescence Probe Based on an Amino-Functionalized Fluorescent Magnetic Nanocomposite for Detection of Folic Acid in Serum. ACS Appl. Mater. Interfaces 2016, 8, 31832–31840. [Google Scholar] [CrossRef]
- Yang, M.; Huang, J.; Fan, J.; Du, J.; Pu, K.; Peng, X. Chemiluminescence for Bioimaging and Therapeutics: Recent Advances and Challenges. Chem. Soc. Rev. 2020, 49, 6800–6815. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Woo, J.; Shin, J.; Cha, B.S.; Kim, S.; Park, K.S. Tetrahedral DNA Nanostructures Enhance Transcription Isothermal Amplification for Multiplex Detection of Non-Coding RNAs. Biosens. Bioelectron. 2024, 250, 116055. [Google Scholar] [CrossRef] [PubMed]
- Ruckman, J.; Green, L.S.; Beeson, J.; Waugh, S.; Gillette, W.L.; Henninger, D.D.; Claesson-Welsh, L.; Janjić, N. 2′-Fluoropyrimidine RNA-Based Aptamers to the 165-Amino Acid Form of Vascular Endothelial Growth Factor (VEGF165). Inhibition of Receptor Binding and VEGF-Induced Vascular Permeability through Interactions Requiring the Exon 7-Encoded Domain. J. Biol. Chem. 1998, 273, 20556–20567. [Google Scholar] [CrossRef] [PubMed]
- Sazani, P.L.; Larralde, R.; Szostak, J.W. A Small Aptamer with Strong and Specific Recognition of the Triphosphate of ATP. J. Am. Chem. Soc. 2004, 126, 8370–8371. [Google Scholar] [CrossRef]
- Ueki, R.; Atsuta, S.; Ueki, A.; Hoshiyama, J.; Li, J.; Hayashi, Y.; Sando, S. DNA Aptamer Assemblies as Fibroblast Growth Factor Mimics and Their Application in Stem Cell Culture. Chem. Commun. 2019, 55, 2672–2675. [Google Scholar] [CrossRef]
- Ueki, R.; Uchida, S.; Kanda, N.; Yamada, N.; Ueki, A.; Akiyama, M.; Toh, K.; Cabral, H.; Sando, S. A Chemically Unmodified Agonistic DNA with Growth Factor Functionality for in Vivo Therapeutic Application. Sci. Adv. 2020, 6, eaay2801. [Google Scholar] [CrossRef]
- He, F.; Wang, M.; Wang, J.; Wang, H.-H.; Nie, Z. An Extracellular miRNA-Responsive Artificial Receptor via Dynamic DNA Nano-Assembly for Biomarker-Driven Therapy. Angew. Chem. Int. Ed. Engl. 2023, 62, e202305227. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Seelig, G. Dynamic DNA Nanotechnology Using Strand-Displacement Reactions. Nat. Chem. 2011, 3, 103–113. [Google Scholar] [CrossRef]
- Madsen, M.; Gothelf, K.V. Chemistries for DNA Nanotechnology. Chem. Rev. 2019, 119, 6384–6458. [Google Scholar] [CrossRef]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, M.B. miR-122—A Key Factor and Therapeutic Target in Liver Disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef]
- Huh, C.-G.; Factor, V.M.; Sánchez, A.; Uchida, K.; Conner, E.A.; Thorgeirsson, S.S. Hepatocyte Growth Factor/c-Met Signaling Pathway Is Required for Efficient Liver Regeneration and Repair. Proc. Natl. Acad. Sci. USA 2004, 101, 4477–4482. [Google Scholar] [CrossRef] [PubMed]
- Bissels, U.; Wild, S.; Tomiuk, S.; Holste, A.; Hafner, M.; Tuschl, T.; Bosio, A. Absolute Quantification of microRNAs by Using a Universal Reference. RNA 2009, 15, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, J.M.; Seila, A.C.; Yeo, G.W.; Sharp, P.A. RNA Sequence Analysis Defines Dicer’s Role in Mouse Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 18097–18102. [Google Scholar] [CrossRef] [PubMed]
- Atic, A.I.; Thiele, M.; Munk, A.; Dalgaard, L.T. Circulating miRNAs Associated with Nonalcoholic Fatty Liver Disease. Am. J. Physiol.-Cell Physiol. 2023, 324, C588–C602. [Google Scholar] [CrossRef]
- Exploring Mechanisms of FGF Signalling through the Lens of Structural Biology. Available online: https://pubmed.ncbi.nlm.nih.gov/23403721/ (accessed on 3 July 2024).
- Andrade, R.J.; Chalasani, N.; Björnsson, E.S.; Suzuki, A.; Kullak-Ublick, G.A.; Watkins, P.B.; Devarbhavi, H.; Merz, M.; Lucena, M.I.; Kaplowitz, N.; et al. Drug-Induced Liver Injury. Nat. Rev. Dis. Primers 2019, 5, 58. [Google Scholar] [CrossRef]
- Ding, D.; Zhao, H.; Wei, D.; Yang, Q.; Yang, C.; Wang, R.; Chen, Y.; Li, L.; An, S.; Xia, Q.; et al. The First-in-Human Whole-Body Dynamic Pharmacokinetics Study of Aptamer. Research 2023, 6, 0126. [Google Scholar] [CrossRef]
- Chen, X.; Xu, K.; Li, J.; Yang, M.; Li, X.; Chen, Q.; Lu, C.; Yang, H. Switch-Conversional Ratiometric Fluorescence Biosensor for miRNA Detection. Biosens. Bioelectron. 2020, 155, 112104. [Google Scholar] [CrossRef]
- Yang, R.; Jiang, G.; Liu, H.; He, L.; Yu, F.; Liu, L.; Qu, L.; Wu, Y. A Dual-Model “on-Super off” Photoelectrochemical/Ratiometric Electrochemical Biosensor for Ultrasensitive and Accurate Detection of microRNA-224. Biosens. Bioelectron. 2021, 188, 113337. [Google Scholar] [CrossRef]
- Pirola, C.J.; Fernández Gianotti, T.; Castaño, G.O.; Mallardi, P.; San Martino, J.; Mora Gonzalez Lopez Ledesma, M.; Flichman, D.; Mirshahi, F.; Sanyal, A.J.; Sookoian, S. Circulating microRNA Signature in Non-Alcoholic Fatty Liver Disease: From Serum Non-Coding RNAs to Liver Histology and Disease Pathogenesis. Gut 2015, 64, 800–812. [Google Scholar] [CrossRef]
- Waidmann, O.; Bihrer, V.; Pleli, T.; Farnik, H.; Berger, A.; Zeuzem, S.; Kronenberger, B.; Piiper, A. Serum microRNA-122 Levels in Different Groups of Patients with Chronic Hepatitis B Virus Infection. J. Viral. Hepat. 2012, 19, e58–e65. [Google Scholar] [CrossRef]
- van der Meer, A.J.; Farid, W.R.R.; Sonneveld, M.J.; de Ruiter, P.E.; Boonstra, A.; van Vuuren, A.J.; Verheij, J.; Hansen, B.E.; de Knegt, R.J.; van der Laan, L.J.W.; et al. Sensitive Detection of Hepatocellular Injury in Chronic Hepatitis C Patients with Circulating Hepatocyte-Derived microRNA-122. J. Viral. Hepat. 2013, 20, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Nakao, K.; Miyaaki, H.; Ichikawa, T. Antitumor Function of microRNA-122 against Hepatocellular Carcinoma. J. Gastroenterol. 2014, 49, 589–593. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, T.; Fan, X.; Wang, M.; Duan, Z.; He, F.; Wang, H.-H.; Li, Z. Development of a Modular miRNA-Responsive Biosensor for Organ-Specific Evaluation of Liver Injury. Biosensors 2024, 14, 450. https://doi.org/10.3390/bios14090450
Zhang X, Wang T, Fan X, Wang M, Duan Z, He F, Wang H-H, Li Z. Development of a Modular miRNA-Responsive Biosensor for Organ-Specific Evaluation of Liver Injury. Biosensors. 2024; 14(9):450. https://doi.org/10.3390/bios14090450
Chicago/Turabian StyleZhang, Xinxin, Tingting Wang, Xiangqing Fan, Meixia Wang, Zhixi Duan, Fang He, Hong-Hui Wang, and Zhihong Li. 2024. "Development of a Modular miRNA-Responsive Biosensor for Organ-Specific Evaluation of Liver Injury" Biosensors 14, no. 9: 450. https://doi.org/10.3390/bios14090450
APA StyleZhang, X., Wang, T., Fan, X., Wang, M., Duan, Z., He, F., Wang, H.-H., & Li, Z. (2024). Development of a Modular miRNA-Responsive Biosensor for Organ-Specific Evaluation of Liver Injury. Biosensors, 14(9), 450. https://doi.org/10.3390/bios14090450