CRISPR/Cas12a Collateral Cleavage Activity for Sensitive 3′–5′ Exonuclease Assay



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Exo III Activity Detection Assay
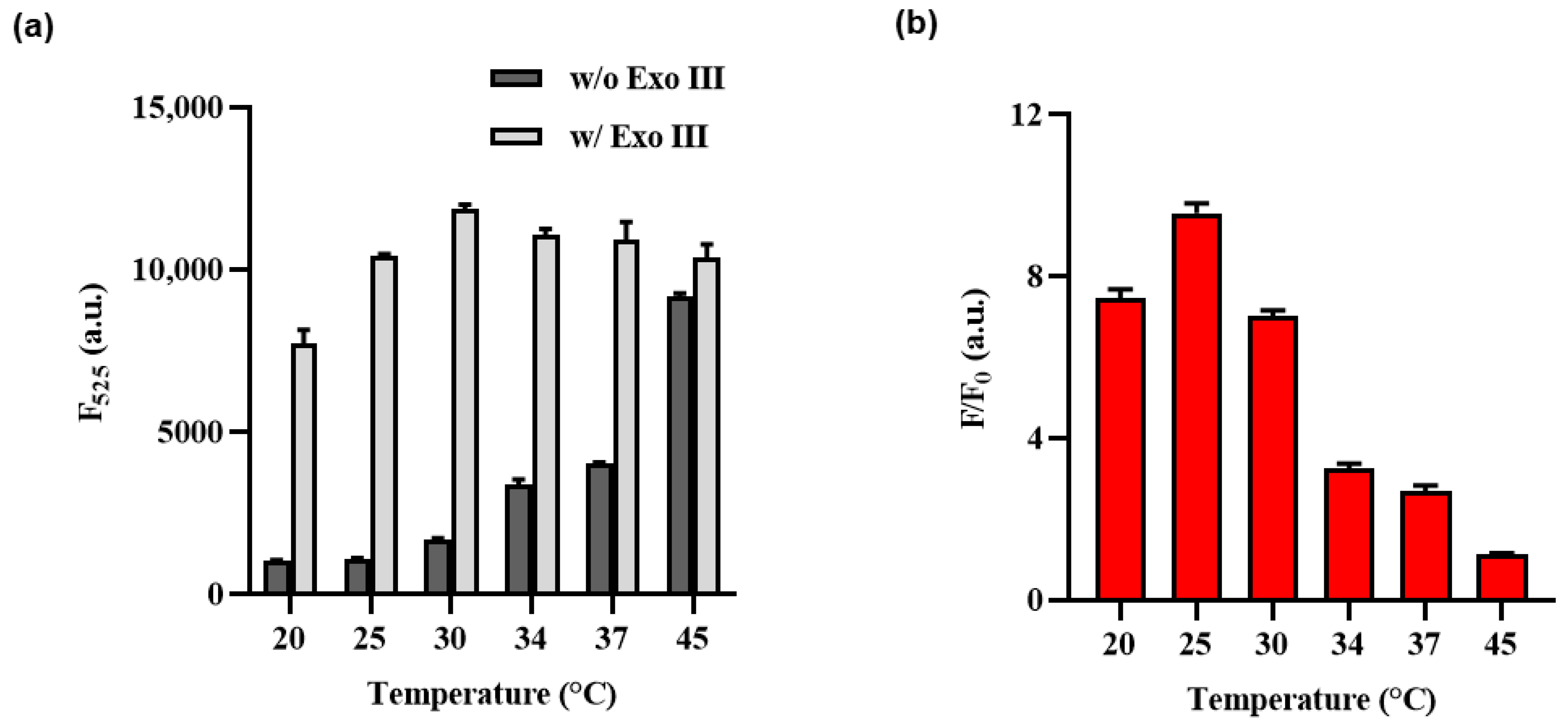
2.3. Optimizing the Cas12a Reaction Temperature Condition of Exo III Activity Detection Assay
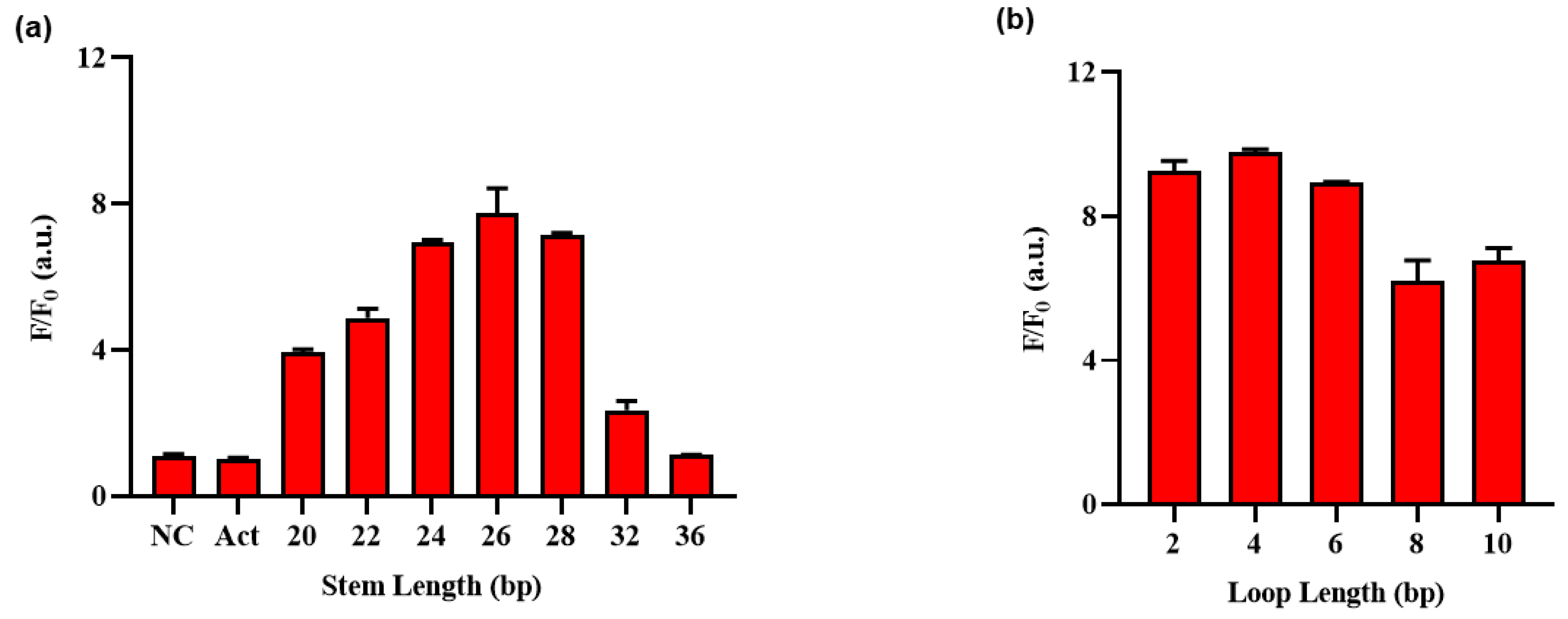
2.4. Optimizing the HP Loop and Stem Length Condition of Exo III Activity Detection Assay
2.5. Polyacrylamide Gel Electrophoresis (PAGE) Analysis
2.6. Exo III Inhibition Assay
3. Results
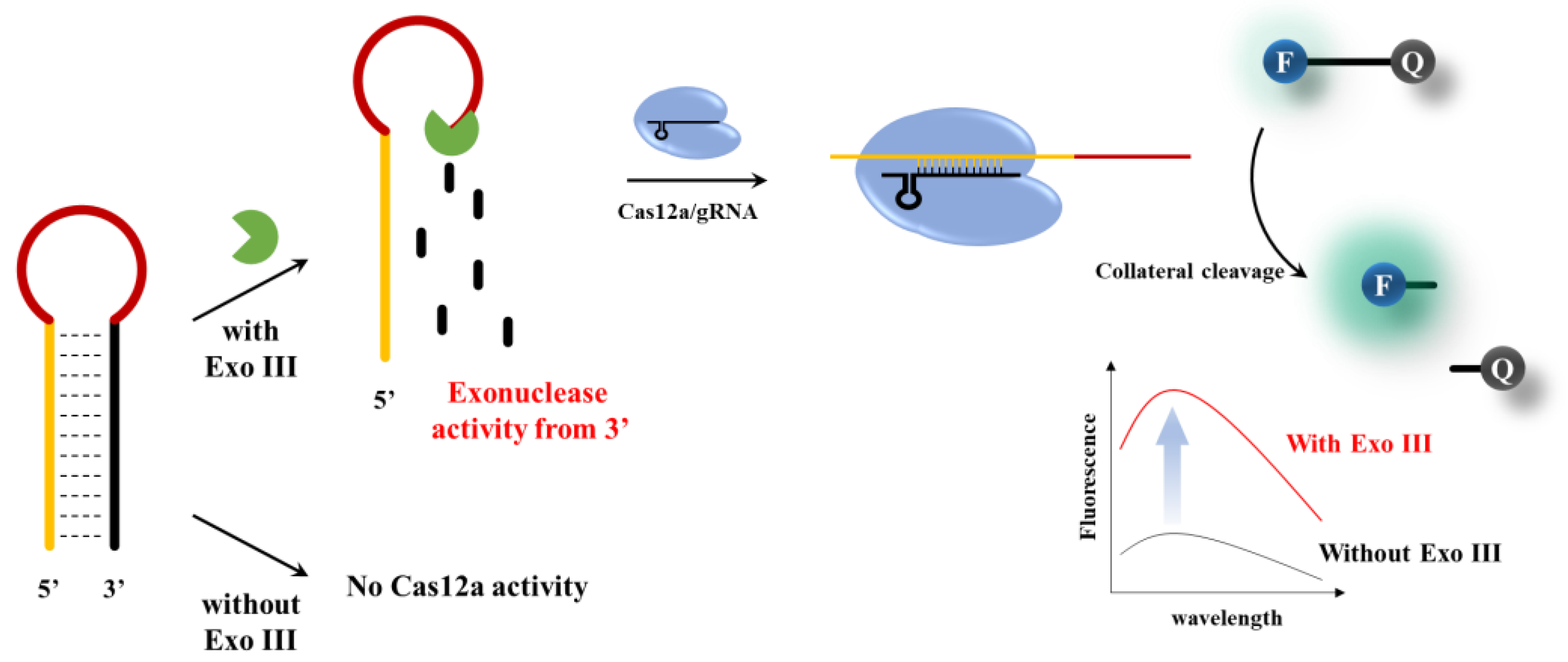
3.1. Detection Principle of Exo III Activity Assay
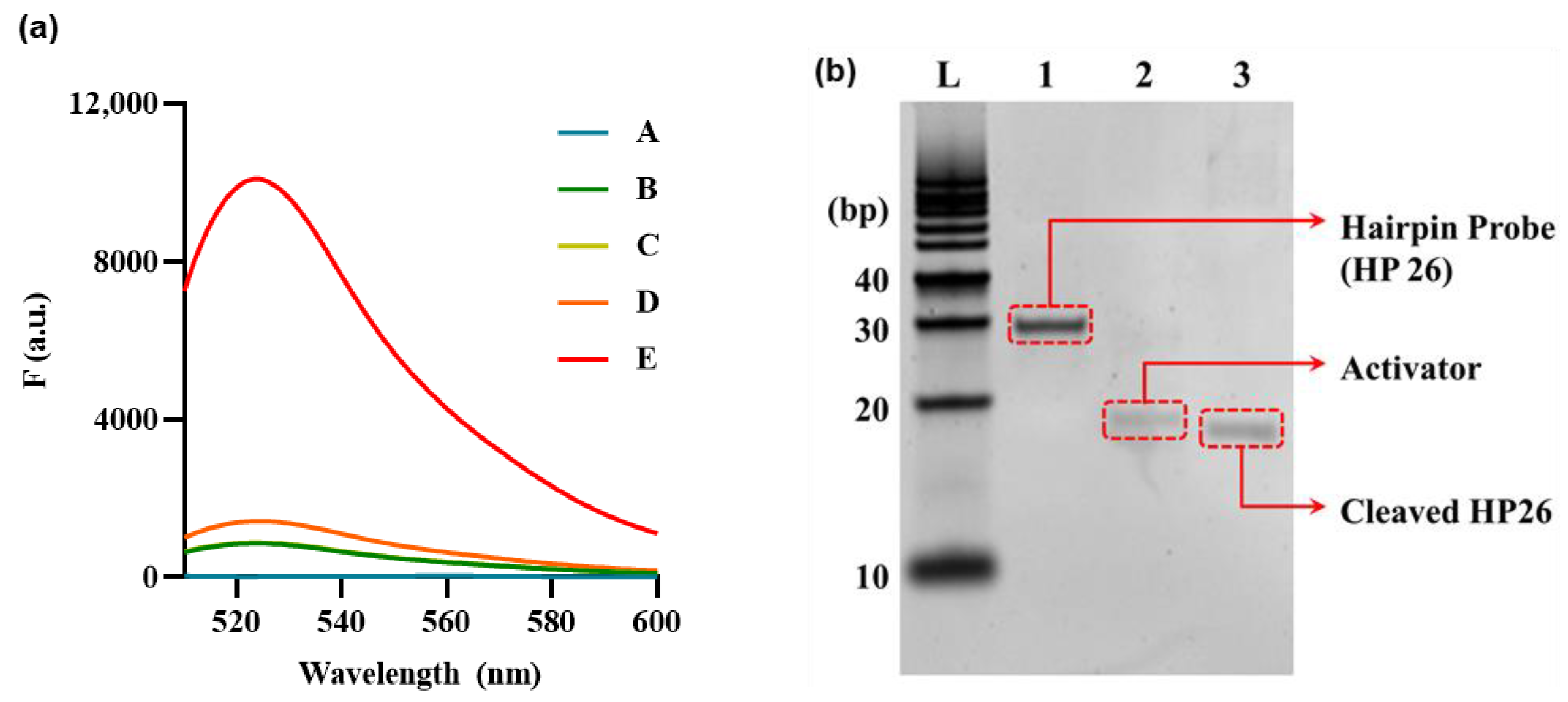
3.2. Feasibility Investigation
3.3. Optimization of Experimental Condition
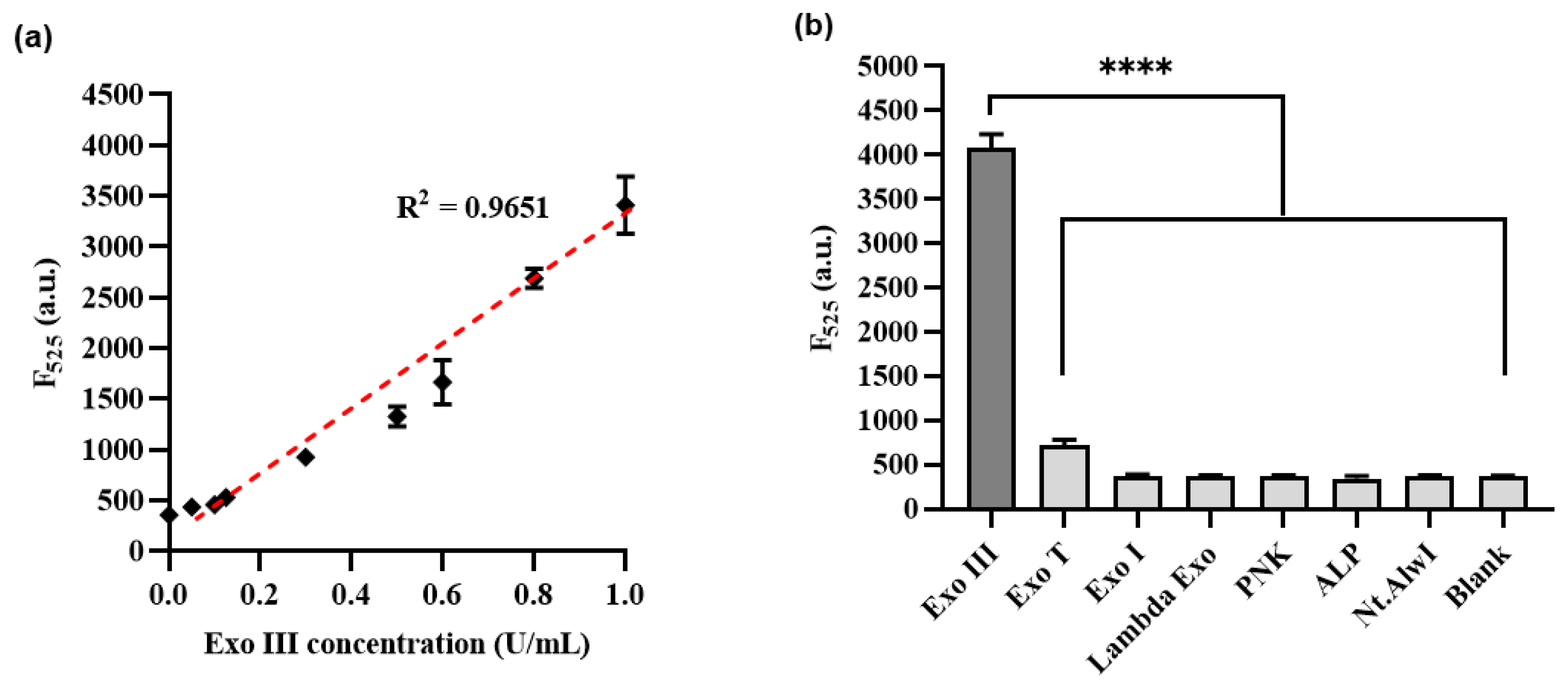
3.4. Detection Performance of Exo III Activity Assay
3.5. Inhibitory Exo III
3.6. Real–Sample Test of the CRISPR/Cas12a Based Exo III Activity Detection Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paull, T.T.; Gellert, M. The 3 to 5 Exonuclease Activity of Mre11 Facilitates Repair of DNA Double-Strand Breaks. Mol. Cell 1998, 1, 969–979. [Google Scholar] [CrossRef]
- Mol, C.D.; Kuo, C.-F.; Thayer, M.M.; Cunningham, R.P.; Tainer, J.A. Structure and Function of the Multifunctional DNA-Repair Enzyme Exonuclease III. Nature 1995, 374, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bai, Y.; Jiang, Y.; Wang, N.; Yang, F.; Zhan, L.; Huang, C. Homo-FRET Enhanced Ratiometric Fluorescence Strategy for Exonuclease III Activity Detection. Anal. Methods 2021, 13, 1489–1494. [Google Scholar] [CrossRef]
- Chekanova, J.A.; Shaw, R.J.; Wills, M.A.; Belostotsky, D.A. Poly(A) Tail-Dependent Exonuclease AtRrp41p from Arabidopsis Thaliana Rescues 5.8 S RRNA Processing and MRNA Decay Defects of the Yeast Ski6 Mutant and Is Found in an Exosome-Sized Complex in Plant and Yeast Cells. J. Biol. Chem. 2000, 275, 33158–33166. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhu, X.; Zhang, C.; Xiao, X.; Zhao, M. In Situ, Real-Time Monitoring of the 3′ to 5′ Exonucleases Secreted by Living Cells. Anal. Chem. 2012, 84, 5059–5065. [Google Scholar] [CrossRef]
- Jain, R.; Aggarwal, A.K.; Rechkoblit, O. Eukaryotic DNA Polymerases. Curr. Opin. Struct. Biol. 2018, 53, 77–87. [Google Scholar] [CrossRef]
- Shevelev, I.V.; Hübscher, U. The 3′–5′ Exonucleases. Nat. Rev. Mol. Cell Biol. 2002, 3, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Brucet, M.; Querol-Audí, J.; Bertlik, K.; Lloberas, J.; Fita, I.; Celada, A. Structural and Biochemical Studies of TREX1 Inhibition by Metals. Identification of a New Active Histidine Conserved in DEDDh Exonucleases. Protein Sci. 2008, 17, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Lehtinen, D.A.; Harvey, S.; Mulcahy, M.J.; Hollis, T.; Perrino, F.W. The TREX1 Double-Stranded DNA Degradation Activity Is Defective in Dominant Mutations Associated with Autoimmune Disease. J. Biol. Chem. 2008, 283, 31649–31656. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, H.; Khusbu, F.Y.; Ma, C.; Ping, A.; Zhang, Q.; Wu, K.; Chen, M. Label-Free Detection of Exonuclease III Activity and Its Inhibition Based on DNA Hairpin Probe. Anal. Biochem. 2018, 555, 55–58. [Google Scholar] [CrossRef]
- Lee, C.; Gang, J. Label-Free Rapid and Simple Detection of Exonuclease III Activity with DNA-Templated Copper Nanoclusters. J. Microbiol. Biotechnol. 2018, 28, 1467–1472. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, Z.; Su, X. Label-Free Detection of Exonuclease III by Using DsDNA-Templated Copper Nanoparticles as Fluorescent Probe. Talanta 2015, 131, 59–63. [Google Scholar] [CrossRef]
- Umer, A.; Naveed, S.; Ramzan, N.; Rafique, M.S. Selection of a Suitable Method for the Synthesis of Copper Nanoparticles. Nano 2012, 7, 1230005. [Google Scholar] [CrossRef]
- Park, J.; Han, H.; Park, C.; Ahn, J.K. Washing-Free and Label-Free Onsite Assay for Inorganic Pyrophosphatase Activity Using a Personal Glucose Meter. Anal. Chem. 2022, 94, 11508–11513. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, F.; Zhao, D.; Yang, X. Highly Sensitive Real-Time Assay of Inorganic Pyrophosphatase Activity Based on the Fluorescent Gold Nanoclusters. Anal. Chem. 2014, 86, 7883–7889. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Feng, X.; Gao, T.; Wang, R.; Mao, Y.; Lin, J.; Yu, X.; Luo, X. A Novel Dual-Functional Biosensor for Fluorometric Detection of Inorganic Pyrophosphate and Pyrophosphatase Activity Based on Globulin Stabilized Gold Nanoclusters. Anal. Chim. Acta 2017, 958, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, J.; Peng, H.; Jiang, J.; Yu, R. A Novel Fluorescence Assay for Inorganic Pyrophosphatase Based on Modulated Aggregation of Graphene Quantum Dots. Analyst 2016, 141, 251–255. [Google Scholar] [CrossRef]
- Xu, M.; Li, B. Label-Free Fluorescence Strategy for Sensitive Detection of Exonuclease Activity Using SYBR Green i as Probe. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 151, 22–26. [Google Scholar] [CrossRef]
- Paul, B.; Montoya, G. CRISPR-Cas12a: Functional Overview and Applications. Biomed. J. 2020, 43, 8–17. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, H.; Wang, H.; Wang, F.; Li, Z. CRISPR/Cas12a-Assisted Ligation-Initiated Loop-Mediated Isothermal Amplification (CAL-LAMP) for Highly Specific Detection of MicroRNAs. Anal. Chem. 2021, 93, 7942–7948. [Google Scholar] [CrossRef]
- Kim, S.; Broströmer, E.; Xing, D.; Jin, J.; Chong, S.; Ge, H.; Wang, S.; Gu, C.; Yang, L.; Gao, Y.Q.; et al. Probing Allostery through DNA. Science 2013, 339, 816–819. [Google Scholar] [CrossRef]
- Wang, Z.; Cui, W. CRISPR-Cas System for Biomedical Diagnostic Platforms. VIEW 2020, 1, 20200008. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a Target Binding Unleashes Indiscriminate Single-Stranded DNase Activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Yang, H.; Rajashankar, K.R.; Huang, Z.; Patel, D.J. Type v CRISPR-Cas Cpf1 Endonuclease Employs a Unique Mechanism for CrRNA-Mediated Target DNA Recognition. Cell Res. 2016, 26, 901–913. [Google Scholar] [CrossRef]
- Swarts, D.C.; Jinek, M. Mechanistic Insights into the Cis- and Trans-Acting DNase Activities of Cas12a. Mol. Cell 2019, 73, 589–600.e4. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, S.; Lee, J.; Park, H.G. CRISPR/Cas12a Collateral Cleavage Activity for an Ultrasensitive Assay of RNase H. Chem. Commun. 2022, 58, 2654–2657. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic Acid Detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef]
- Li, S.Y.; Cheng, Q.X.; Li, X.Y.; Zhang, Z.L.; Gao, S.; Cao, R.B.; Zhao, G.P.; Wang, J.; Wang, J.M. CRISPR-Cas12a-Assisted Nucleic Acid Detection. Cell Discov. 2018, 4, 20. [Google Scholar] [CrossRef]
- Wang, D.X.; Wang, J.; Du, Y.C.; Ma, J.Y.; Wang, S.Y.; Tang, A.N.; Kong, D.M. CRISPR/Cas12a-Based Dual Amplified Biosensing System for Sensitive and Rapid Detection of Polynucleotide Kinase/Phosphatase. Biosens. Bioelectron. 2020, 168, 112556. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Y.; Yu, Y.; Zhang, Y.; Peng, J.; Niu, L.; Zhang, J. Hydrazone Ligation Assisted DNAzyme Walking Nanomachine Coupled with CRISPR-Cas12a for Lipopolysaccharide Analysis. Anal. Chim. Acta 2021, 1174, 338747. [Google Scholar] [CrossRef]
- Hang, X.M.; Zhao, K.R.; Wang, H.Y.; Liu, P.F.; Wang, L. Exonuclease III-Assisted CRISPR/Cas12a Electrochemiluminescence Biosensor for Sub-Femtomolar Mercury Ions Determination. Sens. Actuators B Chem. 2022, 368, 132208. [Google Scholar] [CrossRef]
- Fu, R.; Wang, Y.; Liu, Y.; Liu, H.; Zhao, Q.; Zhang, Y.; Wang, C.; Li, Z.; Jiao, B.; He, Y. CRISPR-Cas12a Based Fluorescence Assay for Organophosphorus Pesticides in Agricultural Products. Food Chem. 2022, 387, 132919. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Gupta, V. Methods for the Determination of Limit of Detection and Limit of Quantitation of the Analytical Methods. Chron. Young Sci. 2011, 2, 21. [Google Scholar] [CrossRef]
- Yang, W.; Ruan, Y.; Wu, W.; Chen, P.; Xu, L.; Fu, F. A “Turn-on” and Label-Free Fluorescent Assay for the Rapid Detection of Exonuclease III Activity Based on Tb3+-Induced G-Quadruplex Conjugates. Anal. Bioanal. Chem. 2014, 406, 4535–4540. [Google Scholar] [CrossRef] [PubMed]
- He, H.Z.; Chan, W.I.; Mak, T.Y.; Liu, L.J.; Wang, M.; Chan, D.S.H.; Ma, D.L.; Leung, C.H. Detection of 3′→5′ Exonuclease Activity Using a Metal-Based Luminescent Switch-on Probe. Methods 2013, 64, 218–223. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Wu, X.; Zhao, J.X. A Graphene Oxide-Based Fluorescence Assay for the Sensitive Detection of DNA Exonuclease Enzymatic Activity. Analyst 2019, 144, 6231–6239. [Google Scholar] [CrossRef]
- Liu, Q.; Lian, J.; Liu, M.; Jin, Y.; Li, B. A Simple “Turn-on” Fluorescent Biosensor for Sensitive Detection of Exonuclease III Activity through Photoinduced Electron Transfer and Self-Hybridization of a DNA Probe. Anal. Methods 2018, 10, 2257–2262. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, H.; Jing, J.; Zhang, X. A Label-Free Ratiometric Fluorescence Strategy for 3′–5′ Exonuclease Detection. N. J. Chem. 2018, 42, 16630–16634. [Google Scholar] [CrossRef]
- Burkin, K.M.; Bodulev, O.L.; Gribas, A.V.; Sakharov, I.Y. One-Step Label-Free Chemiluminescent Assay for Determination of Exonuclease III Activity towards Hairpin Oligonucleotides. Enzym. Microb. Technol. 2019, 131, 109419. [Google Scholar] [CrossRef]
- Silva-Portela, R.C.B.; Carvalho, F.M.; Pereira, C.P.M.; De Souza-Pinto, N.C.; Modesti, M.; Fuchs, R.P.; Agnez-Lima, L.F. ExoMeg1: A New Exonuclease from Metagenomic Library. Sci. Rep. 2016, 6, 19712. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Limit Detection (U/mL) | Linear Range (U/mL) | Ref. |
|---|---|---|---|
| Tb3+ | 0.8 | 5–100 | [35] |
| CuNPs | 0.02 | 0.05–2 | [12] |
| ThT | 0.5 | 0–10 | [10] |
| SYBR Green I | 0.7 | 1–200 | [18] |
| Homo–FRET | 0.17 | 0.25–8 | [3] |
| luminescent | 1 | 0–25 | [36] |
| Graphene oxide | 0.001 | 0.01–0.5 | [37] |
| Photoinduced electron transfer | 0.0003 | 0.0005–5 | [38] |
| NMM/DAPI | 4.42 | 10–100 | [39] |
| Luminol | 4.8 | – | [40] |
| CRISPR/Cas12a | 0.0073 | 0.05–1 | This work |
| Added Exo III (U/mL) | Measured Exo III (μg/mL) a | SD b | CV (%) c | Recovery (%) d |
|---|---|---|---|---|
| 0.85 | 0.88 | 188.25 | 4.377 | 103.5 |
| 0.45 | 0.46 | 123.55 | 5.459 | 102.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeung, J.H.; Han, H.; Lee, C.Y.; Ahn, J.K. CRISPR/Cas12a Collateral Cleavage Activity for Sensitive 3′–5′ Exonuclease Assay. Biosensors 2023, 13, 963. https://doi.org/10.3390/bios13110963
Jeung JH, Han H, Lee CY, Ahn JK. CRISPR/Cas12a Collateral Cleavage Activity for Sensitive 3′–5′ Exonuclease Assay. Biosensors. 2023; 13(11):963. https://doi.org/10.3390/bios13110963
Chicago/Turabian StyleJeung, Jae Hoon, Hyogu Han, Chang Yeol Lee, and Jun Ki Ahn. 2023. "CRISPR/Cas12a Collateral Cleavage Activity for Sensitive 3′–5′ Exonuclease Assay" Biosensors 13, no. 11: 963. https://doi.org/10.3390/bios13110963
APA StyleJeung, J. H., Han, H., Lee, C. Y., & Ahn, J. K. (2023). CRISPR/Cas12a Collateral Cleavage Activity for Sensitive 3′–5′ Exonuclease Assay. Biosensors, 13(11), 963. https://doi.org/10.3390/bios13110963