Simultaneous Observation of Mouse Cortical and Hippocampal Neural Dynamics under Anesthesia through a Cranial Microprism Window

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
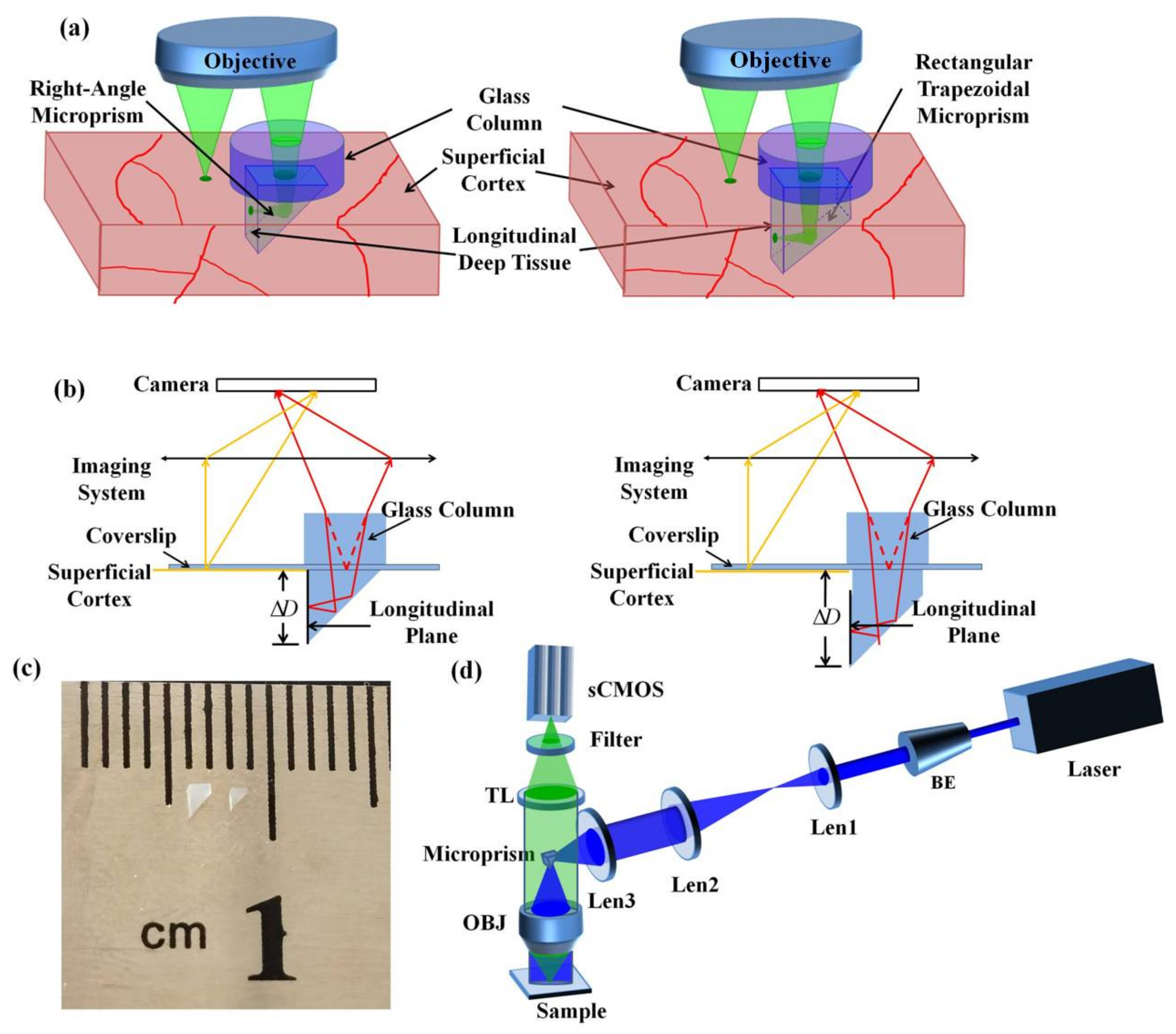
2.1. Strategy of Multi-Planar Imaging
2.2. Optical Mesoscope Setup
2.3. Animal Surgery
2.4. Fluorescent Beads Preparation and Anesthesia Treatment
3. Results
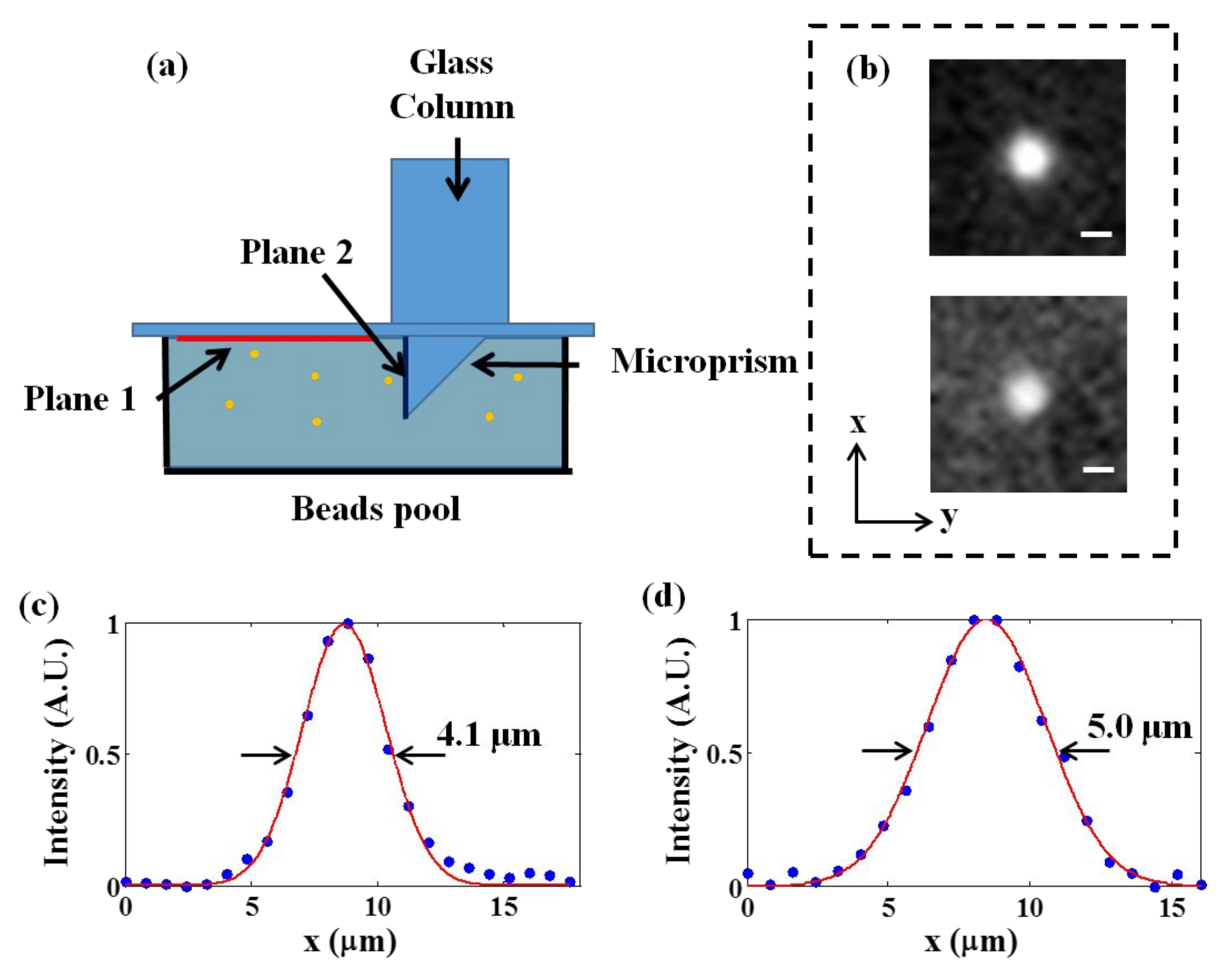
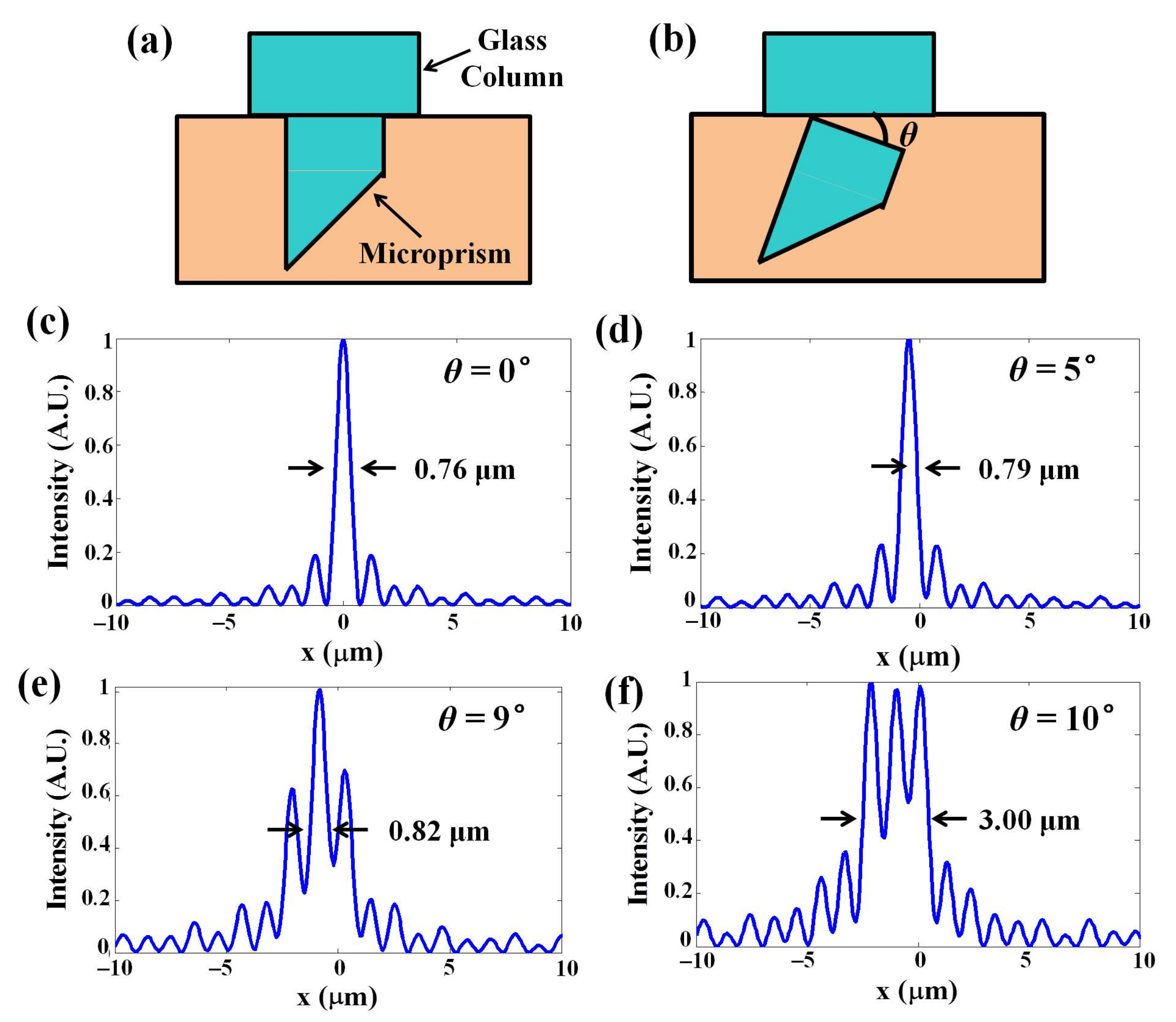
3.1. System Characterization
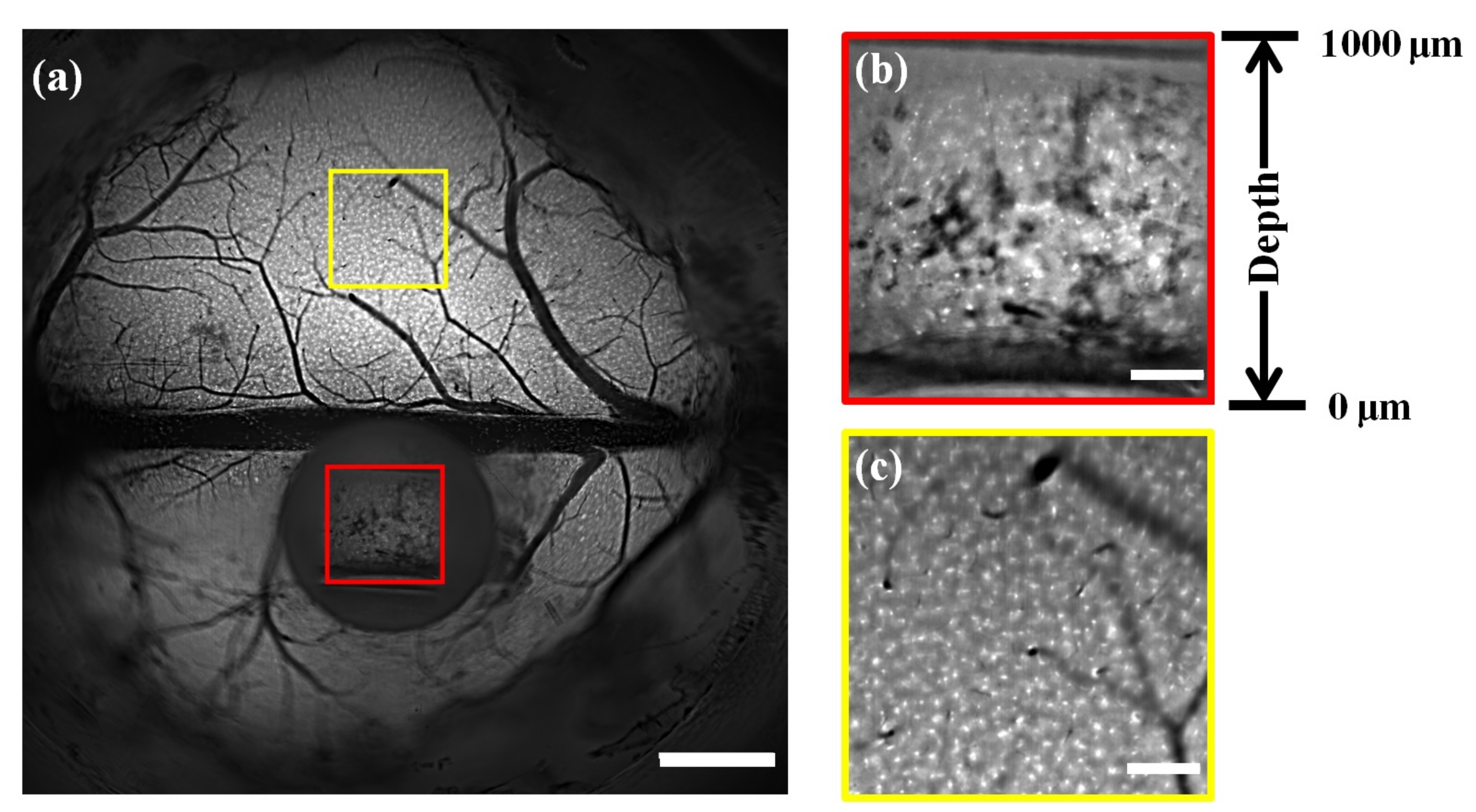
3.2. In Vivo Imaging of Microglial Cells in Mouse Brains by Right-Angle Microprisms
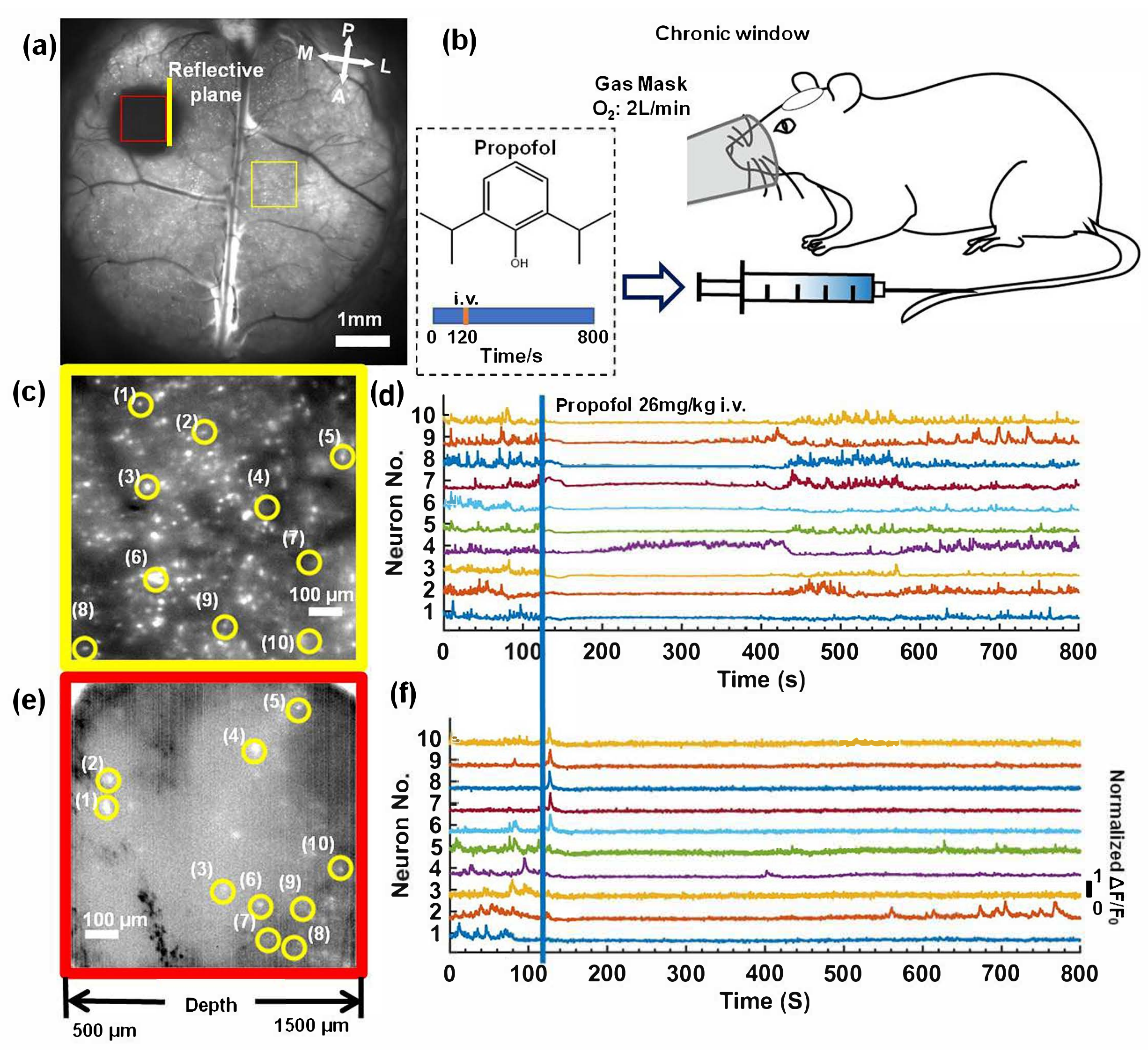
3.3. Deep In Vivo Functional Imaging of Neuron Cells in Mouse Brains by Rectangular Trapezoidal Microprism during Anesthesia
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lichtman, J.W.; Conchello, J.A. Fluorescence microscopy. Nat. Methods 2005, 2, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Suo, J.; Wu, J.; Xie, H.; Shen, Y.; Chen, F.; Wang, G.; Cao, L.; Jin, G.; He, Q.; et al. Video-rate imaging of biological dynamics at centimetre scale and micrometre resolution. Nat. Photonics 2019, 13, 809–816. [Google Scholar] [CrossRef]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, A.; Adams, G.K.; Aura, C.; Leopold, D.A. Distinct superficial and deep laminar domains of activity in the visual cortex during rest and stimulation. Front. Syst. Neurosci. 2010, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Crozier, R.A.; Wang, Y.; Liu, C.H.; Bear, M.F. Deprivation-induced synaptic depression by distinct mechanisms in different layers of mouse visual cortex. Proc. Natl. Acad. Sci. USA 2007, 104, 1383–1388. [Google Scholar] [CrossRef] [Green Version]
- Tolner, E.A.; Kloosterman, F.; van Vliet, E.A.; Witter, M.P.; da Silva, F.H.L.; Gorter, J.A. Presubiculum stimulation in vivo evokes distinct oscillations in superficial and deep entorhinal cortex layers in chronic epileptic rats. J. Neurosci. 2005, 25, 8755–8765. [Google Scholar] [CrossRef] [Green Version]
- Lamme, V.A.; Zipser, K.; Spekreijse, H. Figure-ground activity in primary visual cortex is suppressed by anesthesia. Proc. Natl. Acad. Sci. USA 1998, 95, 3263–3268. [Google Scholar] [CrossRef] [Green Version]
- Aksenov, D.P.; Li, L.; Miller, M.J.; Iordanescu, G.; Wyrwicz, A.M. Effects of anesthesia on BOLD signal and neuronal activity in the somatosensory cortex. J. Cereb. Blood Flow Metab. 2015, 35, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Gaese, B.H.; Ostwald, J. Anesthesia changes frequency tuning of neurons in the rat primary auditory cortex. J. Neurophysiol. 2001, 86, 1062–1066. [Google Scholar] [CrossRef]
- Makino, Y.; Polygalov, D.; Bolaños, F.; Benucci, A.; McHugh, T.J. Physiological signature of memory age in the prefrontal-hippocampal circuit. Cell Rep. 2019, 29, 3835–3846. [Google Scholar] [CrossRef] [Green Version]
- Ying, J.; Keinath, A.T.; Lavoie, R.; Vigneault, E.; El Mestikawy, S.; Brandon, M.P. Disruption of the grid cell network in a mouse model of early Alzheimer’s disease. Nat. Commun. 2022, 13, 886. [Google Scholar] [CrossRef] [PubMed]
- Shin, L.M.; Rauch, S.L.; Pitman, R.K. Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Ann. N. Y. Acad. Sci. 2006, 1071, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Werley, C.A.; Chien, M.P.; Cohen, A.E. Ultrawidefield microscope for high-speed fluorescence imaging and targeted optogenetic stimulation. Biomed. Opt. Express 2017, 8, 5794–5813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofroniew, N.J.; Flickinger, D.; King, J.; Svoboda, K. A large field of view two-photon mesoscope with subcellular resolution for in vivo imaging. elife 2016, 5, e14472. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.H.; Stirman, J.N.; Yu, Y.; Hira, R.; Smith, S.L. Diesel2p mesoscope with dual independent scan engines for flexible capture of dynamics in distributed neural circuitry. Nat. Commun. 2021, 12, 6639. [Google Scholar]
- Wang, K.; Milkie, D.E.; Saxena, A.; Engerer, P.; Misgeld, T.; Bronner, M.E.; Mumm, J.; Betzig, E. Rapid adaptive optical recovery of optimal resolution over large volumes. Nat. Methods 2014, 11, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Sun, W.; Richie, C.T.; Harvey, B.K.; Betzig, E.; Ji, N. Direct wavefront sensing for high-resolution in vivo imaging in scattering tissue. Nat. Commun. 2015, 6, 7276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, N. Adaptive optical fluorescence microscopy. Nat. Methods 2017, 14, 374–380. [Google Scholar] [CrossRef]
- Horton, N.G.; Wang, K.; Kobat, D.; Clark, C.G.; Wise, F.W.; Schaffer, C.B.; Xu, C. In vivo three-photon microscopy of subcortical structures within an intact mouse brain. Nat. Photonics 2013, 7, 205–209. [Google Scholar] [CrossRef]
- Wang, T.; Wu, C.; Ouzounov, D.G.; Gu, W.; Xia, F.; Kim, M.; Yang, X.; Warden, M.R.; Xu, C. Quantitative analysis of 1300-nm three-photon calcium imaging in the mouse brain. eLife 2020, 9, e53205. [Google Scholar] [CrossRef] [PubMed]
- Rathenberg, J.; Nevian, T.; Witzemann, V. High-efficiency transfection of individual neurons using modified electrophysiology techniques. J. Neurosci. Methods 2003, 126, 91–98. [Google Scholar] [CrossRef]
- Moretti, C.; Antonini, A.; Bovetti, S.; Liberale, C.; Fellin, T. Scanless functional imaging of hippocampal networks using patterned two-photon illumination through GRIN lenses. Biomed. Opt. Express 2016, 7, 3958–3967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knittel, J.; Schnieder, L.; Buess, G.; Messerschmidt, B.; Possner, T. Endoscope-compatible confocal microscope using a gradient index-lens system. Opt. Commun. 2001, 188, 267–273. [Google Scholar] [CrossRef]
- Yang, M.; Zhou, Z.; Zhang, J.; Jia, S.; Li, T.; Guan, J.; Liao, X.; Leng, B.; Lyu, J.; Zhang, K.; et al. MATRIEX imaging: Multiarea two-photon real-time in vivo explorer. Light. Sci. Appl. 2019, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckmann, L.; Zhang, X.; Nadkarni, N.A.; Cai, Z.; Batra, A.; Sullivan, D.P.; Muller, W.A.; Sun, C.; Kuranov, R.; Zhang, H.F. Longitudinal deep-brain imaging in mouse using visible-light optical coherence tomography through chronic microprism cranial window. Biomed. Opt. Express 2019, 10, 5235–5250. [Google Scholar] [CrossRef] [PubMed]
- Barson, D.; Hamodi, A.S.; Shen, X.; Lur, G.; Constable, R.T.; Cardin, J.A.; Crair, M.C.; Higley, M.J. Simultaneous mesoscopic and two-photon imaging of neuronal activity in cortical circuits. Nat. Methods 2020, 17, 107–113. [Google Scholar] [CrossRef]
- Andermann, M.L.; Gilfoy, N.B.; Goldey, G.J.; Sachdev, R.N.; Wölfel, M.; McCormick, D.A.; Reid, R.C.; Levene, M.J. Chronic cellular imaging of entire cortical columns in awake mice using microprisms. Neuron 2013, 80, 900–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, C.; Cao, J.; Zhang, R.; Xiao, G.; Hu, J.; Xie, H.; Dai, Q. Real-time brain-wide multi-planar microscopy for simultaneous cortex and hippocampus imaging at the cellular resolution in mice. Biomed. Opt. Express 2021, 12, 1858–1868. [Google Scholar] [CrossRef]
- Speigel, I.A.; Patel, K.; Hemmings, H.C., Jr. Distinct effects of volatile and intravenous anaesthetics on presynaptic calcium dynamics in mouse hippocampal GABAergic neurones. Br. J. Anaesth. 2022, 128, 1019–1028. [Google Scholar] [CrossRef]
- Otto, K.; Thaden, A.V. The Laboratory Mouse, Second ed.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Jeremias, A.; Brown, D.L.; Brown, D.L. Cardiac Intensive Care, 2nd ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2010. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Zhuang, C.; Wang, Z.; Xiao, G.; Chen, K.; Li, H.; Tong, L.; Mi, W.; Xie, H.; Cao, J. Simultaneous Observation of Mouse Cortical and Hippocampal Neural Dynamics under Anesthesia through a Cranial Microprism Window. Biosensors 2022, 12, 567. https://doi.org/10.3390/bios12080567
Zhang R, Zhuang C, Wang Z, Xiao G, Chen K, Li H, Tong L, Mi W, Xie H, Cao J. Simultaneous Observation of Mouse Cortical and Hippocampal Neural Dynamics under Anesthesia through a Cranial Microprism Window. Biosensors. 2022; 12(8):567. https://doi.org/10.3390/bios12080567
Chicago/Turabian StyleZhang, Rujin, Chaowei Zhuang, Zilin Wang, Guihua Xiao, Kunsha Chen, Hao Li, Li Tong, Weidong Mi, Hao Xie, and Jiangbei Cao. 2022. "Simultaneous Observation of Mouse Cortical and Hippocampal Neural Dynamics under Anesthesia through a Cranial Microprism Window" Biosensors 12, no. 8: 567. https://doi.org/10.3390/bios12080567
APA StyleZhang, R., Zhuang, C., Wang, Z., Xiao, G., Chen, K., Li, H., Tong, L., Mi, W., Xie, H., & Cao, J. (2022). Simultaneous Observation of Mouse Cortical and Hippocampal Neural Dynamics under Anesthesia through a Cranial Microprism Window. Biosensors, 12(8), 567. https://doi.org/10.3390/bios12080567